
Abstract
Dendritic cell (DC) activation induces expression of co-stimulatory surface molecules, as well as migration into secondary lymphoid organs, where they activate naïve T-cells. A family of plant derivatives, eremophilane-type petasite sesquiterpenes, can regulate the immune system through DC targeting due to their anti-inflammatory effects. Peroxisome proliferator–activated receptor gamma (PPARγ) is involved in inhibition of inflammatory responses and induction of DCs to acquire a mucosal phenotype. Since mucosal DCs are central in innate immune responses, we hypothesized that eremophilane-type petasite sesquiterpenes exerted their anti-inflammatory effects by inhibiting DC maturation and activation through PPARγ. This study assessed the bicyclic eremophilane-type petasite sesquiterpene compounds Fukinone and 10βH-8α,12-Epidioxyeremophil-7(11)-en-8β-ol (ZYFDC21 and ZYFDC22) in the maturation and activation of mouse DC. We measured surface expression of co-stimulatory molecules by flow cytometry and cell-free supernatant cytokine production upon lipopolysaccharide stimulation by enzyme-linked immunosorbent assays (ELISAs) in the presence or absence of PPARγ agonists. DCs were generated from C57BL/6 mice bone marrow cells and harvested. Cells were exposed to bicyclic eremophilane-type petasite sesquiterpenes ZYFDC21 or ZYFDC22 in the presence or absence of synthetic PPARγ agonists (GW1929 and TGZ) or the natural PPARγ ligand 15d-PGJ2, followed by overnight activation with LPS. We observed differences in the upregulation of surface expression of CD86, along with TNF, IL-6, and IL-12p70 released by DCs stimulated with LPS, when using combinations of bicyclic eremophilane-type petasite sesquiterpenes ZYFDC21 or ZYFDC22, and PPARγ agonists, in particular the PPARγ ligand 15d-PGJ2. Our results indicate that bicyclic eremophilane-type petasite sesquiterpenes ZYFDC21 or ZYFDC22 inhibit maturation and activation of DC, and this activity is augmented upon PPARγ activation.
Keywords
Background
Sesquiterpenes have been known to have anti-inflammatory activity in a variety of settings, showing inhibitory effects on nitric oxide production in lipopolysaccharide (LPS)-activated mouse macrophages.1–8 Some sesquiterpenes inhibit inflammation by targeting dendritic cell (DC) maturation and activation. For example, a sesquiterpene glycoside isolated from Kandelia candel inhibited pro-inflammatory cytokine production from LPS-stimulated bone marrow–derived DCs, 9 and micheliolide, a sesquiterpene lactone, inhibits the production of interleukin-6 (IL-6) and tumor necrosis factor (TNF) from LPS-stimulated primary DCs. 10 While some examples of the anti-inflammatory effects of sesquiterpene on DCs have been demonstrated, the molecular targets of specific sesquiterpenes and their interactions with endogenous inflammatory signaling pathways are unknown.
One possible target of sesquiterpenes in many inflammatory cells is the peroxisome proliferator–activated receptor (PPAR) pathway, which plays an important role in several cellular functions, including maturation and differentiation. PPARs were initially identified as receptors that controlled physiological responses to dietary intake of fatty acids.11,12 Three PPAR subtypes have been identified, alpha, delta and gamma, and are ligand-activated nuclear receptors which can be activated by polyunsaturated fatty acids, eicosanoids, and various synthetic ligands. PPAR gamma (PPARγ) is primarily expressed in adipose tissue and, to a lesser extent, in the colon, immune system, and the retina. PPARγ was first identified as a regulator of adipogenesis, but also plays an important role in cellular and adipocyte differentiation, insulin sensitization, glucose metabolism, atherosclerosis, and cancer. 13 It has been shown that PPARγ ligands have anti-inflammatory effects on mast cells, monocytes, macrophages, and DC, by modulating expression of co-stimulatory and adhesion molecules, altering their phenotype and leading to an impaired expression of pro-inflammatory cytokines/chemokine factors involved in T-cell activation and recruitment.14–18
Several sesquiterpenes or terpenoid-like compounds have been shown to either directly activate PPARγ or to modify its response to other ligands. For example, odoratin, an undecanortriterpenoid from Chromolaena odorata, moderately activates PPARγ; 19 tirotundin and tagitinin A, both sesquiterpene lactones, transactivate PPARγ-dependent promoters, including PPARγ response element (PPRE), small heterodimer partner (SHP), and ABCA1 gene promoters in dose-dependent manner, 20 and artemisinic acid, the quintessential sesquiterpene, reduces expression of PPARγ in human adipose tissue-derived mesenchymal stem cells. 21 Altogether, these data suggest that sesquiterpenes may similarly influence DC function through the PPARγ pathway. Recently, our group isolated two novel eremophilane-type sesquiterpene compounds from Petasites tatewakianus Kitam. 2 We hypothesized that these novel sesquiterpenes would inhibit DC maturation and activation, and that this activity would be augmented in the presence of a PPARγ agonist. In this study, we demonstrate, for the first time, that the novel bicyclic eremophilane-type petasite isolated sesquiterpenes have the ability to efficiently inhibit DC maturation and activation, and this inhibition is potentiatedby the synthetic, as well as naturally occurring, nuclear peroxisome proliferator-activated receptor γ agonists.
Materials and methods
Plant material
Bicyclic sesquiterpenes Fukinone (ZYFDC21), and 10βΗ-8α,12-Epidioxyeremophil-7(11)-en-8β-ol (ZYFDC22), were isolated and purified from rhizome of P. tatewakianus, at the School of Pharmacy, Shanghai University of Traditional Chinese Medicine as previously described. 22
Generation of bone marrow DCs from C57BL/6 mice
Female C57BL/6 mice (6–10 weeks old) were obtained from The Jackson Laboratory. All mice were treated according to protocols approved by the University of Alberta Animal Care and Use Committee. Bone marrow–derived DCs (BmDC) were generated using a standard protocol with little modification. 23 Briefly, bone marrow was flushed dispersed and collected from femurs and tibias of female C57BL/6 mice, passed through a 70 µm nylon mesh, and suspended in bone marrow–derived DC-complete media (RPMI 1640 containing 5 mM 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 50 U Pen/Strep, 2 mM glutamine, 50 µM 2-ME, 50 mM gentamycin sulfate, and 10% fetal bovine serum (FBS)) in the presence of granulocyte-macrophage colony-stimulating factor (GM-CSF) and IL-4 (10 ng/mL; PeproTech, Rocky Hill, NJ, USA) and cultured in tissue culture dishes (Thermo Fisher, Carlsbad, CA, USA) in a humidified atmosphere of 5% CO2 in air at 37oC. All media components, except for GM-CSF and IL-4, were obtained from Gibco (Carlsbad, CA, USA). During culture, half of the media was replaced on days 3 and 6. On day 8, BmDC were harvested, and their morphology was confirmed by optical microscopical analysis (Supplementary Figure 1).
Effect of sesquiterpenes and PPARγ agonists on BmDC
Initially, 0.2 × 106 BmDC/mL were deposited, per well, in a 12-well plate and incubated with either eremophilane-type petasite sesquiterpene, ZYFDC21 (50 µM) and ZYFDC22 (25 µM), in the presence or absence of synthetic PPARγ agonists troglitazone (TGZ, 5 µM or 10 µM; Sigma Aldrich Canada, Oakville, ON, Canada) or N-(2-benzoylphenyl)-O-(2-(methyl-2-pyridinylamino) ethyl)-
Flow cytometry of BmDC
After stimulation, 1 × 105 BmDC were incubated with their respective conjugated antibodies for 60 min at 4°C and washed twice. Data from 30,000 cells were collected by a CytoFlex flow cytometer (Beckman Coulter, Brea CA, USA) and VersaComp antibody capture beads (Beckman Coulter, Brea CA, USA) were included to generate a compensation matrix. Data analysis was performed using the FloJo V10 LLC software (Ashland, OR, USA). Gating was initially defined based on side scatter (SSC) versus forward scatter (FSC), BmDC positive gating was determined using an APC-labeled Armenian Hamster anti-mouse CD11c (BD Pharmingen, San Diego, CA, USA). CD11c+ subpopulation was then analyzed by the expression of CD80 and CD86 surface molecules with a FITC-Armenian Hamster IgG Anti-Mouse CD80 (Affymetrix eBioscience, Santa Clara, CA, USA) and APC-Rat anti-mouse CD86 antibodies (BD Pharmingen, San Diego, CA, USA) and compared to their respective isotype controls. Results were expressed as the median of fluorescence intensity (MFI) ± standard error of the median (SEM).
Cytokine release analysis
Levels of TNF, IL-6, and IL-12p70 released in the cell-free supernatants were quantified using commercial enzyme-linked immunosorbent assay (ELISA) according to the instructions of the ELISA Kits (Affymetrix eBiosciences). Results were expressed as means ± SEM.
Statistical analysis
Experiments were performed in triplicate, with BmDC obtained from at least three biological replicates (n ⩾ 3). Values are expressed as mean ± SEM. All statistical analyses were performed using GraphPad Prism statistical (GraphPad, Sand Diego, CA, USA). Statistical differences in the mean values among treatment groups were determined by using a one-way analysis of variance (ANOVA) test with post hoc analysis with Tukey’s multiple comparison tests. In all cases, a value for P < 0.05 was considered statistically significant.
Results
PPARγ activation inhibits DC maturation
In order to determine whether PPARγ agonists modified the maturation of DC, we first analyzed the surface expression of the maturation markers CD80 and CD86 on BmDC by flow cytometry. Figure 1(a) shows that BmDC exposed to different concentrations of synthetic PPARγ agonists TGZ (5 and 10 µM) or GW1929 (40 µM) presented CD86 MFI levels similar to control untreated cells; yet LPS stimulation induced upregulation of the co-stimulatory molecule CD86 on BmDC (MFI 31431 ± 7316, n = 5). Interestingly, when the BmDC were pretreated with the synthetic PPARγ agonists TGZ (5 or 10 µM) or GW1929 (40 µM) for 3 h followed by 20 h stimulation with LPS, there was a significant 75%–80% inhibition in the CD86 surface expression on BmDC compared to the LPS stimulation alone (MFI 6563 ± 1938 and MFI 5989 ± 2072 for TGZ 5 and 10 µM and 7459 ± 2317 for GW1929, respectively; n = 5). Expression of CD80- and CD86-positive BmDC after each treatment is shown in the supplementary Figure 3 as percentage values. We also examined the expression of CD80 after BmDC were exposed to the synthetic PPARγ agonists, followed by LPS overnight stimulation. We observed a 12%–20% inhibition in the expression of CD80 when cells were pretreated with TGZ or GW1929 (Figure 1(b)).

PPARγ activation inhibits DC maturation and cytokine secretion. CD86 surface expression in BmDC after 3 h preincubation with PPARγ agonists TGZ (5 and 10 µM), GW1929 (40 µM), and 15d-PGJ2 (0.5 and 5 µM) followed by LPS overnight stimulation was examined by flow cytometry. (a) CD86 results and (b) CD80 results are expressed as differences in MFI ± SEM between LPS activated-BmDC and PPARγ agonists ± LPS (n = 3–5; **P < 0.01 and ***P < 0.001). BmDC were incubated with PPARγ agonist TGZ (5 and 10 µM), GW1929 (40 µM), and 15d-PGJ2 (0.5 and 5 µM) for 3 h ± LPS overnight stimulation, and cell-free supernatants were collected and tested for (c) TNF, (d) IL-6, or (e) IL-12p70 release by ELISA. Results are from cytokines released from LPS-activated BmDC and cells treated with PPARγ agonists ± LPS. Data are expressed as means ± SEM (n = 3–5; *P < 0.05 and ***P < 0.001).
Natural PPARγ ligand 15d-PGJ2 modulates DC maturation
The cyclopentenone metabolite of PGJ2, 15d-PGJ2, is a naturally occurring derivative of prostaglandin D2 (PGD2) and has been shown to directly activate PPARγ24–26 BmDC were preincubated with 15d-PGJ2 (0.5 or 5 µM) for 3 h, and as shown in Figure 1(a), 15d-PGJ2 alone had no effect on the expression of CD86 at either of the concentrations tested. 15d-PGJ2 treatment for 3 h significantly decreased LPS-induced expression of CD86 by 60% and 50% (MFI 12,166 ± 1138 at 0.5 µM and 15147 ± 1376 at 5 µM; n = 3). CD80 surface expression did not reach statistical difference (9%–17% for 0.5 and 5 µM; Figure 1(b), n = 3). BmDC stimulated with LPS or complete media were included as positive and negative controls, respectively.
PPARγ activation promotes the inhibition of BmDC cytokine secretion. We analyzed the effects of the activation of PPARγ on the cytokine secretion of TNF, IL-6, and IL-12p70 released in the cell-free supernatant of BmDC, after 3 h treatment with TGZ (5 or 10 µM) or GW1929 (40 µM) by commercial ELISAs. As shown in Figure 1(c), pre-treatment with GW1929 significantly inhibited release of TNF (about 65% ± 5% compared to LPS). However, under the same conditions, BmDC release of IL-6 was unaffected by treatment with the PPARγ agonists, compared to LPS stimulation alone (Figure 1(d)). IL-12p70, the bioactive isoform of the cytokine, was also evaluated in the cell-free supernatants of BmDC exposed to 5 and 10 µM TGZ with and without LPS stimulation. We found that TGZ significantly inhibited (68% ± 1% and 66% ± 2%, respectively) IL-12p70 production, as shown in Figure 1(e).
PPARγ ligation skews BmDC cytokine response
We were interested in studying the response of BmDC to the treatment with the natural PPARγ ligand 15d-PGJ2 (0.5 and 5 µM), and we found that BmDC treated for 3 h with 15d-PGJ2 plus LPS inhibited TNF release by 29% ± 9% and 33% ± 9% at 0.5 and 5 µM, respectively; however, this inhibition was not statistically significant (Figure 1(c)). IL-12p70 showed a 33% ± 5% significant inhibition at 0.5 µM (Figure 4(c)).
Bicyclic petasite eremophilane-type sesquiterpenes potentiate the effects of PPARγ agonists on BmDC maturation and activation
Petasite sesquiterpenes have been shown to have anti-inflammatory activity in a variety of settings. We sought to assess the effects of two petasite eremophilane-type sesquiterpene compounds Fukinone (ZYFDC21) and 10βH-8α,12-Epidioxyeremophil-7(11)-en-8β-ol (ZYFDC22) isolated from the rhizome of P. tatewakianus on the maturation and activation of BmDCs. To evaluate the cytotoxic effects of the bicyclic compounds, we performed dose-response assays with several cell lines, using the XTT assay kit (Roche, data not shown). We selected sub-toxic doses of ZYFDC21 (50 µM) and ZYFDC22 (25 µM) and further evaluated their cytotoxic effects on BmDC after 1, 3, 24 and 48 h incubation, measuring viability by trypan blue exclusion (supplementary Figure 2). BmDC viability was ⩾95% under all tested conditions, and therefore, these concentrations were used for all experiments.
There is evidence that some sesquiterpenes exert anti-diabetic, anti-carcinogenic, and anti-inflammatory effects, mediated by the PPARγ pathway. 20 We sought to identify whether the sesquiterpenes would inhibit BmDC maturation and activation and whether this inhibitory activity would be augmented by the presence of a PPARγ synthetic agonist. For that purpose, BmDC were exposed to the synthetic PPARγ agonists TGZ (5 or 10 µM) or GW1929 (40 µM) in the presence or absence of the petasite sesquiterpenes ZYFDC21 (50 µM) or ZYFDC22 (25 µM) for 3 h, followed by the overnight LPS stimulation. First, we assessed the effects of bicyclic sesquiterpenes on BmDC maturation by flow cytometry. The presence of the sesquiterpenes ZYFDC21 (Figure 2(a)) and ZYFDC22 (Figure 3(a)) alone induced a modest increase in CD86 expression (MFI 6985 ± 1825 and 6882 ± 1274, respectively) compared to control, untreated BmDC (MFI 2073 ± 510). Exposure to a combination of ZYFDC21 (50 µM) plus the synthetic PPARγ agonist TGZ (5 or 10 µM) or GW1929 (40 µM) followed by overnight LPS stimulation resulted in a significant downregulation in CD86 surface expression (up to 80% compared to LPS levels alone; Figures 2(a) and 4(a)). The combination of ZYFDC22 (25 µM) with TGZ (5 or 10 µM) or GW1929 (40 µM) followed by overnight LPS activation resulted in a significant 85% and 80% inhibition, respectively, in CD86 surface expression (Figures 3(a) and 5(a)).

Bicyclic eremophilane-type petasite sesquiterpene ZYFDC21 potentiates the effects of PPARγ agonists on BmDC maturation and activation. BmDC were pretreated with the eremophilane-type sesquiterpene ZYFDC21 ± PPARγ synthetic agonist TGZ (5 or 10 µM) for 3 h, followed by LPS overnight stimulation, and DC were collected, fixed, and analyzed by flow cytometry. (a) Differences in the CD86 surface expression are represented as differences in MFI ± SEM between LPS activated-BmDC and the combination of ZYFDC21 + TGZ ± LPS (n = 5; ***P < 0.001). Cytokine release by BmDC treated with ZYFDC21 plus PPARγ agonist TGZ (5 and 10 µM) for 3 h ± LPS overnight stimulation, and cell-free supernatants were collected and tested for (b) TNF, (c) IL-6, or (d) IL-12p70 release by ELISA. Results are from cytokines released from LPS-activated BmDC, compared to BmDC cells treated with ZYFDC21, plus TGZ ± LPS. Data are expressed as means ± SEM (n = 5; *P < 0.05, **P < 0.01, and ***P < 0.001).
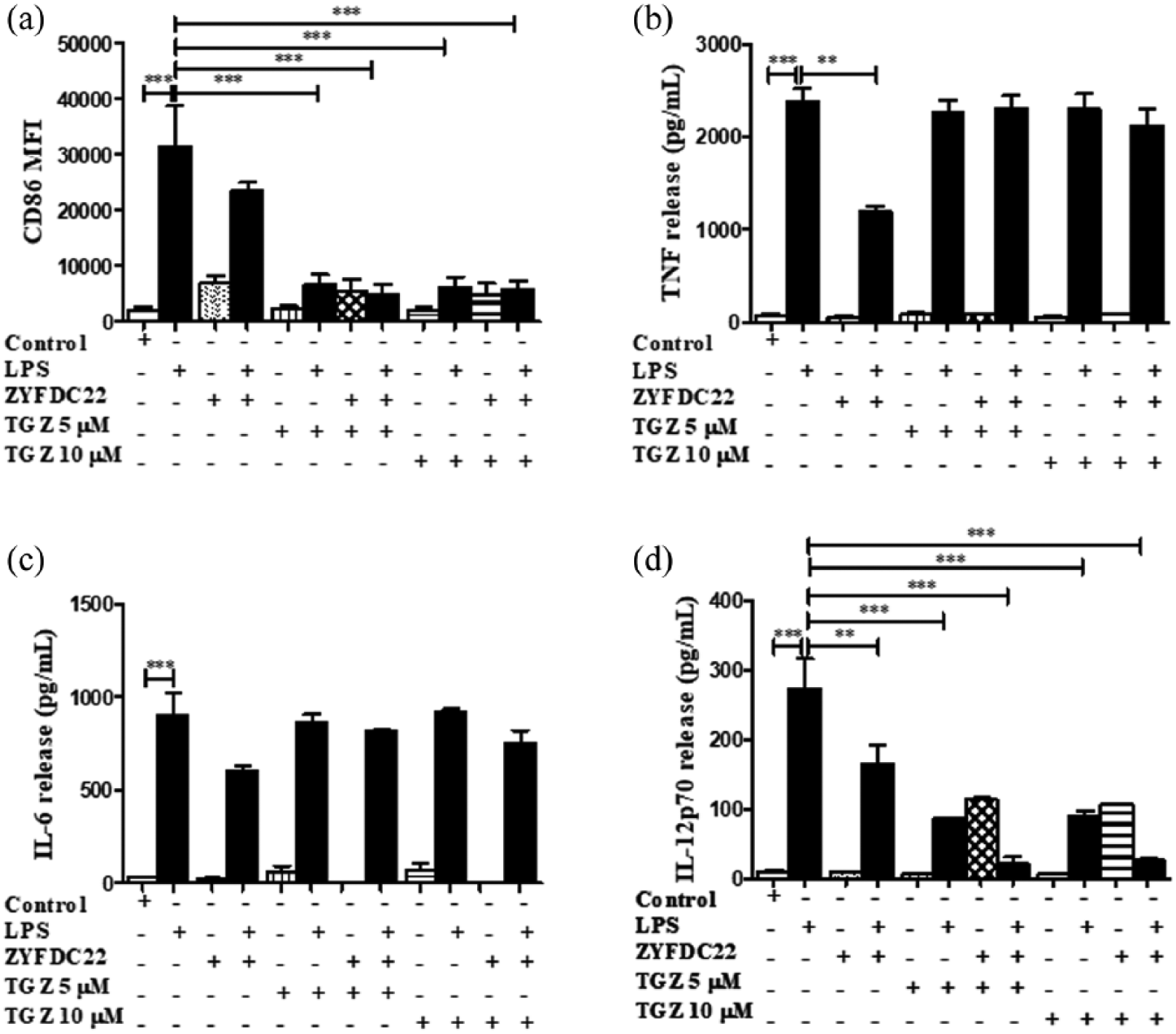
Bicyclic eremophilane-type petasite sesquiterpene ZYFDC22 potentiate the effects of PPARγ agonists on BmDC maturation and activation. BmDC were pretreated with the eremophilane-type sesquiterpene ZYFDC22 plus PPARγ synthetic agonist TGZ (5 or 10 µM) for 3 h, followed by LPS overnight stimulation, and DC were collected, fixed, and analyzed by flow cytometry. Differences in the (a) CD86 surface expression are represented as differences in MFI ± SEM between LPS activated–BmDC and ZYFDC22 + TGZ + LPS (n = 5; ***P < 0.001). Cytokine release by BmDC preincubated with ZYFDC22 and PPARγ agonist TGZ for 3 h ± LPS overnight stimulation, and cell-free supernatants were collected and tested for (b) TNF, (c) IL-6, or (d) IL-12p70 release by ELISA. Results are from cytokines released from LPS-activated BmDC, compared to cells treated with ZYFDC22 + TGZ ± LPS. Data are expressed as means ± SEM (n = 5; *P < 0.05, **P < 0.001, and ***P < 0.001).

Petasite sesquiterpene ZYFDC21 in combination with GW1929 inhibited CD86 and cytokine secretion on BmDC. BmDC were pretreated with the eremophilane-type sesquiterpene ZYFDC21 plus PPARγ agonist GW1929 (40 μM) for 3 h, followed by LPS overnight stimulation, and DC were collected, fixed, and analyzed by flow cytometry. (a) Differences in the CD86 surface expression are represented as differences in MFI ± SEM between LPS-activated BmDC and ZYFDC21 + TGZ + LPS (n = 5; ***P < 0.001). Cytokine release by BmDC preincubated with ZYFDC21 plus GW1929 ± LPS overnight stimulation, and cell-free supernatants were collected and tested for (b) TNF, (c) IL-6, or (d) IL-12p70 release by ELISA. Results are from cytokines released from LPS-activated BmDC and compared to cells treated with ZYFDC21 plus GW1929 ± LPS. Data are expressed as mean ± SEM (n = 5; *P < 0.05, **P < 0.001, and ***P < 0.001).
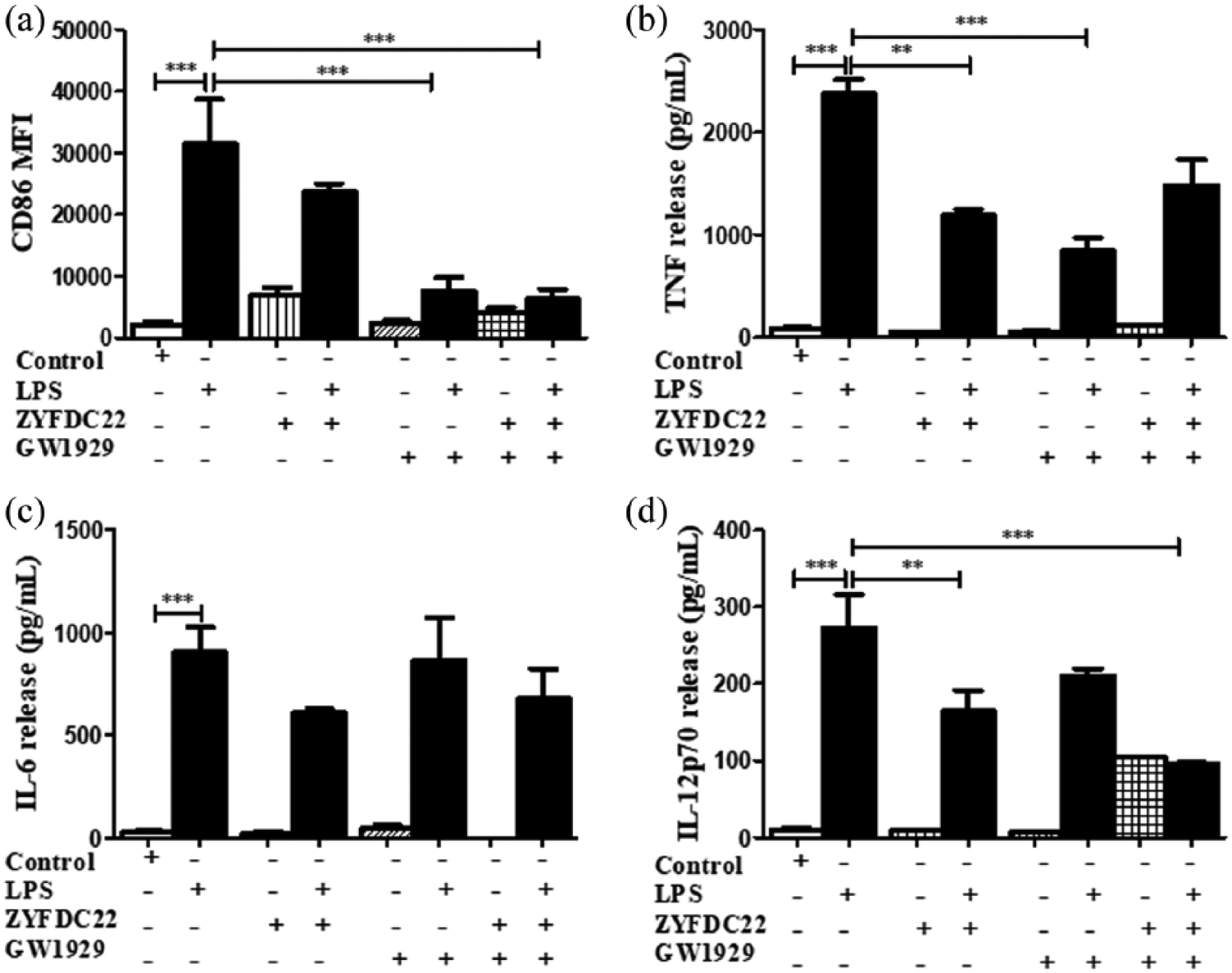
Petasite sesquiterpene ZYFDC22 in combination with GW1929 inhibited CD86 and cytokine secretion on BmDC. BmDC were pretreated with the eremophilane-type sesquiterpene ZYFDC22, plus GW1929 (40 µM) for 3 h, followed by LPS overnight stimulation, DC were collected, fixed and analyzed by flow cytometry. Differences in the CD86 surface expression (a), are represented as differences in MFI ± SEM between LPS activated-BmDC and ZYFDC22 + GW1929 + LPS (n = 5; ***P < 0.001). Cytokine release by BmDC preincubated with ZYFDC22 plus GW1929 ± LPS overnight stimulation, cell–free supernatants were collected and tested for TNF (b), IL-6 (c), or IL-12p70 (d) release by ELISA. Results are from cytokines released from LPS-activated BmDC, compared to cells treated with ZYFDC22 plus GW1929 ± LPS. Data are expressed as means ± SEM (n = 5; **P < 0.001 and ***P < 0.001).
Second, we evaluated the activation of the immune response by the presence of the pro-inflammatory mediators TNF, IL-6, and IL-12p70 released in the cell-free supernatants of BmDC treated for 3 h with the synthetic PPARγ agonists (TGZ or GW1929), in combination with the petasite sesquiterpene (ZYFDC21 or ZYFDC22) and followed by LPS overnight stimulation. BmDC treated with the sesquiterpene ZYFDC21 and synthetic PPARγ agonist GW1929 and stimulated with LPS showed a significant 43% ± 9% inhibition on TNF release (Figure 4(b)). IL-6 released values showed that this cytokine was not significantly affected by any of the tested treatments.
In these studies, we found that IL-12p70, the bioactive isoform of IL-12, seems to be involved in the PPARγ/petasine sesquiterpene pathway. BmDC exposed for 3 h to the sesquiterpenes ZYFDC21 or ZYFDC22 in combination with synthetic PPARγ agonists, followed by overnight stimulation with LPS showed ≥ 90% inhibition for either sesquiterpene in combination with TGZ (5 and 10 μM). The same was true when BmDC were treated with ZYFDC21 or ZYFDC22 in combination with GW1929, followed by stimulation with LPS, where we observed a substantial inhibition of IL-12p70 release (75%±10% for ZYFDC21 and 64±1% for ZFDC22) (Figure 4(d)).
Petasite sesquiterpenes potentiate the effects of PGD2 metabolites on BmDC maturation and activation
BmDC were exposed to the natural PPARγ ligand (15d-PGJ2, 0.5 and 5 µM) for 3 h in combination with eremophilane sesquiterpenes ZYFDC21 and ZYFDC22, followed by LPS overnight stimulation. We observed a robust inhibition in the expression of the co-stimulatory molecule CD86. Cells incubated in the presence of 15d-PGJ2 and sesquiterpenes ZYFDC21 or ZYFDC22 plus LPS showed downregulation in more than 78% ± 6% and 82% ± 6% in the expression of CD86 with 0.5 and 5 µM of 15d-PGJ2 (Figures 6(a) and 7(a) respectively). Also, when BmDC were exposed to petasite sesquiterpenes ZYFDC21 or ZYFDC22 plus LPS, there was a 50% ± 2% inhibition in the TNF release (Figures 6(b) and 7(b)). In addition, the combination of the natural PPARγ ligand pre-treatment plus petasite sesquiterpenes and LPS overnight stimulation promoted a modest IL-6 inhibition of 9% and 18% for ZYFDC21 (Figure 6(c)), but a solid 39, 60% IL-6 inhibition following a inhibitory trend with the PPARγ ligand (Figure 7(c)), not reaching statistical significance. The natural PPARγ agonist, 15d-PJG2, seems to have an additive inhibitory effect on the release of IL-12, which was significantly inhibited with the combination of sesquiterpene ZYFDC21 and 0.5 µM (54% ± 10%) or 5 µM (56% ± 10%) after LPS stimulation (Figure 6(d)). However, when we tested the sesquiterpene ZYFDC22 in combination of 15d-PJG2, we found a 30% ± 10% IL-12p70 inhibition at 0.5 µM and a 50% ± 10% IL-12p70 inhibition at 5 µM 15d-PJG2, respectively (Figure 7(d)). Both sesquiterpenes were able to significantly inhibit around 35% of IL-12 release after LPS stimulation.

Petasite sesquiterpene ZYFDC21 potentiates the effects of PGD2 metabolites on BmDC maturation and activation. BmDC were pretreated with the eremophilane-type petasite sesquiterpene ZYFDC21 ± the natural PPARγ ligand 15d-PGJ2 0.5 or 5 µM for 3 h, followed by LPS overnight stimulation, and BmDC cells were collected, fixed, and analyzed by flow cytometry. (a) Differences in the CD86 surface expression are represented as differences in MFI between LPS-activated BmDC and the combination of ZYFDC21 + 15d-PGJ2 + LPS (n = 5; **P < 0.01 and ***P < 0.001). BmDC were pretreated with ZYFDC21 ± 15d-PGJ2 (0.5 or 5 µM) for 3 h, followed by LPS overnight stimulation, and cell-free supernatants were collected and cytokine release was analyzed by ELISA. Differences in (b) TNF, (c) IL-6, or (d) IL-12p70 released are represented as differences between LPS activated-BmDC and ZYFDC21 plus 15d-PGJ2 ± LPS (n = 5; *P < 0.05, **P < 0.01, and ***P < 0.001).

Petasite sesquiterpene ZYFDC22 potentiate the effects of PGD2 metabolites on BmDC maturation and activation. BmDC were pretreated with the eremophilane-type petasite sesquiterpene ZYFDC22 ± the natural PPARγ ligand 15d-PGJ2 0.5 or 5 µM for 3 h, followed by LPS overnight stimulation, and BmDC cells were collected, fixed, and analyzed by flow cytometry. (a) Differences in the CD86 surface expression are represented as differences in MFI between LPS-activated BmDC and the combination of ZYFDC22 + 15d-PGJ2 + LPS (n = 5; **P < 0.01 and ***P < 0.001). BmDC were pretreated with ZYFDC22 ± 15d-PGJ2 (0.5 or 5 µM) for 3 h, followed by LPS overnight stimulation, and cell-free supernatants were collected and cytokine release was analyzed by ELISA. Differences in (b) TNF, (c) IL-6, or (d) IL-12p70 released are represented as differences between LPS-activated BmDC compared to ZYFDC22 plus 15d-PGJ2 ± LPS (n = 5; *P < 0.05, **P < 0.01, and ***P < 0.001).
Discussion
DCs are the most potent antigen-presenting cells (APCs) and are involved in initiating the adaptive immune responses. The expression of surface adhesion (CD40) and co-stimulatory (CD80 and CD86) and major histocompatibility complex (MHC) class-II molecules promote the contact between DCs and T-cells, while co-stimulatory molecules signal T-cells to proliferate and differentiate. 27 In the mouse, CD86 is the main activation marker of bone marrow–derived DC, being strongly upregulated after maturation, while CD80 expression is less relevant for murine DC.27,28 In this context, our studies demonstrated that LPS stimulation upregulates CD80 and CD86 expression on BmDC. We also confirmed that the use of synthetic (TGZ and GW1929), as well as natural (15d-PGJ2) PPARγγ ligands decreased the expression of CD86 after LPS stimulation. Furthermore, we observed a significant reduction on TNF cytokine release with the GW1929, while IL-12p70 production was attenuated by TGZ and PGJ2. These results were similar to previous reports.14–16,29–31
Advances in the investigation of plant–derived chemicals used in alternative medicine for the treatment of several chronic diseases have shown that Petasite species from petasite sesquiterpenes possess anti-inflammatory properties.32–34 Due to their anti-inflammatory effects mediated via leukotriene synthesis inhibition, sesquiterpenes have been used for the treatment of inflammatory diseases such as arthritis, migraine, as well as asthma and allergy.35–37
The anti-inflammatory effect of Petasite sesquiterpenes is based on their ability to block Ca2+ channels, decreasing intracellular Ca2+ concentration, inhibiting leukotriene B4 and cysteinyl leukotrienes synthesis in eosinophils and neutrophils.21,33,37–43 The active components are sesquiterpene esters of the eremophilane type, and their bioactivity is attributed to petasine and isopetasine.44,45 Studies by Shimoda et al. 46 showed that the effective constituent in the extract of Petasites japonicus was petasine, which had inhibitory effects on leukotriene synthesis 39 and bronchoconstriction. 47 Another eremophilane-type sesquiterpene ketone, namely, Fukinones (1 and 3), 48 exerted suppressive mechanisms in a type I hypersensitivity model in rats and IgE-sensitized RBL-2H3 cells through inhibition of smooth muscle constriction and inhibition of degranulation, leukotriene release, and TNF production by mast cells.33,46 In this context, Lee et al. 33 also reported anti-allergic and anti-inflammatory effects of several compounds obtained from plants of the petasites genus in an ovalbumin-induced asthma model; the molecule Bakkenolide B isolated from P. japonicus inhibited the migration of eosinophils, macrophages, and lymphocytes to the lungs. Previous studies from our lab showed that different extracts of petasites could inhibit type I and type IV hypersensitivity in mouse models of homogeneous and heterogeneous passive cutaneous anaphylaxis. 49
Elegant studies by Lin, 20 evaluated the agonistic activity of the sesquiterpene lactones tirotundin and targitining A, isolated from Tithonia diversifolia against PPARs. For this, they used a transient transfection reporter assay with HepG2 cells and found that tirotundin and targitining A transactivate PPARγ-dependent promoters, including PPRE (PPARγ response element), SHP, and ABCA1, and that both sesquiterpene lactones transactivate PPARγ by binding directly to the PPARγ ligand-binding domain (LBD). In this context, Zhang et al. 19 showed that five isolated components of C. odorata, another plant used in traditional medicine for their anti-inflammatory activities, had a transactivation effect on PPARγ. More recent studies by Wu et al. 50 demonstrated by luciferase reporter assay in HEK293 cells that the bicyclic sesquiterpene trans-caryophyllene aroma compound of plant foods and teas activates PPARα through direct interaction with the LBD of PPARα. However, trans-caryophyllene showed no binding affinity for or transactivation of PPARγ.
However, Adachi et al. 45 demonstrated that petasin derived from P. japonicus activates adenosine monophospahte–activated protein kinase (AMPK) in the liver, skeletal muscle, and adipose tissue of mice, via phosphorylation of AMPK. AMPK activation enhanced the transcription of the proliferator-activated receptor-γ coactivator-1α (PGC-1α), which regulates the genes involved in energy metabolism including mitochondrial biogenesis.
Our studies showed that the eremophilane-type sesquiterpenes ZYFDC21 and ZYFDC22 increased CD80 and CD86 surface expression in non-stimulated BmDC. In contrast, when the cells were pretreated with sesquiterpene ZYFDC21 (Fukinone) followed by LPS, we observed a decrease in CD86 surface expression. This inhibition was amplified up to 80% by the presence of the PPARγ agonists TGZ, GW1929, and 15d-PGJ2. The inhibitory effect was also observed when we used ZYFDC22 (10βH-8α,12-Epidioxyeremophil-7(11)-en-8β-ol) in combination with TGZ, GW1929, or 15d-PGJ2, followed by LPS stimulation, where 85% of CD86 surface expression was significantly inhibited by the PPARγ agonists. The absence of co-stimulatory molecules, such as CD86, influences DC function, altering their maturation and varying the expression of the necessary signals required for the activation and differentiation of naïve T-cells into type 1 (IL-12 and interferon gamma (IFNγ)) or type 2 (IL-4, IL-5, and IL-10) cytokine-producing cells. In this context, our studies showed that both sesquiterpenes ZYFCD21 and ZYFDC22 inhibited the secretion of the soluble factors TNF and IL-12p70 after LPS stimulation. These results are comparable to those obtained by Uchi et al., 51 who demonstrated that the sesquiterpene lactone parthenolide inhibited DC maturation and cytokine secretion induced by LPS.
The level of IL-12 secreted by DC induced by microbial pathogens, such as LPS, during the immunological synapse is a key factor in the outcome of immune responses. IL-12 is a critical Th1-skewing cytokine that elicits IFNγ production by T-cells and by natural killer (NK) cells, 52 favoring a Th2/Th3 response and inhibiting T cell recruitment. 53 PPARγ is an important modulator on B and T lymphocytes as well as DC14,54,55 and PPARγ ligands include a class of antidiabetic drugs, thiazolidinediones (TZD); as well as naturally produced PGD2 and its metabolite 15-dideoxy-Δ PGJ2 (15d-PGJ2), which associate irreversibly to the receptor through covalent binding, mediating their effects by activation of PPARγ-dependent and independent pathways.17,56 Prostaglandins’ production results in activation of PPARγ-mediated transcription, leading to the inhibition of differentiation, migration, and cytokine secretion by antigen-presenting cells, such as DC or macrophages, hence affecting the priming and effector functions of T lymphocytes. 17
Our studies showed for the first time that DC exposed to the PPARγ ligands TGZ, GW1929, and 15d-PGJ2 in the presence of these novel isolated bicyclic eremophilane-type petasite sesquiterpenes ZYFDC21 and ZYFDC22 followed by LPS stimulation exhibited a significant reduction (up to 95%) in the production of the bioactive isoform of IL-12 (IL-12p70). In this regard, it has been documented that 15d-PGJ2 abrogates IL-12 production by directly inhibiting the function of IκB kinase (IKK), therefore preventing the translocation of nuclear factor-κB (NF-κB) to the nucleus.30,57–59 Our results showed that sesquiterpenes reduced LPS-induced DC maturation and inhibited TNF and IL-6 release, as well as the production of the bioactive isoform of IL-12p70, presumably through the direct activation of PPARγ. Since it is well known that the transcription factor NF-κB plays a key role in the activation of PPARγ in the inflammatory response, it would be of interest to determine whether sesquiterpenes bind directly to the PPARγ receptors, thereby inhibiting IKK, and to analyze the downstream signaling cascades that would prevent the translocation of NF-κB to the nucleus, interfering with the inflammatory response. In summary, our results suggest that the novel Fukinone and 10βH-8α,12-Epidioxyeremophil-7(11)-en-8β-ol sesquiterpenes derived from P. tatewakianus inhibit the maturation of DC, as well as the production of TNF, IL-6, and IL-12p70 after LPS stimulation. These events seem to be mediated and potentiated by the activation of PPARγ. Petasite sesquiterpenes are compounds with significant potential value for the treatment of inflammatory disorders.
Supplemental Material
Supplementary_figure_1 – Supplemental material for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells
Supplemental material, Supplementary_figure_1 for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells by Narcy Arizmendi, Chenjie Hou, Fujiang Guo, Yiming Li and Marianna Kulka in International Journal of Immunopathology and Pharmacology
Supplemental Material
Supplementary_figure_2 – Supplemental material for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells
Supplemental material, Supplementary_figure_2 for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells by Narcy Arizmendi, Chenjie Hou, Fujiang Guo, Yiming Li and Marianna Kulka in International Journal of Immunopathology and Pharmacology
Supplemental Material
Supplementary_figure_3 – Supplemental material for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells
Supplemental material, Supplementary_figure_3 for Bicyclic eremophilane-type petasite sesquiterpenes potentiate peroxisome proliferator–activated receptor γ activator–mediated inhibition of dendritic cells by Narcy Arizmendi, Chenjie Hou, Fujiang Guo, Yiming Li and Marianna Kulka in International Journal of Immunopathology and Pharmacology
Footnotes
Acknowledgements
The authors would like to thank Dr Ramses Ilarraza for his comments on the manuscript revision.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study received financial support from the Canadian Institutes of Health Research (CIHR), the National Research Council of Canada (Intramural Funding), and the Natural Science Foundation of China (Canada-China Joint Health Research Initiatives Grant 81261120567).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.