
Abstract
Background and objective: Living organisms respond to physical, chemical, and biological threats with a potent inflammatory response which alters organ cell signaling and leads to dysfunction. We evaluated the therapeutic effect of bone marrow-based mesenchymal stromal cell (BM-MSC) transplanted in rats to preserve tissue integrity and to restore homeostasis and function in the pancreatitis experimental pattern. Methods: This study involved 40 adult male Wister rats. Repeated L-arginine injections caused chronic pancreatitis (CP), leading to the development of pancreatic damage and shifting the intracellular signaling pathways. Rats were then infused with BM-MSC labeled with PKH26 fluorescent linker dye for 12 weeks. Results: Cell-surface indicators of BM-MSCs such as CD 90 and CD29 were expressed with the lack of CD34 expression. BM-MSC treatment considerably improved the alterations induced in a series of inflammatory markers, including IL-18, TNF-α, CRP, PGE2, and MCP-1. Furthermore, improvement was found in digestive enzymes and lipid profile with amelioration in myeloperoxidase activity. BM-MSC treatment also regulated the (TGF-/p-38MPAK/SMAD2/3) signaling factors that enhances repair of damaged pancreatic tissue, confirmed by reversed alteration of histopathological examination. Conclusion: our results further bring to light the promise of cell transplant therapy for chronic pancreatitis.
Introduction
Chronic pancreatitis (CP) is signified with severe inflammatory responses and distinct stroma formation, with a considerable number of macrophages, monocytes, infiltrate granulocytes, including neutrophils, eosinophils, and pancreatic stellate cells (PSCs). 1 Cascade multiple process leads to extracellular matrix (ECM) depositions and the development of fibrosis (tissue scarring). Gland injury leads to ultimate exocrine and endocrine insufficiency. 2 The Acinar cells demand substantial amino acid intake in order to produce pancreatic enzymes. Excessive and accumulated amounts can, therefore, cause pancreatic toxicity by unintelligible mechanisms. L-arginine serves as ligands for the family C or class 3, G-protein coupling (GPCR) receptors, such as CaSR a calcium-sensing, and G-PRC6A.3,4 These two receptors activated in the pancreas induce an imbalance leading to chronic pancreatitis. 5 It is well known that excessive and repeated doses of L-arginine generate free radicals toxically and trigger mitochondrial depolarization in acinar cells, which potentially induces chronic pancreatitis. 6
The pathogenesis of pancreatitis contributes to a complex network of various cytokines such as interleukins and tumor necrosis factor-α (TNF-α), and molecular pathways such as transforming growth factor-β (TGF-β), SMAD and mitogen-activated protein kinases (MAPK). MSCs constitute mature stem cells most essential differentiated into all cell types. Mesenchymal stromal cells (MSCs) appear to be very promising for the treatment of diseases because of their ability to alter the local injured tissue microenvironment, stimulate intracellular progenitor cells, and secrete various factors. 6 It has the ability to self-renew through cell division and to convert into progenitors of tissue-specific perfectly functioning cells that enable pancreatic repairs and regulate immune response and inflammation. This gives us an alternative therapeutic foundation road to be accomplished. 7 MSCs improve and overwhelm fibrosis by numerous signaling pathways, mainly TGF-β1 regulation, which is a key sever inflammation and fibrosis adjuster.8,9
Investigational procedures of chronic pancreatitis useful for the assessment of treatment interventions are not readily available. We chose to investigate the possible efficacy of mesenchymal stromal cells in the rat model of chronic pancreatitis as a disorder of prolonged pancreatic lesions typical of human pancreatitis. Owing to no organ other than the pancreas appears to be impaired by L-arginine and it does not require surgery intervention. 10 A few studies have evaluated potential role of MSCs in chronic pancreatitis’, in which the persistence and consequences of the inflammatory response led to the formation of the extensive fiber region, as a change of pancreatic stellate cell (PSC) to myofibroblast shape. Establishing fibrotic region as a replacement for glandular parenchyma, thus minimizing pancreatic functionality. 11 The curative role of mesenchymal stromal cell infusion in CP has gained a considerable role. However, the fundamental pathways are still unclear. Therefore, this work aimed to study the possible potential targeting of MSC stem cell treatment against chronic pancreatitis induced by L-arginine in rats.
Materials and methods
Materials and kits
L-arginine and other components used in this study were purchased from Sigma–Aldrich® (St Louis, Missouri, USA). ELISA plat reader used to determine protein level of pancreatic rat tissue TGF-β1 and monocyte chemotactic protein-1 (MCP-1) (Biotek, USA) and serum TNF-α (Abcam, MA, USA). The serum concentrations of C-reactive protein (CRP), prostaglandin (PGE2) and tissue MPO (R & D Systems, Minneapolis, MN), and serum IL-18 (Cusabio, Technology, LLC), were determined via ELISA kits as advised by the manufacturer. anti-p-38 MPAK and anti-p-SMAD2/3 were purchased from Thermo Fisher Scientific Inc., USA for western analysis.
BM-MSCs preparation and culture
The BM was extracted from the femurs and tibias of rats 6 weeks old, then suspended in a culture medium in vitro with 10% fetal bovine serum, Dulbecco’s modified Eagle’s, DMEM, and GIBCO/BRL. In a sufficient medium, 1% penicillin streptomycin was resuspended, cells were collected then incubated in primary culture at (37°C, 5%) humidified CO2 for 12 days. Once confluence large colonies (80–90%) formed, the cultivation was double washed with saline phosphate buffer (PBS) and trypsin 0.25% was added to 1mM EDTA. Cells were centrifuged, resuspended in serum-complement medium then incubated in a flask (50 cm2). We referred to the resulting cultures as first-passage cultures. 12 Their adhesiveness and fusiform shape characterize the MSCs which were identified by flow cytometer as an estimate of the positive expressions of (CD 90, CD29) and the negative of CD34 surface markers of BM-MSCs. 13
Investigational model
In this study, Six weeks old male Wistar rats (170 ± 30 g; n = 40) were used. Rats were housed in a ventilated animal household with standard laboratory conditions and subjected to a 12:12-h light-dark cycle and had free access to water and food. All the ethical protocols for animal treatment were followed by the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 85, revised 1996) and supervised by the Research Ethics Committee of the National Centre for Radiation Research and Technology, Atomic Energy Authority (Ref. no. 202/21). Rats were allocated and randomly divided into four equal groups with 10 rats each to enable differences in treatment to be determined with statistical significance (p < 0.05) as determined using the G-power statistical program. 14
Control group: Untreated normal rats
CP group: Rats were given two intraperitoneal (I.P) injections of L-arginine hydrochloride (250 mg/100 g body mass) in normal saline as two repeated doses at 1-hour intervals on days (1, 5, 9, 13, and 17) 15 and the rats were sacrificed after 12 weeks.
BM-MSCs group: Rats received one single injection I.V. into the tail vein with (1 × 107) cells in 0.2 mL labeled with PKH26 fluorescent linker dye in phosphate buffer saline (PBS) and after 12 weeks the rats were sacrificed. 16
CP + BM-MSCs group: Following the dosage of L-arginine hydrochloride, rats were treated with one injection as in the BM-MSCs group. At the end of the experimental period of 12 weeks, rats were anaesthetized by I.P. injection of urethane (1.2 g/kg b.w), 15 then sacrificed by cervical decapitation. Blood was collected in heparinized and non-heparinized tubes to separate plasma and serum; respectively, as centrifuged at (3000 × g) for 15 min using a Hettich Mikro 22R centrifuge UP8U ’I (Tuttlingen, Germany). Dissected pancreas tissue samples were collected and separated into two parts each; the first portion was stored at −80°C for further molecular and biochemical analysis. The second portion of pancreas tissues were fixed in 10% neutral formalin and processed for histological evaluation.
Experimental parameters
Lipase and amylase analysis
Sera of pancreases enzymatic activities were assessed using a spectrophotometric approach, which was carried out in accordance with a previously established procedure. 17
Lipid profile assay
In serum, total cholesterol (TC), triglyceride (TG), and high-density lipoprotein cholesterol (HDL-C) were determined by Bio diagnostic kits, Cairo, Egypt. The equation LDL-c = TC– (HDL-c) –TG/5 was used to calculate low density lipoprotein cholesterol (LDL-c). 18
Western blot analysis identifying the protein expression of p-38 MPAK, & SMAD2/3.
After homogenizing pancreatic tissue using RIPA buffer, it was centrifuged at (23,182 g) for 20 min at 4°C. Protein content was determined using BCA kit. Eight percent in each sample were overloaded by sodium sulfate-polyacrylamine electrophoresis (PAGE), transferred to the nitrocellulose membrane using semidry transmission equipment (Bio-Rad, Hercules, CA, USA) (AMERSHAM BIOSCIENCE, Piscataway, NJ, USA). Then they were incubated with 5% TBST milk block and washed with a 1:1000 dilution of anti-p-38 MPAK, anti-p-SMAD2/3 overnight on a roller shaker at 4°C. The filters were then rinsed and tested for immunoglobulin conjugated with horseradish peroxidase from Amersham Life Science, Little Chalfont, United Kingdom. The quantity of protein studied was measured by densitometric analysis using a densimeter used for laser scanning (Biomed Instrument Inc., USA). After normalization with household protein, β-actin values were provided.
PKH26 labeling stem cells
BM-MSCs were labeled with PKH26 (Sigma-Aldrich, Saint Louis, MO). Cells were first centrifuged, washed twice in serum-free medium, and then pelleted and suspended in dye solution. Cells were injected intravenously into rat tail vain. Pancreatic tissues were examined with a fluorescence microscope to detect and trace the cells. Identification and tracking of cells stained with PkH26 in pancreatic tissues were examined with a fluorescence microscope, Olympus BX50F4, no. 7M03285, Tokyo, Japan.
BM‐MSC differentiation assays
• Adipogenesis assay: Insulin (5 g/mL) and dexamethasone (10-9 M) were used to incubate culture cells for 21 days. Using Oil Red O staining, the accumulation of lipid droplets in vacuoles was seen. As established by phase-contrast imaging, the presence of highly refractive intracellular lipid vacuoles suggests adipogenic differentiation.
19
• Chondrogenesis assay: The TGF-β1 (1 ng/mL) and (50 g/mL) ascorbic acid 2-phosphate were added to a 500 L chondrogenic medium. We collected the cells after 21 days, fixed them in paraffin, cut them into 3 m slices, and stained them with 0.1% Alchian blue for glycosaminoglycans.
20
• Osteogenesis assay: Over the course of a 21-day period, cell cultures were supplemented twice per week with 50 g/mL ascorbic acid 2-phosphates and dexamethasone in concentrations of 109M. For the next 20 min, the cells were fixed in 10% formalin at room temperature.
21
Alizarin Red staining at 2% wt/vol for 20 min, pH adjusted to 4.1 with ammonium hydroxide, revealed the presence of calcium-rich hydroxyapatite in the ECM.
20
Histopathological examinations
In 10% neutral formalin buffer, pancreas samples were immersed. Fixed specimens were subsequently cleaned in xylene, integrated into paraffin, cut at 4–6 U thickness and stained with hematoxylin and eosin. 22 The pancreas alternation parameters were graded as: inflammatory infiltration cell (0 = negative, 1 = 5%, 2 = <50%, 3 = ≥50%), atrophy of acinar cells (0 = negative, 1 = slight, 2 = moderate, 3 = severe) and fibrosis (0 = absent, 2 = mild, 3 = spread).
Statistical analysis
The data were statistically evaluated, and significance testing was carried out via the one-way ANOVA test followed by a post-hoc test for several comparisons, using the statistical package SPSS 15.0 (Statistical Program for Social Sciences). The slandered error (SE) and the difference between the means are considered significant at p < 0.05. The mean of six values were taken for each data.
Results
MSCs morphology engraftment and cell surface MSC specific markers characterization
The MSCs have been excised from rats. Undifferentiated, isolated, and cultivated BM-MSCs achieved 70%–80% integration in 14 days. Adhesiveness and form of the BM-MSCs showed an equivalent heterogeneous fibroblast (lengthy spindles) morphology shown in (Figure 1(a)–(c)). The flow cytometry results showed that CD 105, CD 90, and CD29 were positively expressed and CD34 was negatively expressed. (A and B): Morphological fibroblast-like appearance of MSCs and cell surface MSC specific markers. Isolation and culture first day (a) cultured (MSCs) derived from rat were morphologically defined by a fibroblast-like appearance at passage one, two, and three, respectively (b, c) (Scale bar = 200 μm). The surface MSCs specific markers including CD 90 and CD29 was positive in isolated cells but negative for CD34, which is a hematopoietic stem cell marker. The violet field represents the IgG expression control isotype, while the black line depicts the expression marker.
BM-MSCs differentiation
Figure 2 showed that Alizarin Red stain of calcium deposits, Alchian blue stain of sulfated proteoglycans, and Oil Red stain of fat vacuole deposits indicated the differentiation ability of BM-MSCs toward osteoblasts, adipocytes, and chondrocyte at 14 and 21 days after differentiation, respectively The differentiation of MSCs. 1,2, and 3 indicate the differentiation into osteocytes, chondrocytes, and adipocytes, respectively.
The efficacy of BM-MSCs on pancreatic enzymes
To consider the impact of BM-MSCs on pancreas malfunction, the activities of lipase and amylase were evaluated after induction of CP in the serum of BM-MSCs–treated and untreated animals. As revealed in (Figure 3), CP resulted in pancreatic injury as demonstrated by the elevated enzymes compared to control, which was attenuated by BM-MSCs injection compared to CP. Effect of BM-MSCs on digestive enzymes (Lipase and Amylase) in all groups. Each value represents the mean of 6 values ±SE. The values with different superscripts are significantly different at p < 0.05 versus (a) control group, (b) CP group, (c) BM-MSCs group, and (d) CP + BM-MSCs group.
Effects of BM-MSCs on lipid profile
Concerning the lipid profile, administration of L-Arg (group 2) caused a significant augmentation (p < 0.05) in TC, TG, and LDL-c with reduction in HDL-c compared to control group. While BM-MSCs treatment twisted to a significant reduction (p < 0.05) in serum TC, TG, and LDL-c with a significant increase (p < 0.05) in HDL-c compared to the CP group (Figure 4). Effect of BM-MSCs on TC, TG, LDL-c, and HDL-c in all groups. Each value represents the mean of 6 values ±SE. The values with different superscripts are significantly different at p < 0.05 versus (a) control group, (b) CP group, (c) BM-MSCs group, and (d) CP + BM-MSCs group. TC: total cholesterols, TG: total glycerides, LDL-c: low density lipoprotein cholesterol, HDL-c: high density lipoprotein cholesterol.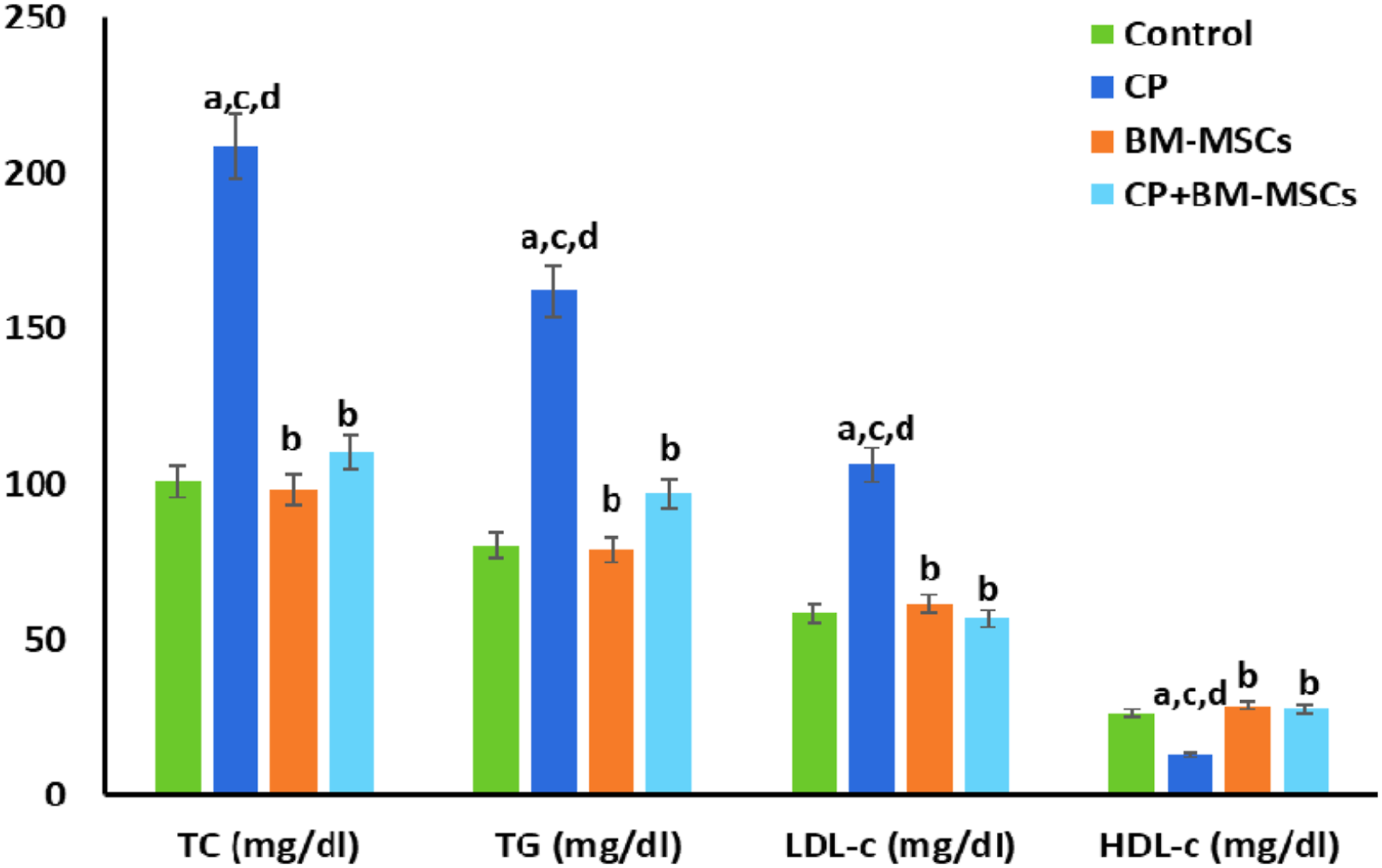
BM-MSCs regulate the PGE2 concentration and inflammatory markers mediated chronic pancreatitis
Assessment of BM-MSCs for anti-inflammatory activity is shown in Figure 5. The data revealed a highly significant dramatic increase in cytokine; IL-18, fibrogenic factor; TGF-1β, chemokine; MCP-1, and heme enzyme; MPO levels in the CP group compared to the corresponding control group. Meanwhile, data displayed a significant reduction (p < 0.01) in these inflammatory markers and growth factor TGF-B1 in BM-MSCs treated CP animals. As shown in Figure 5, compared with the normal group, the concentrations of CPR and PGE2 significantly increased in the serum of CP rats (p < 0.01). Treatment with stem cells significantly reduced the concentrations of PGE2 compared to CP rats (p < 0.05). Effect of BM-MSCs among all groups on inflammatory and fibrosis factors A) IL-18, B) TNF-α, C) TGF-β1, D) CRP, E) MCP-1, F) MPO, G) PGE2. Each value represents the mean of 6 values ±SE. The values with different superscripts are significantly different at p < 0.05 versus (a) control group, (b) CP group, (c) BM-MSCs group, and (d) CP + BM-MSCs group. IL-18: interleukin-18, TNF-α: tumor necrosis factor alpha, TGF-β1: transforming growth factor beta 1, CRP: c reactive protein, MCP-1: monocyte chemotactic protein-1, MPO: Myeloperoxidase, PGE2: prostaglandin.
The influence of BM-MSCs on the protein expression of p-38 MPAK and SMAD 2/3 in pancreatic tissue
Rats with chronic pancreatitis manifested a significant increase in the protein expression of p-38 MPAK, SMAD 2/3 with respect to the normal control one. Treatment of CP rats with BM-MSCs induced a regulation in the p-38 MPAK, p-SMAD2/3 protein expression compared to CP group as illustrated in (Figure 6(a)–(c)). The capability of BM-MSCs to transform into pancreatic lineage cells, which agreed with our results was found as upper expression in p-38 MPAK, p-SMAD2/3 levels in the CP + BM-MSCs group when compared to the corresponding values in the BM-MSCs group (Figure 6(a)–(c)). Signaling fibrosis factors by Western Blot analysis: phospho-protein expressions of both A) P-SMAD2/3 and B) p-38 MAPK and (C) PAGE of p-38MAPK/t-p-38MAPK and p-SMAD2/t-SMAD⅔ protein expression normalized to β-actin of pancreas tissues. Each value represents the mean of 6 values ±SE. The values with different superscripts are significantly different at p < 0.05 versus (a) control group, (b) CP group, (c) BM-MSCs group, and (d) CP + BM-MSCs group.
BM-MSCs homing
The homing of red fluorescent-labeled MSCs in the treatment groups is represented in Figure 7. A large number of significant fluorescent MSCs was observed in the BM-MSCs group (Figure 7(c)) and the number was greatest in the immunofluorescent PKH-26–labeled MSCs in the pancreas tissue of the CP + BM-MSCs group, whereas both the control (Figure 7(a)) and the CP (Figure 7(b)) groups were negative for the fluorescent dye. BM-MSCs stained with PKH26 and their relocation assess inside the wounded islets. The (a) control, (b) CP, (c) BM-MSCs, and (d) CP + BM-MSCs groups images were taken with a florescent microscope to localize the PKH26-labeled cells. (Scale bar = 100 μm).
Histopathological results
Histopathological examination of pancreas tissue showed that control and BM-MSCs groups in Figure 8(a) and (b) have typical histological structure that includes both endocrine and exocrine structures as grade (0). However, rats treated by the L-Arg (CP) group demonstrated abnormalities in pancreas characterized by pancreatic lobule atrophy, vacuolation of the acinar epithelial lining, and loss of some Langerhans cells compared with the control group. Variable degrees of degenerative changes in pancreatic acini with loss of normal lobular architecture were seen. Some acinar cells showed vacuolation with pyknosis of their nuclei, reduced basal basophilia and mild fibrosis with focal mononuclear cell aggregation, mainly lymphocytes and macrophages, were noticed. Moreover, apoptosis of Langerhans’ cells appeared as eosinophilic apoptotic bodies in-between cells with grade 3 (Figure 8(a), (c)–(f)). Nevertheless, CP + BM-MSCs treated group (Figure 8(a), (g), (h)) showed an improvement in the pancreatic Langerhans histological picture. (A and B): Pancreatic tissue section showing (a and b) normal histological structure of both exocrine (ex) and endocrine tissues (endo), (c) loss of lobular architecture with vacuolation of acinar cells arrow and interlobular fibrosis arrowhead, (d) loss of some Langerhan’s cells (Lc) with mild fibrosis arrow, (e) apoptosis of Langerhan’s cells arrow, (f) vacuolation, (v) of acinar cells with focal mononuclear cells aggregation arrow, (f) good improvement of pancreatic lobules and islets of Langerhan’s (IL), (g) Hyperplastic proliferation of Langerhan’s cells (Lc) (H and E ×200). B): Chronic pancreatitis histological grading.
Discussion
The impact of CP is steadily growing and the fundamental etiopathogenesis process is not established. Because progenitor cells, such as beta-cells, constitute the primary foundation of pancreatic cells, proliferation, and differentiation into islet-like cell clusters pave the way for islet renewal. A possible therapy that induces differentiation into pancreatic progenitor cells may be a useful strategy for beta cell curative. 23 We examined the effectiveness of BM-MSCs in the current work as a renovation into acinar as well as cells in the rat model of L-arginine. Autotarget of MSCs is a mechanism that explains the relocation of injured cells in response to interaction between transcription factors released from damaged tissues and receptors expressed on the surface of MSCs. 24 After damage, MSCs preferably go home to sites of inflammation where they spread through the endothelium and enter the wounded tissue bed. 24
The destruction and exchange of acinar tissue-distinctive chronic pancreatitis involved signaling pathways with severe inflammation induced by L-arginine as a co-factor for the fibrosis that is confirmed by pancreas tissue histopathology. Our data also denoted a marked activation of digestive enzymes as a significant decrease in pancreatic functioning accompanied by a high level of IL-18, TNF-α, and CRP cytokines activated through immunoregulatory factor PGE2 up-regulation. As well, an elevation of p-38 MPAK and SMAD2/3 protein expression levels was observed besides a significant increase in secreted TGF-β levels, along with marked elevated of MCP-1 which is a critical chemokine in pancreatitis pathophysiology.
Overexpressed PGE2 is a marker in the pancreatic ductal and acini of chronic pancreatitis patients. PGE2 is a powerful inflammatory intermediate and has a link to pancreatic stellate cell profibrotic activity. This agrees with earlier findings, coordinating a set of circumstances that cause damage to many biological components, such as DNA, lipids, and proteins. 25 L-Arg exposure induced tissue damage which is characterized by excessive fundamental signals through inducting reactive oxygen species overproduction and activating the inflammatory mediators’ responses to organ injuries.26,27 Repeated and accumulated doses of L-Arg raise the pro-inflammatory level, which is associated with prostaglandin release and the generation of free radicals, nitrosative stress, which is known as a substrate for synthase NO.
Under these circumstances, such as L-Arg high doses, oxidative injury, severe inflammation, and its consequences, a disruption may occur in the functions of the pancreas, which clarifies increase in lipase and amylase enzymes in CP rats. The most dramatic result of this research is that a single infusion of BM-MCSs restored lipid profile levels in CP rats. Supporting our study, the BM-MCSs interact with oxidative stress processes by chelating free radicals. In this study, the lipid profile is improved after treatment of CP rats with MSC. Ahmed et al. 28 clarified that the normalization of the lipid profile by BM-MSCs is caused by improving β-cell function and reducing insulin resistance. As well, the stroma can repair pancreas injuries which lead to differentiation into beta-cells by the secretion of a variation of bioactive factors by MSCs. 29
Studies indicate that injection of rat MSCs decreased IL-6 and TNF-α with inhibition of extracellular matrix excessive secretion and expression of MCP-1. 30 The Cascade performance in the TGF-β1 signaling pathway starts by stimulating PSCs and then activating cytokines TNF-α, IL-18, and MCP-1 by upregulating the activity of MAPK pathway. 31 Subsequently, Smads form a complex by TGF-β1 activation and enter the nucleus to stimulate transcription genes that promote the profibrotic pathway of pancreas. 32
Our findings were in line with these prior studies that explain how L-Arg induces persistent inflammatory reactions such as acinar cell necrosis, which is followed by the release of cytokines. Profibrogenic is the vital progression underlying the diagnosis of severe pancreatitis. L-arginine induction led to severe necrosis of ductal cells with massive edema, cellular infiltration, and interlobular fibrosis. Pancreatitis induced by L-arginine can be improved with treatments directed at ROS scavenging effects, leading to healing organs. Our results indicate that the therapeutic effectiveness of BM-MSCs could be through healing inflammatory property involved in chronic pancreatitis, which initiates the mobilization and differentiation of MSCs into the microenvironment and helps in repairing the tissue. 33
Meanwhile, TGF-β growth factors have main role in the regulation of SMAD signaling, 34 which is an essential pathway of osteogenic mesenchymal stromal cells. 32 Activating TGF-β1 mRNA by arginine was released from wounded pancreas tissue and induces rising pro-inflammatory cells and deposits of collagen, fibronectin, glial fibrillary acidic protein, and p-Smad3 activation. 32 Based on these results, we also studied the effect of BM-MSCs on TGF-β overexpression induced pancreatic stellate cell activation and extracellular matrix stimulation. It was demonstrated that BM-MSCs modulate the expression of TGF-β1 upon treatment, which suggests that BM-MSCs have a major role in fibrosis cure. Our study data showed that transplanted BM-MSCs suppressed AMP-kinase and inhibited the spontaneous activation of TGF- β1, it suggests that p38 MAPK leads to the activation of PSCs.
In vivo conditions, our data and others demonstrated a limited capacity for reconstituting marrow mesenchymal cells and differentiation. It can be caused by increased inflammations and excessive tissues stress of the host disease, which prohibits MSCs from progressing in chronic pancreatitis therapy. 35 In the current study, pancreatic tissue BM-MSCs marked with luminosity of PKH26 have been observed confirming that these cells are nested in the pancreas, indicating that BM-MSCs may be located in the wounded pancreas, distinguish into beta cells, and redevelop the vastly specific tissue of pancreas. Its spatial distribution occurs in the pancreas, as it is expressed and secreted within islets, that are delimited by several factors, such as inflammatory cytokines, oxidative stress, and β-cell damage. 36
Further researches have revealed that MSCs homed in the damaged liver tissue 37 have generated insulin in the pancreas of diabetic rats.38,39 It was previously reported2,7,27,32 that, MSCs were conceivable and the direct interaction have indicated chronic pancreatitis was relieved with MSC transfusion and improved pancreas impairment function. However, there are some limits to our study. For example, exploring the actual mechanism of TGF beta, by using inhibitors to distinguish its role through comparing it to controls, to fully explaining its internal mechanism with MSCs in treating chronic diseases.
Conclusions
Transplantation of BM-MSCs decreased the increment of pancreatic enzymes, cholesterol, triglycerides, and LDL-c with elevated HDL-c levels. In addition, it has an anti-inflammatory effect by secreting several kinds of bioactive molecules. This was confirmed by attenuation of IL-18, TNF-α, CRP, MPO, and PGE2 levels accompanied by regulating the TGF-B1/AMPK/SMAD signaling factors elevation with reduced MCP-1 levels. A limitation of our study is the lack of molecular (siRNAs) or genetic (KO-MSCs) approaches to confirm the role of TGF-B1/MAPK/SMAD in the study. Bioassay and molecular evaluations were supported by histopathological findings.
Footnotes
Acknowledgments
The authors would like to thank Dr Ahmed Othman (Professor of Histopathology, Faculty of Medicine, Cairo University) for carrying out blinded histopathological examination in this work.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Ethics approval
All ethical protocols for animal treatment were guided by the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 85, revised 1996) and were supervised by the Research Ethics Committee of the National Center for Radiation Research and Technology, Atomic Energy Authority (Ref. no. 202/21).
Animal welfare
The present study followed National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978) and supervised by the animal facilities, National Center for Radiation Research and Technology, Atomic Energy Authority.