
Abstract
Hepatic ischemia/reperfusion injury (IRI) is a clinical condition that may lead to cellular injury and organ dysfunction that can be observed in different conditions, such as trauma, shock, liver resection, and transplantation. Moderate levels of nitric oxide (NO) produced by the endothelial isoform of the NO synthase protect against liver IRI. GIT-27NO is a NO-derivative of the toll-like receptor 4 antagonist VGX-1027 that has been shown to possess both antineoplastic and immunomodulatory properties in vitro and in vivo. In this study, we have investigated the effects of this compound in vitro, in a model of oxidative stress induced in HepG2 cells by hydrogen peroxide (H2O2), and in vivo, in a rat model of IRI of the liver. GIT-27NO significantly counteracted the toxic effects induced by the H2O2 on the HepG2 cells and in vivo, GIT-27NO reduced the transaminase levels and the histological liver injury by reducing necrotic areas with preservation of viable tissue. These effects were almost similar to that of the positive control drug dimethyl fumarate. These data suggest that the beneficial effect of GIT-27NO in the hepatic IRI can be secondary to anti-oxidative effects and hepatocyte necrosis reduction probably mediated by NO release.
Introduction
Hepatic ischemia/reperfusion injury (IRI) is a condition that can occur in different settings, such as trauma, shock, liver resection, and transplantation1,2 and is followed by a period of reperfusion that revascularizates and reoxygenates the organ.3,4 Hepatic IRI may also affect multiple tissues and organs as this condition can seriously affect liver function, even producing irreversible damage, which leads to a cascade of multiple organ dysfunction.5–7 Several pathogenetic factors, including anaerobic metabolism, oxidative stress, mitochondrial damage, secretion of reactive oxygen species (ROS), intracellular Ca2+ overload, cytokines and chemokines produced by Kupffer cells and neutrophils, and nitric oxide (NO), have been proposed to contribute to the regulation of hepatic IRI processes. Matrix metalloproteinases can also be an important mediator of early leukocyte recruitment and target in acute and chronic liver injury associated with ischemia. 8
IRI can be classified into warm IRI, which primarily occurs at the level of the hepatocytes during liver resection surgery or during different types of low-flow states, and cold IRI, which usually damages sinusoidal endothelial cells and non-parenchymal cells and occurs after the hypothermic preservation of the graft during liver transplantation. 9
IRI of the liver is associated with marked increase in serum liver enzymes, such as alanine transaminase (ALT), aspartate transaminase (AST), lactate dehydrogenase, and/or serum glutamic-oxaloacetic transaminase and by histological assessment with the Suzuki classification. 10 The ischemic damage is worsened by reperfusion, resulting in local inflammation, cell death, tissue injury, and organ failure.3,4 Hepatic IRI may also influence the function of many other organs, such as the lung, kidney, intestine, pancreas, adrenals, and myocardium. 7 There are limited therapeutic options currently available for prevention and treatment of hepatic IRI that include pharmacological and genetic therapies and surgical interventions. 11
However, as these approaches have limited effects and none of them is so far approved as standard of care for treatment or prevention of hepatic IRI, new studies are warranted to identify new therapeutic approaches.
Another molecule that has attracted attention as pathogenetic factor and also as potential therapeutic target of hepatic IRI is NO, which is a highly reactive gaseous signaling molecule produced from the oxidation of L-arginine by the enzyme NO synthase (NOS). 12 In mammals, there are three different isoforms of NOS: neuronal NOS (nNOS, NOS I); endothelial NOS (eNOS, NOS III), which are constitutive and calcium-dependent isoforms; and inducible NOS (iNOS, NOS II), which is inducible and calcium-independent. 13 nNOS is mostly expressed in neural tissue and does not play a significant role in IRI of the liver. 12 However, eNOS, which is constitutively expressed in sinusoidal endothelial cells, and iNOS, which can be expressed by hepatocytes and immune cells in inflammatory conditions, are involved in this clinical condition. 14
It has been demonstrated that either endogenously produced or exogenously administered NO may exert positive or negative effects on the development of hepatic IRI, depending on the isoform that produces it, the concentration, the site of production, and the type of NO donor that is administered.10,12,15,16 For example, it has been demonstrated that moderate levels of NO produced by the eNOS isoform protect against liver IRI.10,12,15,16 However, the role of NO generated by iNOS is more controversial.10,12,15,16 High levels of NO generated by iNOS may have a negative effect on the course of the condition, interacting with superoxide anion and subsequently producing the peroxynitrite anion, which is a highly toxic molecule associated with pathological liver conditions and induces leukocyte adhesion, inflammatory cell infiltration and parenchyma cell dysfunction.15,16 However, another study indicates that iNOS-derived NO exhibits either positive or negative effects in livers without steatosis during the early phase of IRI, depending on the experimental model and on the length of ischemia and reperfusion and the protective action would occur primarily in the late phase of the condition. 12
A recent review investigated the roles of NO in pharmacological protection against hepatic IRI. In particular, it is recapitulated that NO may ameliorate liver IRI through multiple mechanisms, such as anti-inflammatory effects, reduction of oxidative stress, modulation of mitochondrial respiration and ROS reduction, cytoprotection, and regulation of the microcirculation. 17 NO may also prevent cell apoptosis inhibiting caspase activities. 8 As discussed above, modulation of endogenous NO in a timely and dose-adequate fashion may represent one of the possible approaches for prevention and/or treatment of hepatic IRI. Along this line, it has been shown that NO-producing drugs and inhaled NO may exert positive effects on IRI of the liver.8,18
Rodent models of hepatic IRI provide a useful tool to generate preclinical proof of concept studies that may be worthy of further investigation in the clinical setting.11,19
Along this line of research, we have evaluated the effects of another NO-releasing drug in the development of experimental hepatic IRI in a well-known rat model of the corresponding human disease. For this purpose, we have used (S,R)-3-phenyl-4,5-dihydro-5-isoxazole acetic acid–nitric oxide (GIT-27NO) that is a NO-producing agent generated from the covalent linkage of a NO moiety to the parental immunomodulatory and anti-inflammatory compound (S,R)-3-phenyl-4,5-dihydro-5-isoxazole acetic acid (VGX-1027). 20
Materials and methods
Reagents and cells
GIT-27NO and VGX-1027 were obtained as described elsewhere.21,22 While GIT-27NO was provided from Ganial Immunotherapeutics (Locust Valley, NY, USA), its parental compound VGX-1027 was a kind gift from VGX Pharmaceuticals (Blue Bell, PA, USA). GIT-27NO (MW = 328 g/mol) and VGX-1027 (MW = 205 g/mol) were stored at +4°C as a 20-mg/mL stock solution in 2.5% dimethyl sulfoxide (DMSO)–H2O and they were diluted in culture medium immediately before use. Dimethyl fumarate (DMF) was bought from Sigma–Aldrich and was stored at 4°C as a 100-mg/mL stock solution in DMSO and was diluted in culture medium immediately before use. Control cell cultures were treated with an adequate volume of diluted DMSO.
The HepG2 human hepatoblastoma cells23–25 were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA) and were maintained in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% fetal calf serum (FCS), 2.0 mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and incubated at 37°C and 5% CO2/95% air.
Bioinformatic analysis
In order to explore the molecular pathways involved in the liver IRI and to identify a suitable in vitro model, we interrogated the publicly available microarray datasets, GSE14951 and GSE47739, obtained from the Gene Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/gds). The GSE14951 dataset included whole-genome transcriptomic levels from donor livers at baseline and following transplantation–reperfusion. 26 The GSE47739 microarray dataset included data from HepG2 challenged for 60 min with hydrogen peroxide (H2O2) 0.5 mM. 27 Genes with adjusted P-value <0.05 and log (fold change) > 1 were considered for the analysis. Gene enrichment analysis was performed using the Enrichr software (https://amp.pharm.mssm.edu/Enrichr/). 28
Drug toxicity by MTT assay
Drug toxicity was evaluated using MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay after 5 h of cell exposure to three scalar concentrations of GIT-27NO (1000, 500, and 250 nM) and of the parental compound VGX-1027 (1000, 500, and 250 nM). DMF (5, 10, and 20 µM) 29 was considered as positive control drug. 30 Mitochondrial-dependent reduction of the tetrazolium salt MTT to colored formazan product reflects mitochondrial activity of cultured cells, thus evaluating cell respiration and cell viability. The conversion of MTT to formazan was monitored on an automated microplate reader at 570 nm.31,32 The results of MTT were expressed as percentage compared to the control value obtained in untreated cell cultures, which was arbitrarily set to 100%.
In vitro oxidative stress induced in HepG2 cells by H2O2
Cells were seeded at 2.5 × 104 cells/well in 96-well plate and were incubated overnight to ensure cell adhesion. The first experiment was performed adding different concentrations of H2O2 to the cells, in order to choose the most appropriate. In the second experiment, cells were treated with three scalar concentrations of GIT-27NO and VGX-1027 (1000, 500, and 250 nM) and with DMF (20, 10, and 5 µM) along with two selected doses of H2O2 (2.5 and 1.25 mM) and incubated for 5 h. Then, cell viability was assessed by MTT assay and transaminases were measured in the supernatants by a standard photometric assay using a bichromatic analyzer (Reflotron; Roche Diagnostics GmbH, Mannheim, Germany).
In vitro detection of IRI-related proteins
Cells were seeded at 2.5 × 104 cells/well in 24-well plate and were incubated overnight to ensure cell adhesion. Cells were treated with GIT-27NO, VGX-1027 at the dose of 1000 nM, and with DMF (20 µM) along with H2O2 at the dose of 1.25 mM and incubated for 5 h. At the end of the experimental period, cells were collected for the isolation of proteins, using M-PER lysis buffer (Thermo Fisher Scientific, Waltham, MA, USA), following manufacturer’s instructions.
In the cell lysate, the levels of CDKN1A, DUSP5, PMAIP1TP53INP1, VCAN, and c-JUN were measured by enzyme-linked immunosorbent assay (ELISA) test using commercially available kits (Abcam, Cambridge, UK; LifeSpan BioSciences, Seattle, WA, USA).
Animals
Eight-week-old male Wistar rats weighing 200–220 g were purchased from Harlan Laboratories (Udine, Italy). The rats were kept under standard laboratory conditions (non-specific pathogen free) with free access to food and water and were allowed to adapt to their environment 1 week before the beginning of the study. The protocol was first approved by the local ethical committee OPBA (Prot. No. 246) and then by the Ministry of Health (Ministerial Authorization No. 31/2016-PR, 14/01/2016). Protection of animals used in the experiment is in accordance with Directive 2010/63/UE (22), enforced by the Italian D.L. No. 26 of 4 March 2014.
In vivo model for hepatic IRI: experimental procedure
The animals were anesthetized with a single intraperitoneal administration (i.p) of Zoletil 100 at a dose of 50 mg/kg in combination with an intramuscular administration of Dexdomitor at a dose of 40 µg/kg. 33 These doses were optimized in our animal house by the veterinary for our aim. Before each injection, the anesthetic cream EMLA at a dose of 2% was applied to the skin, 1 h before the anesthesia injection, according to the veterinary indications.
After cleansing the skin with betadine soap and alcohol, a midline incision was performed to open the abdomen and to expose the abdominal content, which was then covered with a piece of sterile gauze previously humidified to avoid dryness. After that, using two dampened cotton swabs, the median and left lateral lobes were raised against the diaphragm to better identify the portal triad (portal vein, hepatic artery, and biliary artery), which was clamped for 30 min (ischemic phase).
Then, clamps were removed and the animals were observed for 3 h (reperfusion phase) and after that were sacrificed. During the surgery phase, the animals were subjected to assisted ventilation to avoid a possible respiratory depression induced by the deep anesthesia.
Four groups of rats (9–10 rats for each experimental group) were considered. The first group of rats was treated with i.p according to a prophylactic regimen (3 days before and until 1 h before ischemia) with a dose of GIT-27NO of 50 mg/kg. This dose of GIT-27NO was chosen on the basis of previous studies in vivo in rodent models of tumors and immunoinflammatory hepatitis.20,21,32,34 The second group of rats was treated with the parental compound VGX-1027 at the same dose of GIT-27NO. This dose of VGX-1027 was also chosen on the basis of a previous study indicating this as golden dose capable of ameliorating several murine models of immunoinflammatory and autoimmune diseases. 33
Two control groups were included: a group of rats treated with DMF per os at the dose of 25 mg/kg 30 twice a day starting 3 days before and until 1 h before ischemia (positive control); a group of rats that underwent ischemic intervention and were treated with vehicle (negative control). A group of sham-operated rats was also included.
Rats were sacrificed by 20% CO2 inhalation, using a flow rate of 1 L/min CO2, for a total of 10 min. Euthanasia was confirmed by checking for the absence of heartbeat for 2 min. Each procedure was supervised by an expert veterinary. After laparotomy, blood samples from the inferior vena cava were drawn for the evaluation of transaminases by a standard photometric assay using a bichromatic analyzer (Reflotron; Roche Diagnostics) and the liver was excised and stored for the histopathological study.
Histopathological evaluation
Portions of livers were fixed in 10% neutral buffered formalin solutions for 24 h. Standard histopathological techniques were followed for processing the tissue and preparation of paraffin blocks. Fibrosis, necrosis, and inflammation changes were evaluated in sections (5 μm thick) stained with hematoxylin and eosin. The following scores were used to quantify fibrosis: 0, no fibrosis; 1, fibrous expansion of portal areas with or without fibrous septa; 2, fibrous expansion of portal areas with portal bridging; and 3, marked bridging with nodules (cirrhosis). Inflammation was scored as follows: 0, no inflammation; 1, periportal inflammation; 2, mild-to-moderate portal inflammation; and 3, severe portal inflammation. Necrosis was scored as follows: 0, no necrosis; 1, focal necrosis; 2, periportal or periseptal interface hepatitis; and 3, confluent necrosis. The final score for each rat was obtained by summing the scores of fibrosis, inflammation, and necrosis.
Statistical analysis
Statistical analysis for significant differences was performed according to analysis of variance (ANOVA) or t-test using GraphPad Prism 5.0 software. A P-value less than 0.05 was considered to be statistically significant. The data are presented as mean ± standard deviation (SD) of three independent experiments.
Results
Identification of liver IRI–related molecular pathways
For the identification of the molecular pathways involved in liver IRI and of a suitable in vitro model, the GSE14951 and GSE47739 datasets were analyzed and the differentially expressed genes were subjected to a Venn diagram analysis. In total, 508 and 386 genes were found to be differentially upregulated following liver ischemia–reperfusion (GSE14951) and upon H2O2 treatment of HepG2 cells (GSE47739), respectively; 31 differentially modulated genes were found to be common between the two datasets, entailing a strong statistical significance (P < 0.0001) (Figure 1(a)). A gene network showing the functional associations among the 31 overlapping genes is presented in Figure 1(b). Pathway analysis revealed that the top 3 significantly enriched pathways among the 31 overlapping genes were as follows: “Direct p53 effectors,” “ATF-2 transcription factor network,” and “HIF-1-alpha transcription factor network” (Figure 1(c) shows a network of the pathways enriched by the 31 genes overlapping the GSE14951 and GSE47739 microarray datasets). In particular, six genes belonged to the “Direct p53 effectors”: CDKN1A, DUSP5, PMAIP1, TP53INP1, VCAN, and c-JUN. Overall, this analysis suggests that the oxidative stress model induced in HepG2 cells by H2O2 can be used as an in vitro model for liver IRI.

Common regulatory pathways in liver ischemia–reperfusion injury and in H2O2 challenged HepG2 cells. (a) Venn diagram analysis of the genes commonly upregulated following ischemia–reperfusion injury and in H2O2 challenged HepG2 cells, as determined in the publicly available GSE14951 and GSE47739 microarray datasets. (b) Gene network constructed on the web-based utility STRING (https://string-db.org/) for the 31 genes in common between the GSE14951 and GSE47739 microarray datasets. (c) Network of the pathways enriched by the genes overlapping the GSE14951 and GSE47739 microarray datasets.
Effects of GIT-27NO on the in vitro oxidative stress model induced in HepG2 cells by H2O2
To evaluate the effects of GIT-27NO on oxidative stress in an in vitro model, HepG2 cells were incubated with H2O2 for 5 h in the presence of scalar concentrations of GIT-27NO and cell viability was measured by MTT assay.
The effects of GIT-27NO and of the parental compound VGX-1027 were compared to DMF, a potent anti-oxidant that significantly ameliorates the liver damage caused by IRI. 30
Preliminary studies were performed to ascertain the absence of drug toxicity after 5 h of incubation with HepG2 cells and no toxic effects were observed at all the tested doses of all the test compounds (data not shown).
The addition of H2O2 to the HepG2 cells induced a dose-dependent toxicity, which was significant (P < 0.0001) for the two highest doses (Figure 2). The doses that induced 90% and 50% of cell death (2.5 and 1.25 mM, respectively) were chosen for the experiment with the test compounds.
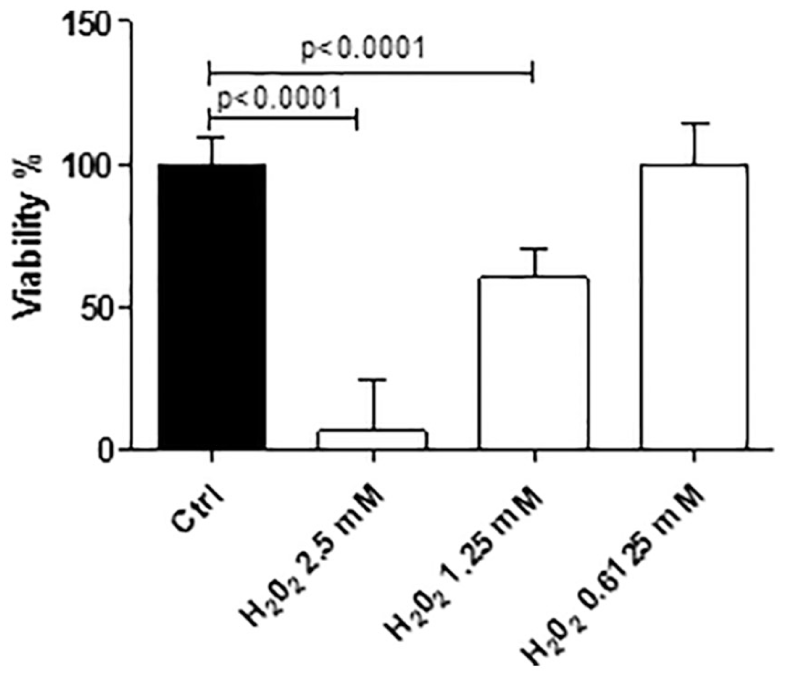
Effects of scalar doses of H2O2 incubated for 5 h with HepG2 cells. The data are presented as percentage of control ± standard deviation (SD) from three independent experiments.
The HepG2 cells were treated with three scalar concentrations of the test compounds along with the selected doses of H2O2. While GIT-27NO and its parental compound VGX-1027 were added at 1000, 500, and 250 nM, DMF was applied at 20, 10, and 5 µM.
The addition of all the test compounds to the cells in the presence of H2O2 at the highest dose of 2.5 mM was not able to counteract the strong toxicity induced by H2O2 (data not shown).
The addition of GIT-27NO, as well as of the reference compound DMF, to the cells in the presence of H2O2 at the dose of 1.25 mM significantly counteracted at the toxic effects induced by the H2O2 alone in all the tested doses and in a dose-dependent manner (P < 0.0005 for all test conditions) (Figure 3(a) and (c)). A significant reduction in the toxicity was observed upon the treatment with the parental compound VGX-1027 only with the highest dose of 1000 nM (P < 0.005) (Figure 3(b)).
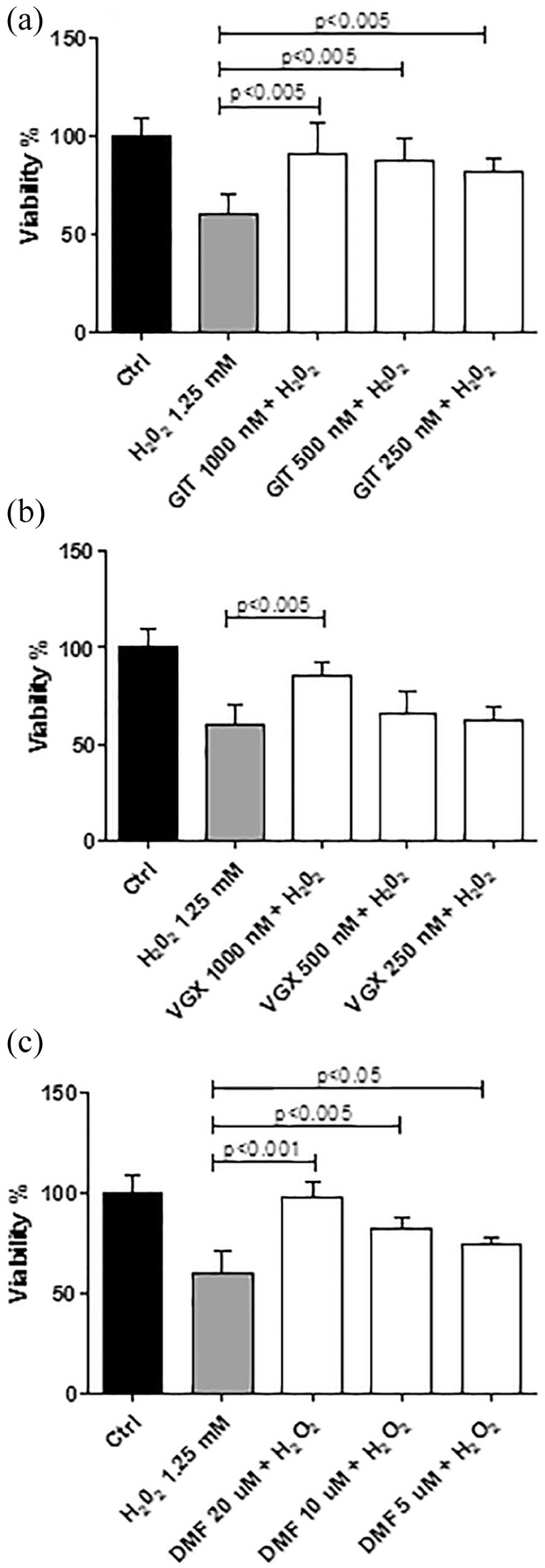
Effects of (a) GIT-27NO, (b) VGX-1027, and (c) DMF on the toxicity induced by H2O2 at the dose of 1.25 mM in HepG2 cells. Cell viability was assessed by MTT assay. The data are presented as percentage of control ± SD from three independent experiments.
AST levels were measured in the supernatants after co-treatment with H2O2 (1.25 mM) and the highest doses of the test compounds (GIT-27NO and VGX-1027 at 1000 nM and DMF at 20 µM). H2O2 induced a significant increase in AST concentrations compared to the control sample (P < 0.01). Treatment with GIT-27NO was able to significantly (P < 0.05) reduce AST levels, to levels comparable to those obtained with DMF (Figure 4).
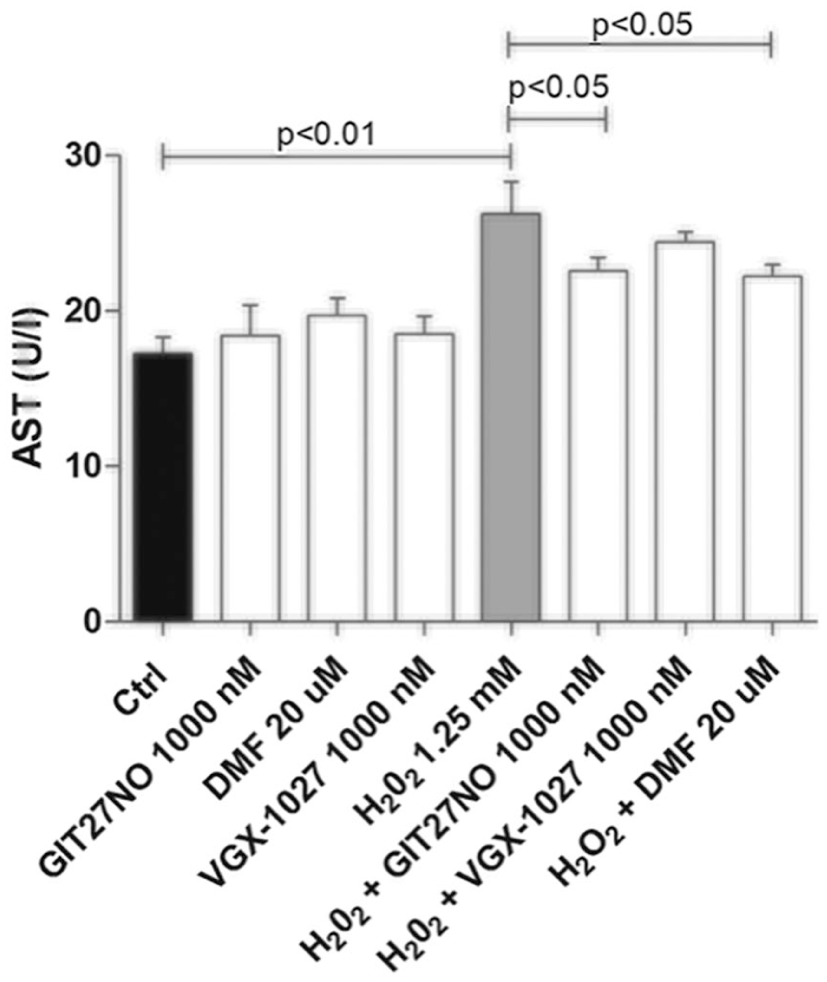
Effects of GIT-27NO, VGX-1027, and DMF on the AST levels induced by H2O2 at the dose of 1.25 mM in HepG2 cells after 5 h of incubation. The data are presented as means ± SD from three independent experiments.
Effects of GIT-27NO on IRI-related proteins
In order to investigate the mechanisms involved in the protective effects of GIT-27NO on oxidative stress model in vitro, the levels of the proteins previously identified by the bioinformatic analysis were evaluated upon treatment of H2O2 exposed cell with either 1000 nM GIT-27NO or the parental compound VGX-1027. The genes belonging to the top enriched pathway, the “Direct p53 effectors” pathway (Figure 1(c)), were chosen for the analysis (i.e. CDKN1A, DUSP5, PMAIP1, TP53INP1, VCAN, and c-JUN).
As shown in Figure 5, the incubation of cells with H2O2 significantly increased the expression of all the investigated proteins (P < 0.001 vs control cells). GIT-27NO treatment was associated with a significant reduction in the levels of the six analyzed proteins, entailing an effect comparable to that of DMF (Figure 5). Although the parental compound VGX-1027 significantly reduced the levels of PMAIP1 (P < 0.05), VCAN (P < 0.05), and TP53INP1 (P < 0.01), only a trend of reduction was observed for CDKN1A, DUSP5, and c-JUN (Figure 5).

Effects of GIT-27NO (1000 nM), VGX-1027 (1000 nM), and DMF (20 µM) on the levels of CDKN1A, DUSP5, PMAIP1, TP53INP1, VCAN, and c-JUN in HepG2 cell lysates after incubation with H2O2 (1.25 mM). The data are presented as means ± SD from three independent experiments.
Effects of GIT-27NO on the in vivo model of hepatic IRI
The positive data obtained in the in vitro study prompted us to evaluate the effects of GIT-27NO in vivo in a surgery model of hepatic IRI induced in rats. After the end of reperfusion, blood was collected from the cava vein and serum ALT was measured using an automated chemical analyzer. As expected, in vehicle-treated IRI rats, the plasma level of serum liver damage marker ALT was markedly increased compared to sham-operated rats (31 ± 5 U/L) and the treatment with DMF significantly (P < 0.001) counteracts this effect. A trend to the reduction of ALT plasma levels was observed in the group of rats treated with VGX-1027. However, the treatment with GIT-27NO induced a significant (P < 0.01) reduction of ALT as compared to vehicle-treated rats (Figure 6). These results confirm the effects obtained in vitro where GIT-27NO was able to reduce supernatant transaminase levels induced by H2O2 in HepG2 cells.

Effects of GIT-27NO, VGX-1027, and DMF on ALT levels. After the 3-h reperfusion phase, rats were sacrificed through CO2 inhalation and plasma was collected from cava vein for ALT dosage by a standard photometric assay using a bichromatic analyzer. The data are presented as means ± SD from three independent experiments.
In addition, semi-quantitative scoring of histopathological data showed that treatment with GIT-27NO resulted in a significant reduction of liver injury score of IRI rats as compared to vehicle-treated IRI rats (P < 0.0001) (Figure 7; Table 1).
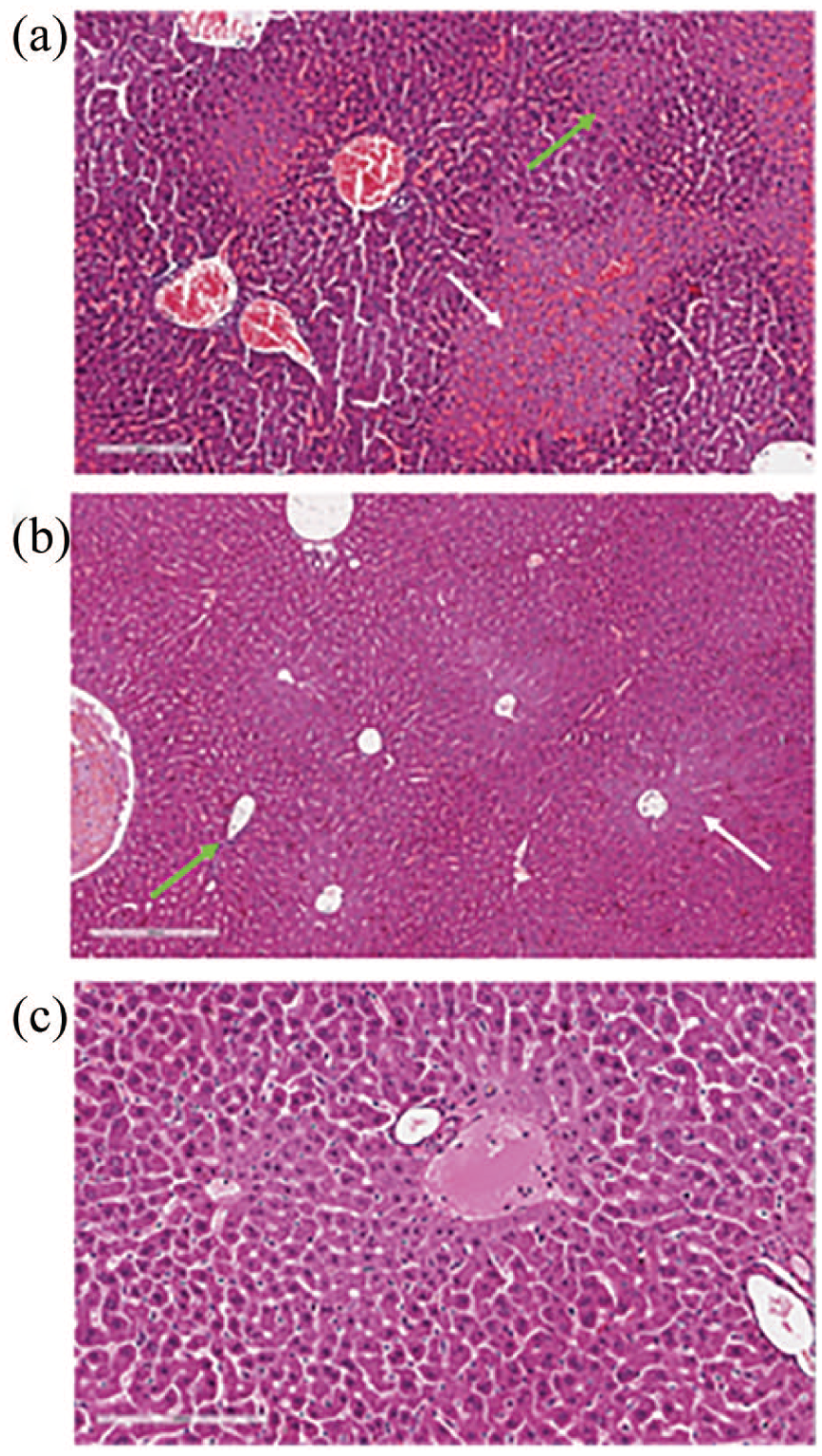
Effects of GIT-27NO on liver histological score in a rat model of IRI. Liver sections were stained with hematoxylin and eosin. (a) Rat treated with vehicle (H&E ×100); (b) rat treated with GIT-27NO (H&E ×100); (c) sham-operated rat (H&E ×150). Histological score: (a) fibrosis, 0; inflammation, 3 (green arrow); necrosis, 3 (white arrow); (b) fibrosis, 0; inflammation, 1 (green arrow); necrosis, 1 (white arrow); and (c) fibrosis, 0; inflammation, 0; necrosis, 0.
Effects of GIT-27NO, VGX-1027, and DMF on liver histological score in a rat model of hepatic IRI.

DMF: dimethyl fumarate; IRI: ischemia/reperfusion injury.
P < 0.001; **P < 0.0001 by t-test.
In particular, treatment with GIT-27NO significantly reduced necrotic areas with preservation of viable tissue. Also, the inflammation score was significantly reduced in treated rats. As reported in the figure, the inflammation was slight and circumscribed to periportal spaces instead of the severe portal inflammation observed in vehicle-treated rats. The present effects were similar to that one obtained with the positive control drug DMF (Table 1).
As reported in Figure 7, the GIT-27NO-treated rats compared to vehicle-treated IRI rats showed a significant reduction in sinusoidal congestion (score 2 vs 4), cytoplasmic vacuolization (score 1 vs 3), and necrosis (score 2 vs 4).
Discussion
GIT-27NO, differently from the parental compound VGX-1027, exerts powerful antitumor properties both upon in vitro and in vivo conditions.20,21,31,32,34 In particular, GIT-27NO is effective against numerous rodent and human tumor cell lines from different origin.20,21,31,32,34 However, GIT-27NO has also been shown to possess immunomodulatory properties as it is capable to suppress clinical, serological, and histological signs of murine immunoinflammatory hepatitis induced by injection of ConA which is a T-cell-mediated form of immunoinflammatory hepatitis that may serve as a model of human autoimmune hepatitis. 32 This study indicates that GIT-27NO may be useful for hepatic immunoinflammatory events, including IRI. Previous studies have demonstrated the protective effects of exogenous NO on hepatic IRI.35–37 A recent review indicates that NO might exert these protective effects by several mechanisms that entail increasing NO bioavailability, downregulation of p53 gene expression, reduced production of inflammatory chemokines, and ROS by activation of sGC-GTP-cGMP signal pathway that may reduce liver cell apoptosis and regulate hepatic immune functions. 17 Inhibition of caspase activity by the released NO through nitrosylation of cysteine residuals, as it has been demonstrated with NO-donating compounds in experimental models of immunoinflammatory hepatitis, could have also been implicated in the protection against hepatic IRI afforded by GIT-27NO in this study via possible reduction of apoptosis of liver cells.38,39
In agreement with these observations, we presently demonstrated that the NO-releaser GIT-27NO significantly counteracted the toxic effects and reduced the AST levels induced by the H2O2 on the HepG2 cells and reduced the signs of histological liver injury and serum ALT levels in a rat model of hepatic IRI. In particular, we have demonstrated in vitro that GIT-27NO significantly modulated the molecular pathway associated with liver IRI, identified by the bioinformatic analysis. The use of high-throughput genome data has allowed the uncovering of novel etiopathogenic pathways and molecular targets in different setting, such as autoimmune disease,40–43 cancer,44–47 fibrotic diseases, 48 and neurodegenerative disorders. 49 As the parental compound VGX-1027 only induced a mild reduction of the parameters considered both in vitro and in vivo, it seems possible that the strong protection obtained with GIT-27NO that was almost similar to that of the positive control drug DMF might indicate a synergistic effect between VGX-1027 and NO released by the compound. It should also be noted that VGX-1027 is a toll-like receptor 4 (TLR4) antagonist and signaling through TLR4 plays a key role in the development of experimental hepatic IRI.33,50
Overall, our study shows that, as compared to its parental compound VGX-1027, GIT-27NO is able to exert both in vitro and in vivo a stronger protective effect against hepatic oxidative stress. It is of interest for the potential clinical translation of these findings to the clinical setting that, as previously demonstrated in vitro, 21 unlike other exogenous donors of NO, GIT-27NO is only capable of releasing NO in the presence of cells. If this also occurs in vivo, GIT-27NO could be a more tailored NO donor that releases NO at the level of the organs and cells avoiding common unwanted side-effects related to immediate NO release in the circulation such as it occurs with the use of nitroglycerin for angina and that includes headache, weakness, dizziness, light-headedness, nausea, and flushing. Overall, these data suggest that the beneficial effects exerted by GIT-27NO in the hepatic IRI may be secondary to its ability to release NO and by doing so to interfere with anti-oxidative effect and hepatocyte necrosis reduction possibly in synergy with TLR4 antagonism and modulation of cytokine production afforded by the VGX-1027 scaffold. GIT-27NO has been shown to have a good therapeutic window in vivo as lethal dose of the compound is 160 times higher than that required to exert its pharmacological efficacy in the hepatic IRI model. 21
Along with the present data and its ability to prevent ConA-induced immunoinflammatory hepatitis, 32 these data suggest that GIT-27NO may represent a valuable drug candidate for treatment of several immune inflammatory events of the liver such as those that occur during IRI.
Footnotes
Availability of data and materials
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.