
Abstract
Over the past century, the increasing human impact on the planet’s natural systems has been directly linked to the growing concern over biodiversity loss, and ultimately, human health. Reproduction, a critical factor underpinning species sustainability, is a complex and energetically demanding process that follows similar mechanisms across species. Further, it is one of the first biological functions to be altered in the event of ecosystem stress. The One Health paradigm, connecting humans, animals and plants to their environments, is a valuable framework for assessing the influence of anthropogenic changes on reproductive success across species. This review assesses the impacts of environmental pollution and climate change on the reproductive health of humans, animals and plants to better understand the broader scope of reproductive decline across the biodiversity spectrum, a fundamental step toward implementing One Reproductive Health objectives.
Introduction
The concept of One Health, initially One Medicine, has occupied the thoughts and practices of human and veterinary medical professionals throughout the past 100 years, and much longer (Zinsstag et al., 2011). Since the new millenium, however, a greater understanding of the dependence of human and animal health on ecosystem health in the emergence or resurgence of infectious and non-infectious diseases has led to the establishment of “One World–One Health” initiatives (Destoumieux-Garzón et al., 2018). Throughout Earth’s history, life has endured periods of both flourishing and crisis, marked by five major mass extinctions, each leading to significant biodiversity loss (Figure 1). Scientists now warn that we are entering the sixth mass extinction, driven not by natural events, but by human activities (Barnosky et al., 2011; Ceballos et al., 2015). The current trajectory of biodiversity loss mirrors the catastrophic declines seen in past epochs, raising urgent questions about the sustainability of life on Earth in the Anthropocene. Since 1970, a span of only 50 years, the human population has more than doubled (Butler, 2016), while vertebrate population sizes have more than halved (Almond et al., 2020). These changes in population numbers, along with the concomitant shift in human behavior toward a “throw-away society” that encompasses excessive consumption of short-lived products (Vince, 2013), have had numerous downstream consequences that include not only the loss of biodiversity, but also habitat loss, environmental degradation and contamination, climate change, and disease emergence (Deem et al., 2019). The continuous and increasing infringement on the planet’s natural systems in this new epoch has necessitated novel and innovative approaches to assessing, resolving and maintaining the health of humans, animals, and plants, and the complex environments that sustain them.
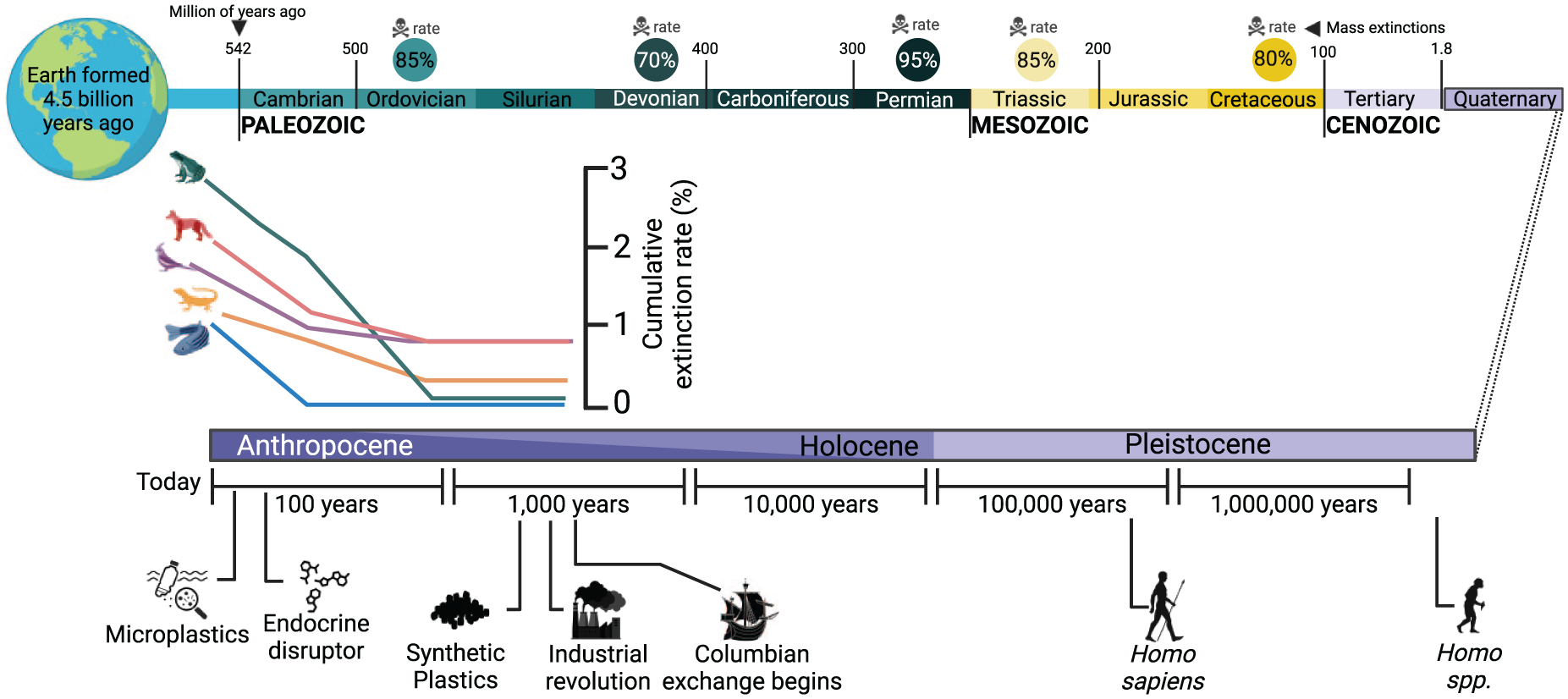
Timeline of Earth’s major mass extinctions and biodiversity trends in the Anthropocene. This illustration presents the timing of Earth’s five major mass extinctions (skull symbols), along with the percentage of species lost during each event: the Ordovician-Silurian (85%), Devonian (70%), Permian (95%), Triassic (85%), and Cretaceous (80%). The progression of geological epochs, from the Cambrian through the Anthropocene, with key human evolutionary and industrial milestones, are indicated. The recent sharp decline in vertebrate populations is shown (adapted from (Le Gac et al., 2021)), and corresponds to the Anthropocene epoch, where human-induced environmental impacts, such as industrialization, deforestation, and climate change, have escalated, potentially leading to the sixth mass extinction.
A key factor underpinning the long-term sustainability of species and ecosystems globally is reproductive success. As history has shown us, both too much or too little reproductive output can undermine the stability of a population, particularly when natural mechanisms required to keep the balance in check are lacking or disrupted. This review touches on the current state of reproductive health, specifically fertility, in humans, animals, and plants (for simplicity, sexually reproducing only) to better understand how anthropogenic pressures have altered natural processes with potentially drastic, if not irreversible, consequences. Groups of living organisms can act as sentinels for each other in this rapidly changing landscape; alterations in human reproductive functions may be indicative of similar occurrences in wild fauna and flora, and vice versa. A better understanding of the broader scope of reproductive decline across the biodiversity spectrum will provide us with the necessary knowledge to implement One Reproductive Health objectives.
Brief overview of reproduction
Reproduction in living organisms takes many forms, each tailored to specific ecological and evolutionary needs. Asexual reproduction, such as binary fission in bacteria and budding in yeast, allows organisms to reproduce rapidly without a mate (Bell, 2019). Fragmentation, as seen in starfish, and vegetative propagation in plants, also exemplify asexual methods where new individuals arise from parts of the parent organism (Smith, 1982). Parthenogenesis, found in some reptiles, fishes, and insects, enables females to produce offspring from unfertilized eggs (Avise, 2008). Polyploidy, common in plants, involves multiple sets of chromosomes and contributes to reproductive isolation and speciation (Comai, 2005). Additionally, hermaphroditism, present in certain fish and invertebrates, allows organisms to possess both male and female reproductive organs, increasing reproductive opportunities (Ghiselin, 1974). Finally, apomixis in plants produces seeds without fertilization, leading to genetically identical offspring, a valuable trait in agriculture (Bicknell, 2004).
While these reproductive strategies illustrate the diversity of life, our focus here will be on sexual reproduction—a process characterized by the fusion of male and female gametes to produce genetically unique offspring (Alberts et al., 2002). Reproduction can be year-round in certain species, and strictly seasonal in others where gamete production, and subsequently, fertilization, is influenced by environmental factors, such as temperature, light, rainfall and nutritional resources (Mastromonaco and Gonzalez-Grajales, 2020). Sexual reproduction is not only responsible for producing more individuals of a species, but also promotes genetic variation due to the chromosome recombination events that occur during meiosis to support adaptation and evolution of a species in response to a changing environment.
Reproductive success in humans, animals, and plants is generally defined as an organism’s capacity to produce offspring that will subsequently have the potential to give rise to successive generations (Stearns, 1998). It is influenced by numerous intrinsic and extrinsic factors, and generally related to good physical health and genetic make-up. Evaluation of reproductive status, a prediction of potential reproductive output, typically involves the assessment of gamete quantity and quality, among other metrics, which can include morphological (structural/anatomical) and physiological (biochemical/hormonal) characteristics (Garolla et al., 2020). While quantifying reproductive success in humans and domestic or companion/farm animals is relatively well established, the vast and diverse kingdom of animals creates challenges in measuring this outcome. Nonetheless, considering nervous and endocrine systems are largely conserved across animal taxa and play major roles in reproduction, comparative analysis between humans and animals is mutually beneficial to comprehend the effects of environmental disturbances on reproductive success. Correspondingly, the vast and diverse kingdom of plants creates even greater challenges in measuring reproductive success since they do not have nervous or endocrine systems like those of humans and animals (Box 1).
Homeostasis, hypothalamic-pituitary-gonadal (HPG) axis, and sexual differentiation.

Alterations in reproductive health lead to a variety of outcomes, including reduced gamete production, increased structural and functional abnormalities in gametes, and altered embryo/fetal/neonatal development, thereby reducing the fecundity of the individual and, if widespread, of the species. These changes can be attributed to a host of causative factors that can be divided into two main categories: (i) heritable: e.g. gene mutations, chromosomal errors; and (ii) acquired: e.g. infectious disease, trauma (Imran et al., 2025). Over the past 50 years, there has been significant evidence of increasing rates of heritable and acquired reproductive dysfunction in humans, animals and plants that can be linked to anthropogenic-related challenges, as identified in the sections below.
A century of human-driven change
Historically, there is no disputing the fact that humans have left an indelible mark on the planet. Since the start of the Industrial Revolution in the 18th century, urbanization and industrialization of agricultural societies along with the resulting technological and medical advancements have led to the rapid expansion in human population and, concomitantly, to changes in land-use practices and consumption of natural resources (Ellis et al., 2010; Steffen et al., 2007). The Anthropocene is an era that can be defined in a manner of ways, including the point when human activities: (i) had an influence on the planet’s sediments, (ii) dominated consumption of global net primary production, and (iii) affected global climate (Doughty, 2013). Two of the most widespread, and potentially devastating, consequences with known links to the reproductive success of all living things include environmental pollution and climate change. Environmental toxicants, in the form of natural by-products as well as synthetic molecules and micro-aggregates, continue to be ubiquitous and can be found in air, water, soil, and even the tissues and cells of living organisms. Correspondingly, alterations in temperature, humidity and weather patterns are increasingly evident not only in extreme landscapes, such as the polar regions, but across the globe.
Despite more than three decades of international collaborations to halt the damage created by the activities of a growing human population, the environment remains in a fragile state. Failure to meet the Aichi Biodiversity Targets by 2020 (Nature, 2020), a framework established by the Convention on Biological Diversity (CBD) in 2010, had left the planet in an even more critical state as nations continued to work toward repairing the environment and reversing the number of species at risk. In December 2022, a new framework, the Kunming-Montreal Global Biodiversity Framework, was adopted during the CBD’s Conference of the Parties (COP15) that lays out the necessary steps toward the global vision of a world “living in harmony with nature by 2050” (Convention on Biological Diversity, 2022). Understanding the long-lasting effects of an altered planet on reproductive systems of animal and plant species is essential for developing the necessary tools and strategies to support global policy changes that will ensure the reproductive success of all living things into the future.
For the purposes of this review, the discussion has been focused on a few key concerns. The topic of environmental health and its decline is so extensive, due to the many interconnected systems and multi-factorial stressors involved, that in-depth coverage of animal and plant kingdoms would be difficult to convey within the constraints of a review paper. Gallo et al. (2020) published an informative review on the impact of environmental stressors on animal gamete quality. We hope to add to the literature by identifying similarities and building connections between the animal (inclusive of humans) and plant kingdoms.
Anthropogenic impacts on reproductive success
Environmental pollution
The Anthropocene era—as characterized, in part, by extensive environmental pollution by humans—is widespread across the planet. Synthetic chemicals, including endocrine-disrupting chemicals (EDCs), have been found to mimic natural hormones, causing disruptions in human, animal, and plant health and ecosystem sustainability. The discovery of microplastics (MPs) in 2004 further underscored the pervasive nature of pollution, with these tiny plastic particles contaminating land, water, and air. The emergence of EDCs and MPs has revealed their detrimental effects on reproductive health, development, and growth, contributing to biodiversity loss and ecosystem degradation. These significant consequences emphasize the urgent need for comprehensive environmental management, stricter regulations, and global conservation efforts to mitigate the impact of human activities on the environment.
Endocrine disrupting chemicals
In Rachel Carson’s book “Silent Spring” published in 1962, she documented her awareness and concerns for the use of pesticides and their unintended consequences on wildlife and environmental health, and coined the term “biocides” (Carlson, 1962). The massive impact of her observations alerted the public and corresponded with the creation of the United States Environmental Protection Agency (USEPA) in 1970. Almost 26 years later, Theo Colborn authored the book “Our Stolen Future” published in 1996 based on the outcome of a historic meeting in 1991 regarding the health effects of environmental chemicals with hormone-like activities on humans and wildlife. This reminded global authorities and public of the concept of One World-One Health, and introduced the term “endocrine disrupting chemicals” (Colborn et al., 1996).
Beginning around 1992, international organizations, such as the World Health Organization (WHO), International Program on Chemical Safety (IPCS), Organization for Economic Cooperation and Development (OECD), and nations, including European Union (EU) countries, Japan, and USA, started to discuss national and international mandates to address the concerns of EDCs. In 1996, both the OECD and the US Congress initiated programs and established federal acts to better assess EDCs and protect the public’s food and water (Stiefel and Stintzing, 2023). In general, OECD and EPA adopted the definitions of EDC initially proposed by the WHO and IPCS (Damstra et al., 2022): “An endocrine disruptor is an exogenous substance or mixture that alters function(s) of the endocrine system and consequently causes adverse health effects in an intact organism, or its progeny, or (sub)populations,” and “A potential endocrine disruptor is an exogenous substance or mixture that possesses properties that might be expected to lead to endocrine disruption in an intact organism, or its progeny, or (sub)populations.” The OECD and EPA developed frameworks for screening programs to identify EDCs with potential (anti)estrogen, (anti)androgen, and (anti)thyroid hormone activities, determine adverse effects and dose-response, and assess and manage risk, inclusive of pesticides, commercial chemicals, and environmental contaminants (U.S. Environmental Protection Agency, 2015). Since 2004, the Endocrine Disruptor Screening Program Tier 1 and Tier 2 testing have been developed, validated, and approved by EPA. Comparable guidelines have also since been approved by OECD (2012).
EDC sources and exposures
Potential EDCs in the environment can originate naturally from sources involving phytoestrogenic plants (e.g. soy, legumes), and anthropogenically from agricultural use of various classifications of synthetic pesticides (e.g. atrazine, alachlor), industrial by-products (e.g. dioxin-like compounds; polychlorinated biphenyls, PCBs; polychlorinated dibenzofurans, PCDFs), manufacture and combustion of various materials (e.g. plastics, pharmaceuticals, fossil fuels), as well as from other sources (e.g. heavy metals; Amir et al., 2021; Kassotis et al., 2020; Laws et al., 2021; Marlatt et al., 2022).
Historically, the biochemical and lipophilic nature of some EDCs has resulted in their longevity in the environment as persistent organic pollutants (POPs), a term which emerged from the 2001 Stockholm Convention under the United Nations Environment Program (International Institute for Sustainable Development, 2001). An additional outcome from this meeting was a preliminary list of 12 chemicals classified as POPs, termed the “dirty dozen,” that ultimately became international law (Haffner and Schecter, 2014). These initial chemicals consisted of organochlorines for agricultural use as pesticides (e.g. dichlorodiphenyltrichloroethane, DDT) and industrial waste (e.g. polychlorinated biphenyls, PCBs). As of 2022, additional countries have joined the Convention and more POPs have been listed (Chasek, 2023). Notably, many POPs have since been banned from production, restricted in their use, or put under review by domestic or international laws or treaties due to their adverse effects on human and animal health and wellbeing, and overall global presence in the environment. While many POPs are EDCs, many EDCs are not POPs since they are not necessarily persistent in the environment based on their biochemical nature but, nonetheless, persist due to their continuous release into the environment. Regardless of how they are defined, POPs or EDCs typically exist in the environment at relatively low concentrations but can remain active for various lengths of time from weeks, months, or even years as the original compounds or metabolites.
While exposure to EDCs can occur by air, water, soil, sediment, and food, and enter organisms via inhalation, ingestion, skin, leaves, and roots, the degree of impact on the organism is dependent, in part, on dose and duration of exposure as well as species and life stage (Martin et al., 2022). Regarding the latter, EDC exposure and homeostatic imbalance in a mature organism may be compensated for or less severe, with little to no outward effects or adversity on growth, development, and reproduction. However, the same exposure in an immature organism (embryo, fetus, neonate, or seedling) can result in severe adverse effects that can end the life of the organism or persist within gametes and carry over to subsequent generations to affect growth, development, and reproductive success of an individual or population of a species (Brehm and Flaws, 2019).
EDCs that are highly lipophilic in nature can bioaccumulate in fatty or nonpolar tissues (e.g. glycerol-containing lipids) and biomagnify up the food chain in corresponding tissues eventually magnifying to maximal concentrations in top predator species, such as humans and higher level trophic animal species, eliciting adverse effects in individuals and populations. After absorption and distribution within the organism, EDCs undergo different aspects of metabolism dependent on their biochemical nature. EDCs that are lipophilic readily distribute to organs or parts of the organism that are high in fatty or nonpolar tissues. Once sequestered, they are biologically inactive; however, upon lipolysis or tissue catabolism, EDCs can be liberated and redistributed as biologically active compounds in the vascular or vascular-like systems of organisms, creating conditions of re-exposure, toxicity, and adversity (Martin et al., 2022).
EDC exposure and reproductive success
There is substantial evidence to show that most major classes of EDCs target the androgen or estrogen hormonal pathways as agonists or antagonists—in addition to altering gene expression and epigenetic programing—thus, impacting human, animal, and plant reproductive health acutely, chronically, and transgenerationally (Mandava and Mandava, 2021; Sifakis et al., 2017). Humans and animals at all life stages are susceptible to EDCs, but those in early developmental stages—including the prenatal period—are particularly vulnerable. Fetuses in utero are often at the highest risk due to their reliance on maternal physiological systems and the critical windows of development during gestation (Brehm and Flaws, 2019). Even low-dose EDC exposure during this time can result in long-lasting or transgenerational effects on reproductive health. Postnatally, children and young animals remain highly susceptible due to their rapid growth and development (Di Pietro et al., 2023). They consume more air, water, and food relative to body size than adults, and their immature blood-brain barrier and gastrointestinal tract allow easier passage and absorption of EDCs. Additionally, their skin is more permeable, increasing exposure from air, water, and soil. Their metabolism and excretion systems are also less effective due to the ongoing maturation of major metabolic organs and processes (Di Pietro et al., 2023).
In women, EDCs with estrogenic and androgenic-like activities have been associated with impaired reproductive success due, in part, to infertility, aberrations in duration of the menstrual cycle, ovarian dysfunction, endometriosis, and uterine fibroids (reviewed by (Silva et al., 2023; Vessa et al., 2022)). In men, EDCs have been linked to infertility, testicular dysfunction, cryptorchidism, hypospadias, testicular cancer, and decreased sperm count and quality (reviewed by (Quilaqueo and Villegas, 2022)). Correspondingly, while the animal kingdom is more vast and diverse compared to humans, exposure to many of the same EDCs with estrogenic- and androgenic-like activities in vertebrate and invertebrate species has resulted in similar or analogous types of reproductive effects in female and male organisms (reviewed by (Ghosh et al., 2022; Marlatt et al., 2022)). In frogs, for example, commonly used herbicides and pesticides have been the cause of intersex phenotypes (Hayes et al., 2002) and other aberrations associated with disrupted steroid pathways that are being implicated in on-going amphibian declines (Hayes et al., 2006).
The compilation of EDCs shown to alter reproductive success is not necessarily exhaustive but is representative of a legacy of chemicals (e.g. DDT, DDE, PCBs, dioxin, PBA, phthalates, parabens) and their metabolites. Despite many legacy chemicals having been banned or limited to restricted in use by numerous countries globally, many are still persistent within the environment. Moreover, as manufacturers are faced with discontinuation or limited use of products, there is a consumer demand for producers to design alternative chemicals as replacements. For example, due to emerging evidence and concern that perfluoroalkyl substances (PFAS), known as the “forever chemicals” (Schildroth et al., 2022), are having EDC-like effects on development and reproduction, additional flame retardants, such as polybrominated diphenyl ethers (PBDEs; (Vuong et al., 2020)) and organophosphate flame retardants (OPFRs; (Pantelaki and Voutsa, 2019)), are being used. Alternative plasticizers to phthalates and BPA, such as diisononyl phthalate (DINP), diisodecyl phthalate (DIDP), and Bisphenol S (BPS) and F (BPF), are being used in plastic containers and food packaging. While alternative or replacement chemicals are relatively new to consumer use, there is a global effort to subject these emerging chemicals to continued research and regulatory scrutiny to assess their hazards and risks as potentially more contemporary EDCs that may have adverse effects on human and animal reproductive success.
Understanding the effects of EDCs on plants is essential for tracing contamination pathways that impact both humans and animals relying on plants as a food source. Exposure to pesticides (e.g. 2,4-D, DDT), industrial pollutants (e.g. phthalates, BPA, PCBs), and heavy metals (e.g. fluoride, lead) can harm plant populations, reducing genetic diversity across terrestrial and aquatic species (Saraswat et al., 2024). From germination to seed formation, EDC exposure can disrupt growth, development, and maturation that can alter flower formation timing, flower morphology, or flower fertility (i.e. the ability of the plant to produce viable seeds and reproduce successfully). Further, evidence suggests that EDCs can affect growth and development of organs, such as roots, shoots, leaves, flowers, and fruits, growth rate, pollen germination, plant hormone levels, photosystems, microtubule and cytoskeletal elements, production of reactive oxygen species, and cell division ((Adamakis et al., 2016; Bahmani et al., 2020; Saraswat et al., 2024); Figure 2). EDC exposure can also interfere with hormonal signaling pathways that may involve auxins, cytokinins, gibberellins, abscisic acid, ethylene, and other factors that play crucial roles in regulating plant growth, development, and reproduction ((Bahmani et al., 2020; Saraswat et al., 2024); as shown in Box 1). For example, Speranza et al. (2010) demonstrated that BPA exposure significantly inhibited pollen tube emergence and elongation in kiwifruit (Actinidia deliciosa), along with inducing morphological changes in pollen tubes. These reproductive impairments were linked to altered steroid hormone production, specifically marked increases in 17β-estradiol and testosterone concentrations, as well as elevated expression of stress-response proteins such as BiP and 14-3-3 (Speranza et al., 2011). Additionally, exposure to perfluorooctanoic acid inhibited seed germination and seedling growth in wheat (Triticum aestivum), which also had altered antioxidant enzyme activities (POD and CAT) and increased proline content—a stress response indicator (Zhou et al., 2016).
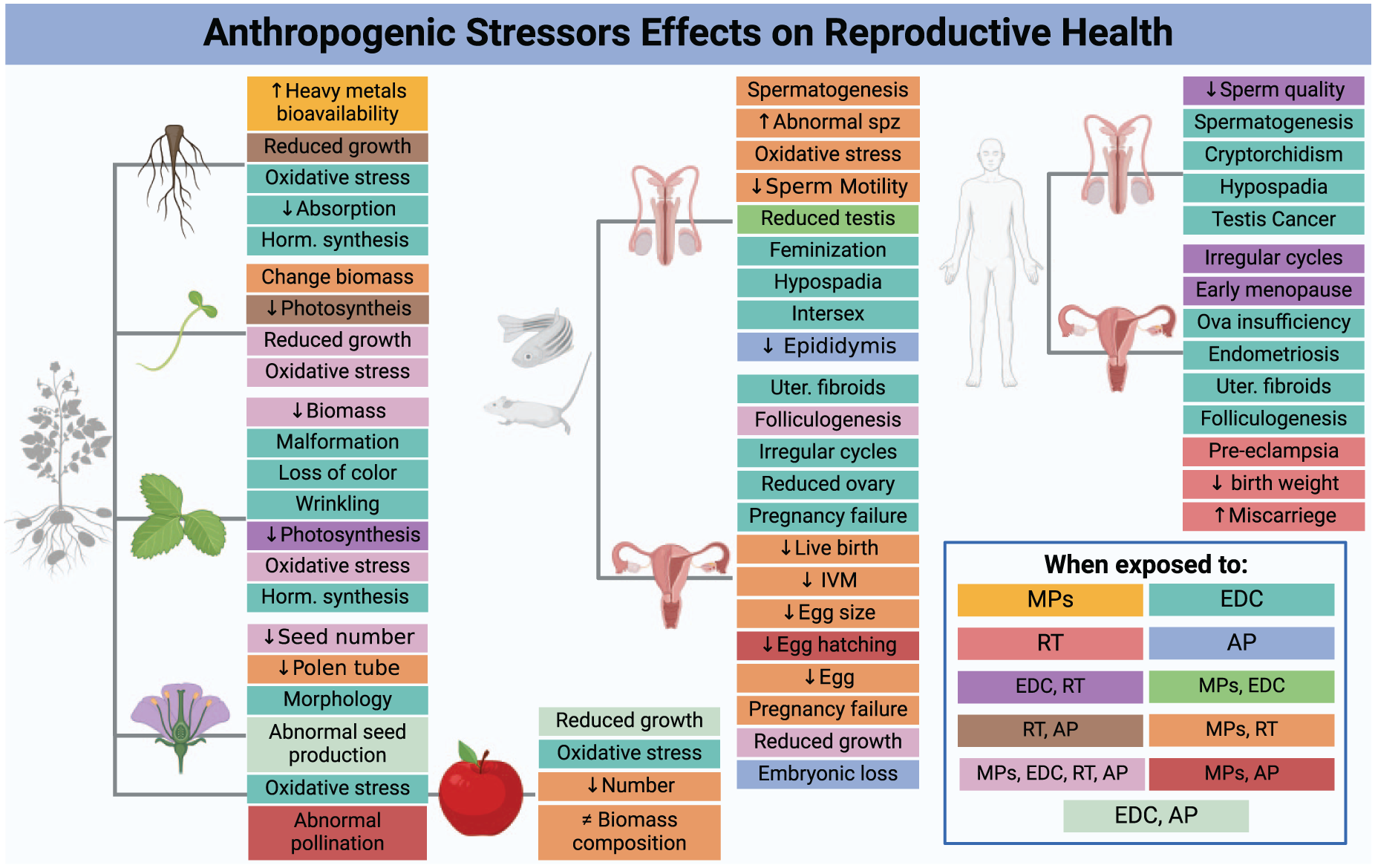
Summary of anthropogenic effects on reproductive success. This figure illustrates the various reproductive outcomes in plants (roots, seedlings, leaves, flowers, and fruits), animals (males and females), and humans (males and females) when exposed to environmental stressors including endocrine disrupting chemicals (EDCs), microplastics (MPs), rising temperatures (RT), and altered precipitation (AP). Each effect is depicted within a colored box, where the color represents the environmental factor causing that effect, as detailed in the legend on the right bottom. Some of the effects of altered precipitation are predominantly secondary (e.g. malnutrition, less access to resources, stress, and higher risks for infections) and are not included.
Microplastics
The history of plastic began in the late 19th century with the invention of Bakelite in 1907, the first fully synthetic plastic, marking the start of the modern plastics era (Nicholson and Leighton, 1942). The widespread production and use of plastics exploded in the mid-20th century, transforming industries and daily life due to their versatility and durability. Decades of careless use and improper disposal have resulted in plastic pollution accumulating almost everywhere on Earth (Rochman and Hoellein, 2020). The ca. 8 billion tons of plastic pollution scattered across the planet have made plastics synonymous with the seemingly irreversible mark of humankind on our planet. By the 1970s, scientists began noticing small plastic particles in the ocean, which were later termed microplastics (MPs). Microplastics have been defined as plastic particles < 5 mm in size that are insoluble in water and are manufactured as such for commercial purposes or derived from the degradation of larger plastic products that have been unintentionally introduced into the environment (Frias and Nash, 2019).
Countries worldwide are implementing stricter plastic policies to combat pollution. Canada banned single-use plastics in 2021, while India’s 2024 Plastic Waste Management Rules target microplastics and require biodegradable alternatives (Drishtti, 2024). California aims for a 65% recycling rate for single-use plastics by 2032 under its new circular economy regulations (CellRecycle, 2024). The EU’s Single-Use Plastics Directive mandates recycled content in PET bottles and tethered caps by 2024 and 2025, respectively (European Commission. Directorate General for Environment, 2021). In 2024, 175 nations gathered in Ottawa to negotiate a legally binding treaty on plastic pollution, expected by year-end, underscoring the global shift toward reducing plastic waste for environmental and public health.
Microplastic sources and exposures
Microplastics are the most recent global concern requiring a One Health lens, prior to the onset of the SARS-CoV-2 pandemic, and significant effort is being made to understand the ramifications of its omnipresence (Prata et al., 2021). On-going studies are investigating the impact of MP exposure resulting from either its physical effects (i.e. size, shape, and concentration) or chemical effects (i.e. toxic properties of the plastic raw materials; (Campanale et al., 2020a)). MPs are now so widespread that they are consistently detected in every biological sample that is surveyed for their presence (Braun et al., 2021; Jenner et al., 2022; Leslie et al., 2022; Ragusa et al., 2021; Schwabl et al., 2019). They have been found in soil across various regions, water bodies from remote lakes to oceans, and even in snow in the Alps. MPs are also present in the air, leading to their inhalation by both animals and humans. Aquatic animals, from small fish to large marine mammals, and terrestrial animals, including livestock and wildlife, show evidence of MP contamination. Additionally, MPs have been detected in a variety of human tissues, indicating their pervasive presence in the environment and raising significant concerns about their impact on health and ecosystems (MacLeod et al., 2021).
Microplastics can be categorized into primary and secondary plastics (Frias and Nash, 2019). Primary MPs are particles intentionally manufactured for commercial use, for example, microbeads in personal care products, and microfibers from clothing and textiles. Secondary MPs, on the other hand, are formed from the fragmentation of larger plastic items, such as bottles and packaging materials, due to environmental factors like UV radiation, mechanical forces, and chemical degradation (Frias and Nash, 2019). Both types contribute significantly to plastic pollution, with primary sources being a direct result of industrial and consumer activities, and secondary sources emerging from the breakdown of improperly disposed plastic waste.
The growing volume of macro- and microplastics making their way into streams, rivers, lakes and oceans has drawn considerable public attention to the plight of aquatic ecosystems. Images of marine life engulfed and trapped by discarded plastics are a distressing sight, and studies on MPs in freshwater and marine ecosystems have been accumulating exponentially. Yet, despite the tens of thousands of peer-reviewed papers on MPs, few have focused on the terrestrial systems, and very little is known about the occurrence and fate of MPs on land. For example, a recent global synthesis on MPs in soils identified only 41 studies on the subject (Mills et al., 2023). This knowledge gap stands in stark contrast to the fact that 80% of the Earth’s species live on land (Grosberg et al., 2012), and terrestrial systems may represent a larger environmental reservoir of MPs than oceans (de Souza Machado et al., 2018; Hurley and Nizzetto, 2018).
Microplastics enter the body through multiple pathways: ingestion, inhalation, and dermal absorption (Sun and Wang, 2023). People ingest MPs through contaminated food and water, and these particles can permeate the gut and migrate to distant tissues, posing various toxicity risks. Inhalation of airborne MPs occurs both indoors and outdoors, with daily inhalation estimates ranging from 26 to 130 particles (Prata, 2018). Additionally, although less significant, dermal absorption of nanoplastics has been demonstrated, indicating various pathways for MPs to enter the human body (Revel et al., 2018). Animals and plants are also susceptible to microplastic contamination. Animals, especially marine organisms, ingest MPs directly from polluted environments or indirectly through their prey, leading to accumulation and potential translocation within their bodies. Terrestrial animals can consume MPs through contaminated soil, water, plants, and prey, as well as by inhalation. Plants can absorb MPs through their roots, with these particles potentially translocating to other parts of the plant, including edible portions (Li et al., 2019).
Microplastic exposure and reproductive success
While humans have introduced numerous pollutants that can impair reproductive systems (Choi et al., 2016; Cole et al., 2011; de Almeida Monteiro Melo Ferraz et al., 2018, 2020; Du et al., 2016; Kalo and Roth, 2017; Kumar et al., 2014; Rattan et al., 2017; Sone et al., 2004), the potential for MPs to be driving the widespread declines in fertility is particularly noteworthy. Over the same timespan that declines in fertility began to be documented, there has been this correlated shift toward a “throw-away society” that is characterized by the excessive consumption of short-lived and single-use plastic products and a concomitant accumulation of MPs pollution. The ubiquitous and long-lived nature of MPs makes them a troubling contaminant, yet, despite their pervasiveness, very little is known about the effects of MPs on reproductive health, especially of species living in terrestrial ecosystems. While evidence is still extremely limited, emerging studies are showing that MPs represent a potentially serious threat to reproductive health (de Souza Machado et al., 2018; Grechi et al., 2023; Hou et al., 2021; Ijaz et al., 2021; Li et al., 2021).
Most reports that have identified a disruption in reproductive processes, primarily noted as reduced offspring production, have associated the ingestion of plastic particles with a reduced capacity for nutritional uptake, and subsequently energetic depletion, that requires resources to be diverted away from reproduction. In the Pacific oyster (Crassostrea gigas), for example, experimental exposure to polystyrene microspheres resulted in significant changes at both the gamete and larval stages after just 2 months (Sussarellu et al., 2016). The authors hypothesized that the decreased reproductive performance was associated with changes in energy allocation that were necessary to compensate for the ingested MP particles. Their theory was supported by the observed alterations in transcriptomic profiles related to glucocorticoid response, insulin pathway and fatty acid metabolism (Sussarellu et al., 2016).
Recent work in mice and rats demonstrated the detrimental effects of polystyrene MPs on sperm production and motility (Ijaz et al., 2021; Jin et al., 2021). Similarly, Hou et al. (2021) identified an increase in sperm abnormalities and testis atrophy, and the programmed death of sperm cells within the testis of mice exposed to MPs (Hou et al., 2021). From a female perspective, a significant decrease in the number of live births and body weight of neonates was observed in mice dosed with polyethylene microplastics compared to the control (Park et al., 2020). Data have been limited to in vivo studies on laboratory rodents that were force-fed plastics (An et al., 2021; Deng et al., 2021; Jin et al., 2021; Li et al., 2021), however, and there are currently no studies that link MPs to changes in fertility in free-ranging mammals. This is not a trivial point. These in vivo lab studies investigated the effects of plastics on fertility by feeding rodents MPs at concentrations that were hundreds of thousands of times greater than they would be exposed to in the wild (Mills et al., 2023). Consequently, although the findings from these studies are certainly worrying, the extent to which they are representative of the conditions that animals are actually experiencing in the real world is questionable at best. This means that it is unclear to what extent MPs can bio-accumulate in free-ranging terrestrial mammals and influence fertility.
While the evidence supporting the detrimental effects of MPs on reproductive function has been primarily from animal studies, the potential for these particles to carry or leach contaminants, such as EDCs, makes them equally suspect in human reproductive health outcomes (Wright and Kelly, 2017). There are currently no experimental or retrospective studies that link MP contamination to changes in human fertility, but the propensity for cellular toxicity and biomimicry remains a significant concern (Wright and Kelly, 2017). Recent studies documented the first cases of a variety of MPs in human placentas from a number of uneventful pregnancies (Ragusa et al., 2021), and in women’s follicular fluid (Grechi et al., 2023). The introduced particles have the potential to interfere with cellular pathways involved in maternal-embryo signaling, pregnancy-related immunity, and more, making this a disconcerting discovery. Moreover, MPs were already detected in human testis and semen (Montano et al., 2023; Zhao et al., 2023). It was shown that samples where no MPs were detected have the best seminal quality in terms of volume, number, motility, and morphology (Montano et al., 2023).
Not only are MPs a threat to human and animal health, but they can also affect plant fertility. Agricultural lands are a potential trap for MPs; industrialized horticulture depends on plastics for mulch films that are used to enhance soil conditions, and therefore, plant growth (Qi et al., 2018). The type of macro- or microplastic used in the soil during a potted wheat growth experiment impacted reproductive outcomes in the form of fruit numbers and biomass (Qi et al., 2018). Similar outcomes were observed with spring onions whereby particle type played an important role in the downstream effects on plant biomass and other related parameters (de Souza Machado et al., 2019).
A recent study by Lozano et al. (2021) demonstrated that both MP shape and polymer type can influence plant and soil parameters more than concentration. Overall, MPs increased shoot and root mass by about 46% and 48%, respectively, but reduced soil aggregation by 25% and microbial activity by 6%. MP films boosted both shoot and root mass by 60%, while fibers had a lesser impact, increasing root mass by just 6%. The presence of certain MPs shapes, especially fibers, enhanced root biomass, which in turn improved water and nutrient uptake due to the fibers’ water-holding capacity. All MP shapes decreased soil aggregation, which is essential for maintaining soil structure, fertility, and overall ecosystem function. The study also underscored that the MP’s shape is pivotal in determining its impact, with different shapes of the same polymer type, like polypropylene, having varied effects on plant growth and soil properties (Lozano et al., 2021). Contrastingly, a study utilizing a combination of hand-pollination treatments and ultrastructural observations demonstrated that MP deposits negatively impacted seed production and reduced the number of pollen tubes reaching the ovaries, while mass per seed and germination rates remained unaffected (Carvallo and Muñoz-Michea, 2023). While MPs confer certain benefits for plant growth, these findings underscore the detrimental effects of synthetic plastics on pollination, a key ecosystem process, and suggest that MP contamination could have significant consequences for angiosperms and crop production.
A study by Pinto-Poblete et al. found that the synergistic effect between heavy metals and MPs can alter soil properties and plant performance. Specifically, when MPs were combined with cadmium (Cd), there was a noticeable effect on soil microbiological activity, growth, and yield parameters of strawberry (Fragaria × ananassa) plants (Pinto-Poblete et al., 2022). This combination also led to an increased accumulation of Cd in the roots and soil, suggesting that MPs can enhance the bioavailability of certain heavy metals in the soil-plant system. In another study, the combined effects of MPs and nanoparticles on maize (Zea mays) growth were investigated (Yang et al., 2021). The researchers highlighted that certain MPs could promote plant growth, while others, especially at high doses, exhibited strong phytotoxic effects. The presence of both MPs and nanoparticles influenced the soil microbial community composition and diversity, which can have long-term implications for agroecosystems (Yang et al., 2021).
Climate change
The Anthropocene era is also characterized by a change in atmospheric conditions, and as a result, a dramatic shift in climate that has been more pronounced than naturally expected variability. The accumulation of greenhouse gases has exerted its effects on weather patterns incrementally in recent decades. The records being set around the world for extreme weather events, including excessive or prolonged heat and excessive precipitation or drought, have been reported with increasing frequency over several decades as the air and oceans continue to warm. Deviations in temperature and humidity are known stressors for living organisms. Thus, shifting climate patterns, and ultimately seasons, can exert detrimental pressure on homeostasis and reproduction, a focal area of research in food animal and plant species. While polar regions may be more greatly impacted by rising temperatures, range expansion as well as altered growing seasons and migration patterns will influence species sustainability in key biodiversity areas around the world. Climate change, specifically changing temperature and precipitation patterns, is considered one of the most significant threats to the long-term sustainability of biodiversity, food security, and economic growth.
Rising temperatures
Scientists had been aware of the long-term changes in climate (i.e. cooling and warming periods over geologic time) for more than 100 years. Climate fluctuation is a natural phenomenon that has occurred in repeating cycles throughout earth’s history and influenced species and ecosystem adaptations. From the 1960s onward, however, evidence of rapid shifts in climate began to emerge (Weart, 2003). Unbeknownst at the time, this coincided with the post-World War II economic boom that resulted in increased industrialization, urbanization, and fossil fuel consumption. The term “global warming” was coined by Wallace Broecker in 1975 to describe the rapid warming that would result from the increased presence of atmospheric greenhouse gases linked to excessive consumption of fossil fuels, land-use practices and more (Broecker, 1975). Yet, it wasn’t until James Hansen’s seminal report on global warming to US Congress in 1988 that the connection between climate disturbances and human activities was broadcast, including that fossil fuels were driving the “greenhouse effect” (Hansen et al., 1988).
The earth’s atmosphere plays an important role in regulating the planet’s climate and supporting life. A balanced composition of gases, water vapor and particulate matter is critical for optimal heat retention, moisture distribution, and blockage of the sun’s harmful ultraviolet rays. Natural heat retention is necessary to sustain life on earth, but excessive levels of carbon dioxide (CO2) and non-CO2 greenhouse gases in the atmosphere have intensified heat retention resulting in a rapid warming of the planet (Calvin et al., 2023). While increased CO2 emissions from fossil fuel consumption coupled with deforestation-related reductions in carbon sequestration have had a significant impact on atmospheric conditions, fluorinated gases from industrial processes were identified as the key drivers of global warming until restrictions were put in place (Hansen et al., 2000). The 2023 Intergovernmental Panel on Climate Change report stated that the global surface temperature in 2011–2020 was 1.1°C higher than 1850–1900, which corresponded with the highest levels of greenhouse gases in the past 800,000 years (Hansen et al., 2000).
Living systems are capable of some flexibility in thermal tolerance to maintain biological functions; however, elevated ambient temperatures on land and in water can act as potent acute or chronic stressors with marked short- and long-term consequences to metabolism, immunity, reproduction, and more (Sejian et al., 2018). The effects of thermal stress on physiological pathways have been extensively studied, deepening our understanding of an organism’s phenotypic plasticity, or lack thereof, in responding to alterations in homeostasis. Along with the macro-level effects on reproductive health that are described in the section below, researchers have investigated the role of heat shock proteins and other molecular mechanisms in heat-related impacts as reviewed by Boni (2019) .
Rising temperatures and reproductive success
The potential for financial loss and food insecurity has led to a significant number of studies evaluating the effects of thermal stress in food animals and related model species. Hansen’s (2009) review highlighted the damage from both in vivo and in vitro heat stress experiments on hormone production, semen characteristics, sperm DNA integrity, follicular function, oocyte viability and embryo development in cattle, pigs and mice (Hansen, 2009). Laboratory-based studies of experimentally-induced thermal stress in male cavies (Ngoula et al., 2020) and male mice (Zhu and Setchell, 2004) demonstrated that short-term exposure (i.e. 35–60 days) to elevated ambient temperatures can significantly impact sperm parameters with downstream effects on embryo development. In addition to terrestrial mammals, similar outcomes have been observed in aquatic vertebrates. Alix et al. (2020) prepared an extensive review of heat-related consequences on teleost reproductive biology, ranging from altered gamete development to fertilization.
Humans will intentionally seek out warmer climates for leisure activities; however, vacation-based heat exposure does not compare to the increasing heatwaves being experienced in countries around the world. Reports of decreased fertility in human males are primarily related to occupational heat exposure (Thonneau et al., 1998) and seasonality (Lam and Miron, 1996), but in the latter case, the role of heat versus photoperiod could not be effectively delineated. The primary outcome of rising temperatures on human reproduction has been more directly linked to the impact on nutritional resources. Increased female exposure to elevated temperatures during physically demanding agricultural activities along with lower nourishment are known to result in decreased reproductive output through miscarriages and low birthweight offspring (Grace, 2017). Beyond human contexts, food availability is a well-documented driver of reproductive performance across primate species. In-depth reviews of long-term primate studies have shown that variation in nutritional status directly influences both male and female reproductive outcomes, including mating success, offspring survival, and interbirth intervals (Alberts, 2012; Pusey, 2012). These findings demonstrate the critical role of energetic condition and resource predictability in shaping reproductive trajectories across species.
From an evolutionary ecology perspective, Grazer and Martin’s review of laboratory and natural studies also highlighted the negative impact of thermal stress on reproductive traits across all taxa (Grazer and Martin, 2012). Outside of the narrow optimal temperature for reproductive success, minor elevations in temperature resulted in detrimental outcomes that included decreased numbers of offspring, altered mate selection and nesting patterns, and more in species ranging from lions to fishes to insects. Similarly, reports evaluating historical datasets to understand the potential impact of climate change demonstrated the important balance between season length, nutritional resource availability, reproduction, and survival in hibernating or migratory species (polar bears (Molnár et al., 2020), marmots (Cordes et al., 2020), hummingbirds (McKinney et al., 2012)), and American pikas (Ochotona princeps), the latter of which exhibit high thermal sensitivity and are experiencing range contraction due to warming temperatures that impact foraging efficiency and reproductive timing (Beever et al., 2010). The potential for temperature and photoperiod to become uncoupled is cause for concern for species across all taxa exhibiting strict reproductive seasonality. Ectotherms, in particular, are expected to be at high risk for heat-related consequences due to their dependence on environmental conditions to regulate physiological processes and because many of them live at or near their thermal maximums (Sanger et al., 2018). Anole lizard (Anolis carolinensis) eggs exposed to temperatures at their thermal threshold showed reduced embryonic body size, hatching rates, and survival (Sanger et al., 2018). Similar outcomes were observed in the eastern fence lizard (Sceloporus undulatus; (Carlo et al., 2018)), leatherback turtle (Dermochelys coriacea; (Seaman and Milton, 2023)), and loggerhead sea turtle (Caretta caretta; (Fleming et al., 2020)), to name a few.
Interestingly, invertebrates are one of the more widely studied taxa within the context of climate change. Lawrence and Soame (2004) presented an overview of the widespread effects of temperature-based changes on the reproductive output of invertebrates in coastal estuaries that serve as critical stop-over points for migratory birds (Lawrence and Soame, 2004). The impact of rising temperatures on invertebrate sex ratio, timing of spawning, and larval development has downstream effects on available biomass for food for overwintering birds, influencing the birds’ reproductive success as well. In laboratory experiments mimicking heat waves (ecologically relevant temperature elevation over 5–11 days), reduced reproductive rates were observed in the Mediterranean fruit flies (Ceratitis capitata; Grandela et al., 2024), field crickets (Gryllinae; Ratz et al., 2024), and red flour beetles (Tribolium castaneum; Sales et al., 2018). In the beetle study, the researchers observed not only a decrease in sperm production, viability and migration within the female, but also reported the first evidence of transgenerational impact from heat stress with reduced reproductive potential and lifespan of offspring fathered by heatwave-exposed males. The possibility of longer-term transgenerational damage adds another layer of complexity to assessing the consequences of climate change on species sustainability.
As with livestock, thermal stress in plants has been well-studied due to their agricultural value and the potential for substantial crop loss. The findings are similar to animals in which elevated or decreased temperatures have negative effects on reproductive organ development, gamete production, flowering, and more (reviewed by numerous sources, such as Giorno et al., 2013 and Resentini et al., 2023). Although many studies were carried out under experimental conditions, analysis of historical data in natural settings have shown some critical trends. Panchen and Gorelick’s (2017) evaluation of flowering plants in the Canadian Arctic showed a significant advancement in flowering times and seed dispersal times over the past 120 years. A similar assessment in flowering plants in eastern Pennsylvania over the same timespan also documented advancements in flowering times in correlation with rising ambient temperatures (Geissler et al., 2023). At the gamete level, a study on four species of wild catchflies reported a decrease in gametophyte traits and productivity, including antler/ovary lengths and sizes, and seed number and size (Tushabe et al., 2023). Under laboratory conditions, however, a deeper understanding of the mechanisms being disrupted during thermal stress has been attained (reviewed by Resentini et al., 2023). For example, experiments inducing heat stress in rice (Oryza sativa) resulted in irreversible damage to the female reproductive organ leading to sterility and revealed significant changes in metabolic profiles (e.g. sugars, amino acids, reactive oxygen species; Shi et al., 2022).
Altered precipitation
One of the devastating consequences of globally rising temperatures is related to increased evaporation rates, which have resulted in more frequent and intense storms in some regions, and very dry conditions in others. The term rainfall deficit or hydrological drought refers to a period of time with scarce precipitation, which translates to suboptimal water resources on the surface or beneath it (Dai, 2011). Different factors have been linked to the occurrence of droughts. Many studies have shown that human related activities linked to climate change, as mentioned in the previous section, have exacerbated extreme weather. Data reported from Asia found an association between expected changes in flood intensity and increases in air temperature of 2°C (Mirza, 2011). Changes in precipitation patterns can be equally detrimental to the environment, habitats, and the species within them. Consequences such as soil degradation, desertification, plant growth retardation and loss of human life, among others, (reviewed by Shi et al., 2021) threaten food security and economic progress (Gomiero, 2016; Trenberth, 2011). A recent publication found a correlation between the occurrence of drought and a reduction in gross domestic product growth from 0.39% to 0.85% (Zaveri et al., 2023), resulting in economic losses reported in different regions across the globe (United Nations Office for Disaster Risk Reduction, 2021). For example, hydro-meteorological disasters caused $502.56 billion in losses and 90,325 deaths from 1980 to 2017 within Europe alone (Kumar et al., 2020).
Altered precipitation patterns are largely driven by anthropogenic activities, such as deforestation, urbanization, and the consumption of fossil fuels, which disrupt the natural water cycle (Zheng et al., 2023). These activities contribute to increased greenhouse gas emissions, accelerating global warming and enhancing evaporation rates, leading to more extreme weather events, including both storms and droughts. Deforestation reduces the land’s ability to retain moisture, while urbanization increases surface runoff, further contributing to water scarcity in some regions. Additionally, changes in water availability directly affect ecosystem functions by altering both above-ground and below-ground soil microbial and insect communities, which can disrupt the entire soil food web and negatively impact ecosystem health and resilience. For instance, alterations in microbial communities (commonly due to soil pH variations) change ion solubility, nutrient content, and salinity, in turn affecting plant physiology and survival (Nielsen and Ball, 2015). In particular, rhizobial populations have been shown to decrease under dry environmental conditions, affecting the nitrogen fixation abilities and growth of alfalfa (Medicago sativa) and red clover (Trifolium pratense; Dollete et al., 2024).
Altered precipitation and reproductive success
Water availability is fundamental to life, and extreme fluctuations—whether from drought or flooding—induce physiological and environmental stressors that can profoundly impact the reproductive success, development, and survival of species across ecosystems. Drought conditions lead to water scarcity, reduced food availability, and elevated stress levels, affecting organisms at all levels. In contrast, flooding inundates habitats, disrupts reproductive timing, and often leads to unsanitary conditions. These environmental extremes not only shape the reproductive outcomes of plants and animals but also pose significant health and social challenges for humans.
For human communities, drought conditions lead to food and water shortages that can result in economic instability, higher levels of physiological and psychological stress, and ultimately, detrimental impacts on reproductive health. Impaired fetal growth due to malnutrition coupled with limited access to essential resources hampers fetal development and maternal health (Chen et al., 2019; Grace, 2017). Lack of resources underscores the need for resilient health systems, especially to support maternal health during water shortages (Diamond-Smith et al., 2023). In contrast, flooding can inundate communities, posing risks to infrastructure, sanitation, and overall health. Flooding disrupts access to clean water and food, increasing vulnerability to infection, malnutrition, and mental health issues, thereby exacerbating reproductive health challenges with implications for both maternal and fetal wellbeing (Kamal et al., 2018). It is well known that increase in life-threatening scenarios (e.g. unstable infrastructure, unsanitary surroundings and toxic substances; (Giudice et al., 2021)] are commonly associated with depression and post-traumatic stress, which can be detrimental for fetal survival, leading not only to spontaneous abortion, but also long-term consequences for any resulting offspring (Chen et al., 2019; Yüzen et al., 2023). Diamond-Smith et al. (2023) documented that reproductive failure observed as preterm birth and low birth weight were the result of stress, food insecurity, increased risk of developing infectious disease, and blockage to access to maternal social care in countries where unfavorable climatic conditions developed (Diamond-Smith et al., 2023).
Animals, particularly ectothermic species, face selective pressures during drought that directly impact reproductive success. For reptiles like the European adder (Vipera berus), higher embryonic death rates were observed after 2 weeks of water deprivation despite the females maintaining water transfer to the embryos at a cost to their own health (Dezetter et al., 2021). Similar findings were documented in field and laboratory studies of common side-blotched lizards (Uta stansburiana) whereby drought negatively impacted reproductive success in females (Zani and Stein, 2018). Amphibians such as salamanders depend on water-filled ponds for oviposition; drought can result in reproductive failure when these sites dry up (Anderson et al., 2015).
Drought-induced malnutrition in mammals such as lemurs (Propithecus edwardsi) demonstrated the impact of dry weather cycles on reproductive rates (number of offspring surviving to 1 year), resulting in lower fecundity (Dunham et al., 2011). Similarly, ewes (Ovis aries) receiving 50% less water had reduced placental efficiency, commonly determined by the number of cotyledons. As a result, the weights of lambs at birth were reduced (Ocak Yetisgin and Şen, 2020). Mice (Peromyscus californicus) exposed to similar water-deprived conditions developed hypotrophy of the epididymis and seminal vesicles with reduced plasma prolactin concentration (Nelson et al., 1995). In birds, water shortages reduce food availability, delaying parental care and decreasing offspring survival. During persistent dry conditions, bird parents needed more time to search for food resulting in unattended nesting chicks for longer periods. Thus, an indirect link to altered environmental conditions led to increased nestling mortality rates (Schmidt, 1999). Correlations between extreme dry seasons and reproductive failure have also been recorded in four species of passerine birds (Bolger et al., 2005). Flooding also presents reproductive challenges by disrupting animal habitats and increasing predation risks. For birds such as the lark bunting (Calamospiza melanocorys), heavy rainfall and nest flooding lower egg survival and increase predation risks (Skagen and Adams, 2012). Flooding can also render amphibian breeding sites inaccessible, hindering successful egg-laying and subsequent development (Anderson et al., 2015).
Drought conditions are particularly challenging for plants, as they are immobile and heavily reliant on water for physiological functions. Water makes up 80%–90% of plant biomass, and restricted water supply disrupts cellular functions such as photosynthesis, limiting nutrient flow to reproductive organs and reducing flower and seed production (Sánchez-Blanco et al., 2009; Su et al., 2013). Moderate drought (45%–50% soil water content) activates compensatory mechanisms, enabling gene expression responses that allow plants to tolerate dry conditions without compromising yield. However, when soil water content drops to 30%, flower and seed production are significantly reduced (Ma et al., 2014). Consequently, crop yield suffers due to reduced pollen development, lower germination rates, and increased susceptibility to pests, which further hinders nutrient absorption (Etesami and Maheshwari, 2018; Porter, 2005). Drought causes a decrease in cereal production by 9–10% on a global scale (Lesk et al., 2016). For instance, recent publications stated a reduction in rice (Oryza sativa L) and wheat (Triticum aestivum) yield by 25.4% and 27.5%, respectively (Zhang et al., 2018). The effects of drought on reproductive phases might explain these detrimental outcomes. Significant wheat losses due to fewer flowers further exacerbated by female and male sterility have also been documented (Onyemaobi et al., 2017).
Similarly, in tropical and subtropical regions, economic hardship scenarios are commonly reported. African communities have experienced high economic losses when rainfall accumulation is less than 1100 mm per year in territories cultivating banana (Musa spp.). Tropical crops like banana under extreme drought have reduced bunch size and delayed flowering, affecting overall crop quality and increasing economic strain for agriculture-dependent regions (Nansamba et al., 2021; van Asten et al., 2011). In crops like maize (Zea mays), water restriction during late stages can avoid embryo abortion but still results in reduced growth rates and smaller kernel size (McPherson and Boyer, 1977). Similarly, drought-induced stress during crucial developmental phases, such as fertilization, can arrest embryonic development, affecting overall crop yield (Mäkelä et al., 2005; Westgate and Boyer, 1986). While drought is a primary stressor, excess water from flooding can also harm plants, particularly during flowering, by reducing pollinator activity, disrupting nutrient uptake, and increasing susceptibility to fungal infections, ultimately impacting fruit and seed development (Makhmale et al., 2016).
Contaminants exposure interconnected with climate change in reproductive success
Survival and adaptation are foundational principles in biology, deeply rooted in the concept of natural selection proposed by Charles Darwin. All organisms, from humans to animals and plants, have evolved unique strategies to cope with environmental changes, but in today’s world, these adaptations are increasingly challenged by anthropogenic pressures. From a One Health perspective, which emphasizes the interconnectedness of human, animal, and environmental health, it is essential to recognize that threats like environmental pollution and climate change do not act in isolation. Instead, they interact in complex ways, disrupting not only individual species’ survival but also the health of entire ecosystems (Mackenzie and Jeggo, 2019; Figure 3).
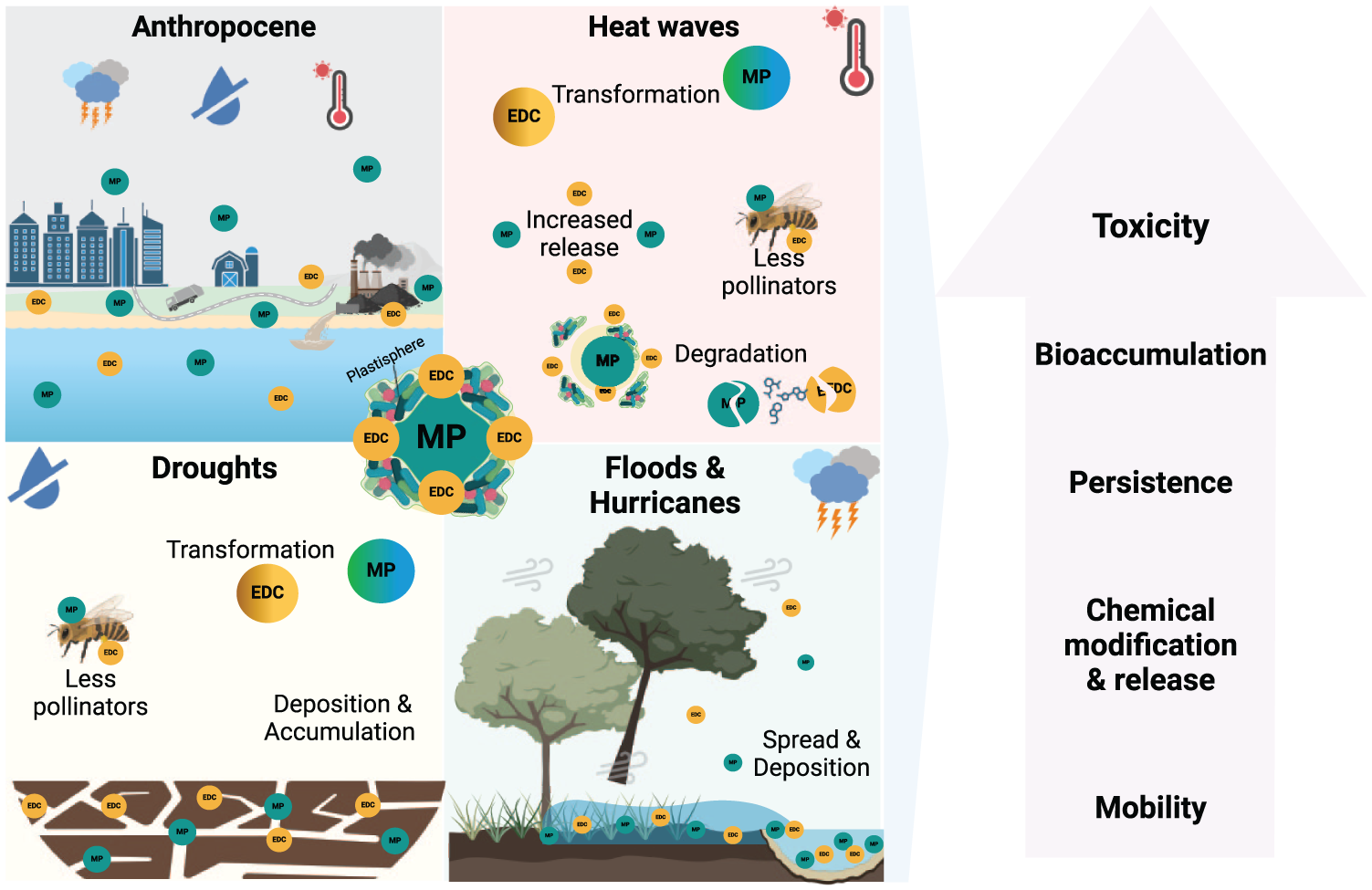
Interactions between EDCs, MPs, and environmental changes in the Anthropocene. Here we show the complex interplay between EDCs, MPs, and various environmental changes exacerbated by climate change, including heat waves, droughts, floods, and hurricanes. The environmental changes contribute to the progressive release, mobility, transformation, and spread of EDCs and MPs, which further intensify their persistence, bioaccumulation, and toxicity in ecosystems. The plastisphere—MPs surrounded by microorganisms, EDCs, and other pollutants—enhances the bioavailability of these harmful substances, increasing their impacts on species and ecosystems. Heat waves can lead to the transformation of EDCs and MPs and increased release of pollutants. Droughts contribute to deposition, accumulation, and further transformation of EDCs and MPs, while affecting pollinators and soil health. Floods and hurricanes spread and deposit pollutants across landscapes, further accelerating the contamination of ecosystems. These interactions drive increased persistence, bioaccumulation, and toxicity of pollutants, which ultimately disrupt hormonal regulation, reproductive health, and species survival, threatening biodiversity and ecosystem function. The interconnected nature of these threats emphasizes the need for a One Health approach, which recognizes the interdependence of human, animal, and environmental health in addressing the compounded impacts of EDCs, MPs, and climate change.
Humans, with their cultural, technological, and cognitive capacities, may seem well-positioned to adapt to environmental changes. However, human health and wellbeing are intricately tied to the health of animals and ecosystems. For instance, the impact of climate change, exacerbated by deforestation, urbanization, and fossil fuel consumption, has altered precipitation patterns, disrupted water availability, and intensified extreme weather events (Calvin et al., 2023). These changes, coupled with pollutants like EDCs and MPs, influence reproductive health and ecosystem function across species. The degradation of natural habitats, soil health, and biodiversity due to these forces undermines food security, economic stability, and health services, affecting humans and animals alike.
The interplay between environmental pollutants, and climate change creates a synergistic threat that compounds their individual impacts on ecosystems and species survival (reviewed by Bolan et al., 2024; de Wit et al., 2022; Holmstrup et al., 2010; Noyes et al., 2009). Climate change amplifies the toxicity of EDCs and MPs by raising temperatures, altering precipitation patterns, and increasing the frequency of extreme weather events. As temperatures rise, bioaccumulation and biomagnification intensify, whereby organisms absorb more pollutants such as EDCs, which then concentrate up the food chain. This leads to more severe disruptions in hormonal regulation, reproductive health, and survival, particularly for species at higher trophic levels, such as large predators. In addition, climate change accelerates the release of EDCs from plastics and other pollutants due to higher temperatures that increase chemical reaction rates. This makes EDCs and MPs more bioavailable in the environment, contaminating soil, water, and organisms more rapidly. Extreme weather events such as floods, storms, and hurricanes also mobilize these pollutants, spreading them into ecosystems and causing a sudden influx of contaminant loads that many species struggle to handle. Plants are similarly affected by the combined impacts of EDCs and climate change. Rising temperatures and altered precipitation disrupt the timing and availability of pollinators, crucial for plant reproduction (Memmott et al., 2007). EDCs further compound this issue by impairing pollinator health and behavior, reducing pollination efficiency and leading to lower seed production and biodiversity loss (Goulson et al., 2015). Additionally, EDCs can interfere with plant hormone signaling, which regulates growth and development, further weakening plant resilience to climate-related stressors.
Microplastics exacerbate the situation by acting as carriers for EDCs and other pollutants (Campanale et al., 2020b). Their large surface area allows them to adsorb chemicals from the environment, which are then ingested by organisms, releasing harmful substances into their systems. This increases the bioavailability of toxicants and magnifies their effects, making it harder for ecosystems to recover. The combined pressures of EDCs, MPs, and climate change impair the reproductive success of species, disrupt food webs, and threaten biodiversity, food security, and ecosystem health on a global scale. Addressing these complex, interconnected threats require a One Health approach that considers the health of humans, animals, and ecosystems as intrinsically linked. Only by acknowledging and mitigating the combined impacts of EDCs, MPs, and climate change can we protect biodiversity, ecosystem services, and overall health across species and environments.
Dispersal disruption and invasive species in reproductive success
Dispersal is a key ecological and evolutionary process that facilitates gene flow, population connectivity, and species persistence across heterogeneous and changing environments (Trakhtenbrot et al., 2005). However, in the Anthropocene, dispersal patterns, especially long-distance dispersal (LDD), are increasingly altered or impaired by anthropogenic activities (Tucker et al., 2018), with significant implications for reproductive success and biodiversity maintenance. Fragmentation of habitats through urban development, infrastructure, and agricultural expansion has led to the physical isolation of populations, reducing the opportunities for genetic exchange and recolonization. Similarly, climate change, through rising temperatures, altered precipitation patterns, increased frequency of extreme weather events, and progressive habitat degradation, continues to destabilize ecosystems, rendering many areas less habitable for native species. These shifts disrupt natural dispersal pathways and reduce population connectivity, exacerbating inbreeding depression, limiting gene flow, and ultimately diminishing reproductive success in many species, particularly those already in decline. (Trakhtenbrot et al., 2005).
For example, the absence of LDD in some rare plant species, such as the endangered orchid Cypripedium calceolus in Europe, has contributed to reduced pollination success and low seedling recruitment due to the fragmentation of suitable habitats and limited seed dispersal by insects (Kull and Hutchings, 2006). In animals, the collapse of natural migratory corridors has similarly constrained dispersal, as seen in large mammals such as African elephants (Loxodonta africana), whose reproductive success is compromised by habitat fragmentation and human-wildlife conflict, which limit gene flow between isolated populations (Blake et al., 2007).
Conversely, human-mediated dispersal has enabled the global proliferation of invasive species through excessive LDD. This is particularly problematic when species are introduced into ecosystems devoid of their natural predators or competitors. These species may then undergo rapid population expansion, outcompeting native species for reproductive resources or directly disrupting native reproductive processes (Traveset and Richardson, 2006). The cane toad (Rhinella marina), introduced in Australia for pest control, rapidly expanded across the continent due to high fecundity and dispersal capability, leading to significant declines in native predator populations through toxin exposure, and indirectly altering predator-prey dynamics essential to ecosystem balance (Shine, 2010).
Reproductive mutualisms are especially vulnerable to disruption by invasive species. In plants, the arrival of non-native species often interferes with native plant-pollinator and plant-disperser interactions. For instance, invasive Carpobrotus edulis in Mediterranean regions not only competes for space and resources with native flora, but its abundant flowers can monopolize pollinator visitation, reducing the reproductive success of co-flowering native species (González-Varo et al., 2013). Similarly, invasive pollinators such as Apis mellifera (European honeybee) can displace native pollinators and alter pollination networks, leading to pollen limitation and decreased seed set in native plants (Aizen et al., 2008). Further, in some ecosystems, invasive seed dispersers or frugivores change the spatial dynamics of plant regeneration. The introduction of Rattus rattus (black rat) to oceanic islands has led to the loss of native seed dispersers and the predation of seeds and seedlings, thereby reducing recruitment of native plants and altering forest composition (Shiels et al., 2014). Moreover, non-native ungulates, such as feral goats and deer, can alter flowering phenology and reproductive organ structure in native plants by browsing, trampling, and altering soil structure (Vavra et al., 2007). These effects cascade into long-term consequences for reproductive success, especially in endemic and narrowly distributed plant species.
The ecological consequences of altered dispersal and species invasions are further compounded by climate change. Invasive species often exhibit broader environmental tolerances and greater dispersal potential than native species, allowing them to capitalize on these new opportunities and further disrupt native reproductive dynamics (Hellmann et al., 2008). For example, warming temperatures have facilitated the expansion of invasive cheatgrass (Bromus tectorum) in North American deserts, where it displaces native grasses and alters fire regimes, leading to reduced reproductive success and recruitment of native perennials (Balch et al., 2013).
Implications for the future
In the coming decades, addressing the complex intersections of pollution, climate change, and reproductive health will be essential for protecting biodiversity and ecosystem stability. Anthropogenic impacts have placed unprecedented pressure on reproductive success across species, which is fundamental to sustaining healthy ecosystems and supporting global biodiversity. As these challenges persist, new approaches to managing environmental health, with a particular focus on reproductive resilience, will be crucial. Global efforts to meet environmental and biodiversity targets will become increasingly critical over the next decades. It is vital for nations to remain committed to establishing realistic and enforceable national frameworks that address the drivers of the ongoing environmental crises. The research community must continue to develop species-specific metrics to assess reproductive success and create tools to mitigate reproductive health challenges across ecosystems. Ensuring the survival and health of species in the face of pollution and climate change is essential to safeguarding biodiversity, ecosystem function, and ultimately, human wellbeing.
Footnotes
Acknowledgements
We acknowledge Dr. Ana Cristina Tamayo, professor specialized in Applied Genetics, for reviewing the manuscript. Figures were created in BioRender.
Author contributions
GFM: conceptualization, writing - original draft and review, project administration, funding acquisition. MAMMF: conceptualization, writing - original draft and review. DRB: writing – original draft and review. LAGG: writing – original draft and review. CAL: writing – original draft and review.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This manuscript was made possible by funding from the Natural Sciences and Engineering Research Council of Canada [GFM].
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.