
Abstract
As the continent with the deepest record of human history, the relationship between landscape formation and human subsistence practices is inseparable. The activities that constitute ‘farming’ are open for some matter of discussion, but essentially speak to the co-evolution of human, plant and animal reproductive systems along a continuum of interdependence. This process is ever evolving, but has resulted in the formation of landscapes in which anthropogenic processes characterize ecosystem functionality in nearly all biomes on the continent. Practices of cultivation, broadly conceptualized, across Africa are varied in the ways in which they have transformed ecological systems and landscapes. The use of fire as a landscape management tool dates to the Pleistocene, and penning of wild sheep to the early Holocene. The alteration of the landscape of fear by these types of human activities had fundamentally restructured trophic systems in Africa prior to the introduction of agriculture. However, the introduction of animal herding and intensive forms of plant cultivation by the middle Holocene correlated to even more significant ecological changes. The creation of agricultural landscapes has had a negative impact on biodiversity in some locations, whereas other some practices at different points in time have positively affected biodiversity. It is now recognized that humans have long influenced the evolution of landscapes wherever they live, but the current research focuses on where, when and how socio-ecological processes become coupled in the palaeoecological record.
Keywords
Introduction
The transition of human subsistence practices based entirely on hunting and gathering to one based on agriculture has been commonly referred to as ‘revolutionary’ for our species. The emergence of agriculture went hand-in-hand with population growth, the growth of cities, transportation and writing systems, significant biological changes in human microbial taxa and body morphologies and a radical reorganization of earth’s biome as the scale of land clearance accelerated towards the end of the Holocene, ca. 12,000 years ago (ka; Ellis et al., 2021; Stephens et al., 2019). The culmination of this process has been coined as the ‘Anthropocene’, a geologic epoch of earth in which human-driven formation processes outstrip natural-driven processes across the entire face of the planet (Crutzen and Stoermer, 2000). A formal designation of the Anthropocene epoch by the Anthropocene Working Group (AWG) of the International Union of Geological Sciences has been established as beginning in the early 1950s based on the presence of artificial radionuclides in sediments, which heralds the advent of the nuclear age (Zalasiewicz et al., 2021). The ca. 1950 chronostratigraphic marker (or ‘Golden Spike’) of the Anthropocene corresponds to what some have called the ‘Great Acceleration’ in which there are myriad detectable anthropogenic impacts on earth’s geologic framework as a result of cumulative deforestation, industrialization and urbanization (Steffen et al., 2007, 2015). However, farming systems did not emerge rapidly or organically, nor are they completed – they have long antecedents in human land management practices that extend deep into the Pleistocene and they are subject to ongoing evolutionary processes in adjustment to changing realms of biodiversity and climate change (Boivin et al., 2016; Hoag and Svenning, 2017).
This paper will provide a critical thematic review of the roots of farming and evaluate its evolutionary impacts on the formation of landscapes over long time cycles on the African continent leading up to the present day. Following a brief overview of the continent’s diverse geography and how farming itself can be defined in this context, the application of ecological theories of fear and trophic cascades in the context of African farming systems will be outlined. The paper then explores two broad, transformative socio-cultural phenomena: the termination of the African Humid Period (AHP, ca. 8–4.5 ka) and the emergence of the Iron Age (ca. 5–2 ka), both of which significantly impacted the formation of the landscape of pre-colonial Africa and have influenced the complexion of socio-ecological systems today.
The coupling of sociological processes with impacts detectable in the palaeoecological record can be difficult to disentangle with precision, but as humanity’s evolutionary homeland, the African continent hosts landscapes from which the longest records of human-environmental impacts can potentially be traced (Boivin et al., 2016). Farming systems are ecological niches that have been forged over millennia of co-evolution between humans and the myriad species with which we interact. The purpose here is not to argue for or against the AWG definition of the Anthropocene as a geologic epoch, but rather to contextualize the emergence of the present landscape using a historical ecological approach. The research programme of historical ecology situates human-mediated disturbances to landscapes as critical features of ecological succession (e.g. Balée, 2006; Crumley, 2021; Lane, 2010) and is therefore not looking for a ‘Golden Spike’ marking the emergence of an epoch, per se. As will be argued below, the impacts of human land tenure are measurable from the diachronic view afforded by humanity’s long history in Africa on millennial timescales (sensu Lane, 2019) and important for understanding what may lie ahead (Anderson et al., 2006; Fraser et al., 2007; Nogués-Bravo et al., 2018).
The Anthropocene has become understood to be far more than a geologic phenomenon – it has come to assume significant political and moral implications as it is the only geologic epoch to be defined by, as well as caused by, a single species (Burdon, 2020; Lewis and Maslin, 2015). Critics of the Anthropocene concept argue that it is the only geologic epoch which is simultaneously denaturalized and ‘renaturalized’ being caused unevenly by humans (beneficiaries of global capitalism being the prime causal agents), but presenting concomitant global impacts (Malm and Hornborg, 2014). Another critique is that the Anthropocene is stratigraphically and conceptually indistinguishable from the Holocene since the latter epoch is primarily characterized by anthropogenic formation processes, particularly in the second half, and the distance afforded by time to observe changes in the sedimentary record are not yet available (Gibbard and Lewin, 2016). Understanding that there are epistemological considerations of what the Anthropocene entails, there is growing scientific consensus that the geologic Anthropocene has antecedents and studying its roots offers the best path towards developing a future with levels of biodiversity that can sustain robust ecosystem functionality (Brondizio et al., 2016). Land host management practices that emphasize conservation of indigenous, sympathetic plant and animal species have proven to create invaluably durable islands of productivity against soil and biologic degradation (Kendall and Drew, 2019; Sinclair et al., 2019). From this vantage point, this paper critically reviews ecological (niche) construction across Africa as an extension of long-term co-evolutionary dynamics. Statistical modelling of previously published data will be undertaken, but the primary objective of this review is to use case studies to understand how recursive ecological regime shifts can unfold, feeding into one another over space and time as niches are constructed, respond and are further modified to accommodate biotic and abiotic pressures that constitute them.
Geographic setting for farming systems in Africa
Africa is a continent of extraordinary diversity in geology, topography and climate systems, which has resulted in a wide array of biodiversity among the endemic plant and animal taxa (Figure 1). The intricate interrelationships between bedrock geology and soil systems to plant and animal communities were analysed by Bell (1982) in a landmark paper, which was one of the first to clearly hypothesize the correlation between nutrient-rich soils of the volcanic plains and the occurrence of grassy savannah lands in much of the subcontinent. Grasslands are perpetuated by attracting grazers who spread seeds in their dung to fertilize the plains, maintaining the savannah ecosystem. In this way, if water availability and slope are held constant, igneous (volcanic) landforms predict the occurrence of grasslands (Bell, 1982). On the other hand, the older, weathered bedrock landforms of metamorphic and sedimentary formations possess fewer nutrients available to plants, which Bell (1982) hypothesizes encourages the storage of scarce nutrients above the ground in forests (e.g. miombo woodlands, tropical canopy), attract browsing animals such as elephants (Loxodanta sp.) and giraffes (Giraffa camelopardalis), who maintain woodland communities. These trophic webs interfinger with one another across Africa and have created differentiated ecological areas from which a diverse range of human subsistence practices has emerged.
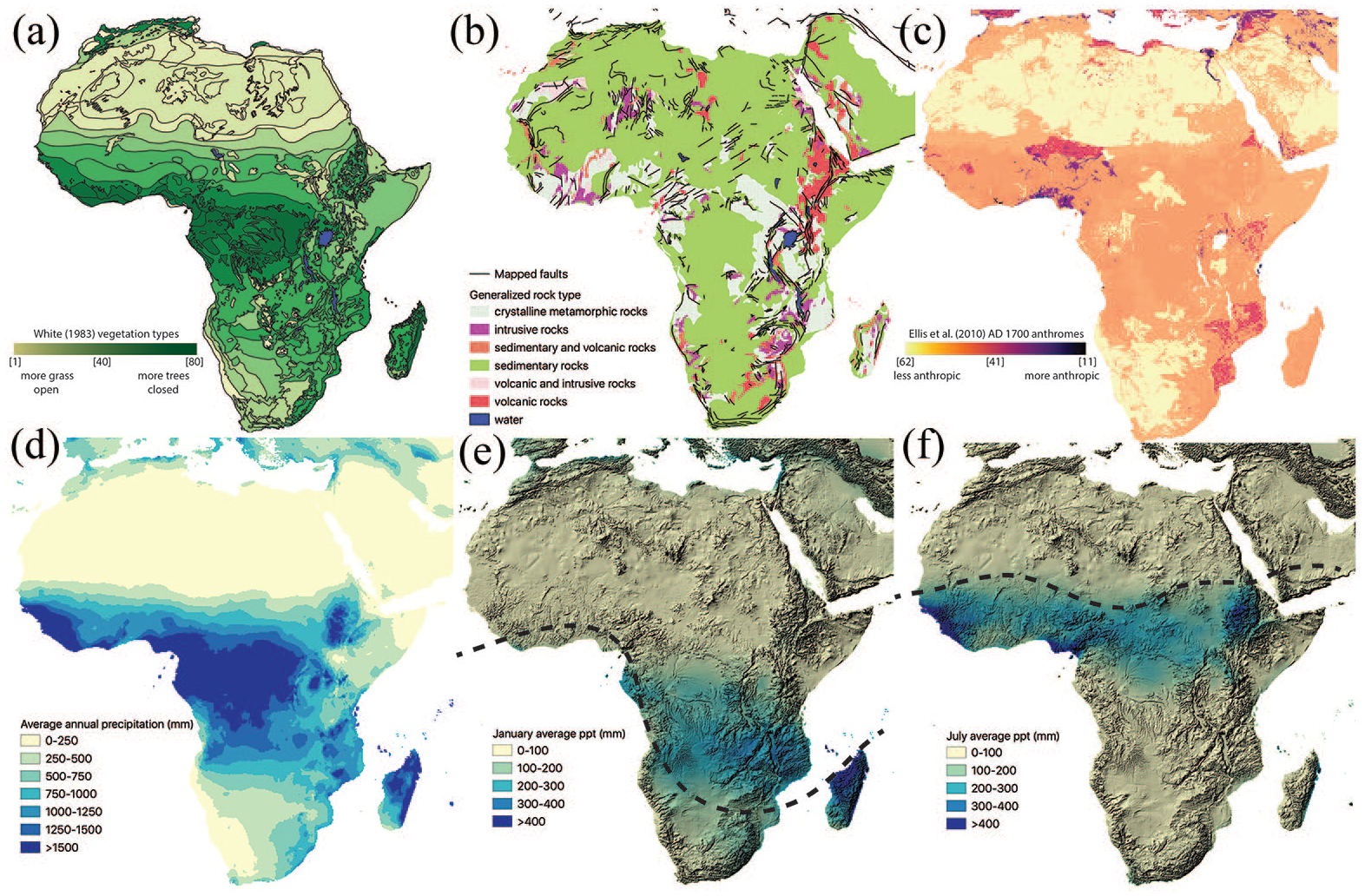
Maps of African geography. (a) Vegetation distributions as mapped by White (1983). The units are mapped in chloropleth distribution from 1 to 80 according to White (1983: Table 4, pp. 57–66), excluding water; (b) Geology supergroups mapped by the Geological Survey of Canada (Chorlton, 2007). Faults mapped and distributed by ESRI in 2001; (c) Anthrome construction ca. AD 1700 as mapped by Ellis et al. (2010) – see also Supplementary Information in Ellis et al. (2021); (d) Annual average precipitation from 1970 to 2000 (Fick and Hijmans, 2017); (e) January average precipitation 1970–2000 (Fick and Hijmans, 2017) with GTOPO30 hillshade background (www.doi.org/10.5066/F7DF6PQS). Dashed line shows the estimated average position of the ITCZ; (f) July average precipitation 1970–2000 (Fick and Hijmans, 2017) with GTOPO30 hillshade background (www.doi.org/10.5066/F7DF6PQS). Dashed line shows the estimated average position of the ITCZ.
A stronger predictor of plant communities than geology is rainfall and topography, which, in the tropical regions of Africa, are strongly dependent upon one another. The long axis of the African continent (between 37°N and 35°S) falls roughly equidistantly across the tropical belt of the Equator along a north-south trajectory. With the annual migration of the Earth and associated precessional tilt towards and away from the sun relative to the reference aspect taken, there is a ridge of converging low pressure known as the Intertropical Convergence Zone (ITCZ) that follows the area of maximum annual insolation. Rainfall associated with monsoonal convection from the oceans towards the inland regions occurs along the ITCZ in the tropical regions of the continent. Given the predictable annual cycle of Earth’s orbit, there is a patterned annual distribution of rainfall associated with the migration of the ITCZ. The Mediterranean and cone of southern African are influenced by different higher/lower latitude atmospheric pressure cells bringing winter/summer rainfall, respectively.
The distribution of precipitation is not uniform across space due to the effects that elevation has on condensing moist air, and elevation is highly variable in eastern half of Africa due to continental rifting processes (Hession and Moore, 2011). The East African Rift System (EARS) has formed over the last ca. 40 million years as the East Africa Plate has separated from the Somali Plate via a series of complex faults and tectonic processes (Tiercelin and Lezzar, 2002). The EARS runs south from the modern-day country of Israel/Palestine forming a trough in the Red Sea, into Ethiopia after which it bifurcates into east and western branches that extend to Malawi (Figure 1b). Extended pushing and pulling of landforms has resulted in extreme contrasts between highland and lowland settings across relatively small distances. Because moisture condenses when air is cooled, the highlands create updraft zones for the convection of tropical moisture that condenses on the windward side of uplands, leaving the leeward side drier. Central and western Africa contain large low-lying river basins (Niger, Congo) that are split by the Yola Rift and have seasonally high annual rainfall. Most of southern Africa is semi-arid to arid in high elevation settings except along the coastlines, which have Mediterranean type climates. The circumstances of this geography have created a landscape of variable resource distributions, which have been hypothesized as foundational to the emergence of our species as bipedal, large-brained apex predators (Potts, 1996).
What is a farmer?
To better understand the impact of farming on landscape formation, an extended definition of ‘farming’ will be proffered here. In the post-Industrial era, contrasts between ‘untouched’ tropical landscapes of central Africa and highly managed commercial viticulture regions of southern Africa or pastoral grasslands of eastern Africa seem rather stark. However, past landscapes and subsistence practices were not so clearly separated, and that which constitutes a ‘farmer’ or a ‘forager’ is a matter of interpretation and occasional vigorous debate (Crowther et al., 2018; Kusimba, 2005; Terrell et al., 2003). Modern notions of property and tenancy introduced during the colonial era have demarcated land tenure and management practices into distinct zones, which were rare prior to ca. 100 years ago.
The simplest definition of a farmer is a person who practices agriculture using domesticated taxa. The humans, plants and non-human animals engaged in farming generally benefit from an evolutionary fitness perspective, and therefore ‘domestication’ is not a process of human dominion or co-opting of nature, but rather co-evolution that is ongoing and lies along a spectrum of expression (Bogaard et al., 2021). The production cycle of farming consists of four crucial elements: (1) propagation, (2) husbandry, (3) harvesting/slaughter and (4) storage/maintenance (Harris, 2007). Whether they are plants or animals, a farmer is engaged in management over the stages of their foods’ life cycles from seed to the point of consumption. This definition stands in contrast to a forager, who may engage in one or more of these elements, but not all simultaneously. However, as Flannery (1968: 68) argues, every forager knows that if a seed is placed in the ground, a plant will grow. Armed with this knowledge, foragers have been actively managing landscapes since time immemorial, since doing so has been the crucial element to human evolutionary fitness (Boivin et al., 2016; Thompson et al., 2021a).
Agronomists with the World Agroforestry Centre (ICRAF) have identified five primary farming systems in Africa: Highland Perennial (HP), Maize Mixed (MM), Cereal Root Crop (CRC), Agro-Pastoral (AP) and Highland Mixed (HM), which is based on the Food and Agriculture Organization of the United Nations land use scheme (Figure 2; Dixon et al., 2001; Garrity et al., 2012). Their analyses identify HP systems as primarily residing in eastern Africa and are largely based on cash and staple crops (especially maize and coffee) in densely populated upland settings. HP systems are less dependent on rainfall than other systems due to their highland settings where precipitation is generally more stable than in lowland settings. MM systems extend over 91 million hectares of eastern and southern Africa, colonizing formerly forested regions within subhumid zones. MM systems heavily focus on legumes, maize and cash crops for export with household livestock rearing. CRC systems are located in the Guinea-Savanna vegetation zone (White, 1983) south of the Sahara. The cereal, root and tuber crops that constitute the vast majority of the productive output of the CRC region are heavily dependent on increasingly precarious rainfall. The region is classified as ‘parkland’ with large stands of tree canopy remaining from which people collect shea nuts, but crop yields in CRC systems are generally lower than in other regions. AP systems are located in the semi-arid, drought-prone regions of northern, eastern and southern Africa. They may include limited cultivation of pearl millet (Pennisetum glaucum) and sorghum (Sorghum bicolor), but are primarily focused on grass conservation and livestock production. Traditionally AP systems relied on seasonal transhumance, but there has been a growing shift to sedentary forms of farming as land tenure systems have become increasingly regulated. Finally, HM farming systems occupy over 40 million hectares in portions of Ethiopia, Eritrea, western Cameroon, Lesotho and Angola. The cool and humid climate conditions have encouraged extensive land clearance for cereal crop production and intensification of farming practices for high-yield outputs. These five systems are practiced by over 70% of the population of Africa at the time that the ICRAF report was generated and have deep roots in historical land management practices (Garrity et al., 2012).
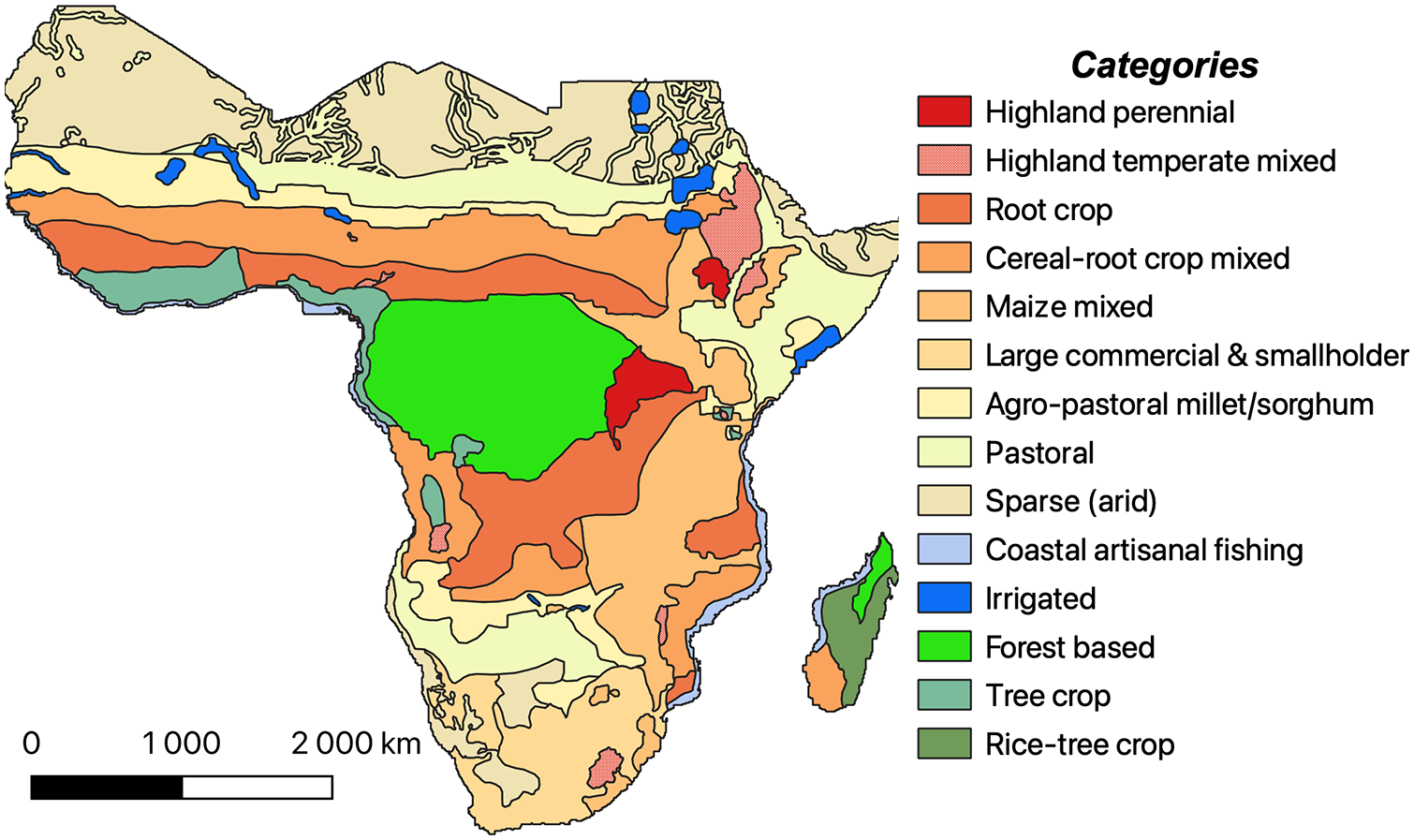
Map of sub-Saharan Africa showing dominant farming systems based on data from Dixon et al. (2001) and updated by Garrity et al. (2012). Shapefiles published by the Food and Agriculture Organization of the United Nations (Auricht and Dixon, 2015).
Like all lifeforms, humans take part in networks of ecological niche construction, whereby reciprocal choices and actions taken by organisms for subsistence and/or reproduction alter the habitats of one another. Niche Construction Theory (NCT) argues that a feedback loop develops between different organisms in an ecosystem in which symbiotic forms of habitat alteration becomes mutually beneficial to a certain range of organisms, which accelerates over time under stable climatic conditions (Laland et al., 2016). Using NCT, landscape formation can be viewed as a multipronged, recursive process in which different actors exert influence on the biome. Returning to Bell (1982), the occurrence of grazers on nutrient-rich volcanic soils promotes the propagation of grasses, which, in turn, further encourage the presence of grazers and myriad sympathetic species. A critical element of NCT is the concept of ‘ecological inheritance’ in which the evolutionary dynamics between the components cohabiting a landscape reflect long legacies of natural selection suited to the co-engineered habitat (Laland et al., 2016).
The practice of agriculture constructs an ecosystem (niche) that is ultimately dependent on the continuance of its four constituent elements (listed above), however farming practices depend on critical antecedents from managed landscapes inherited from foragers. Beyond the immediate plot under cultivation or patch of grass being grazed by a domesticated animal, a regional ecology called an anthrome evolves in response to the selective pressures exerted via anthropogenesis. Mapped estimations of anthrome extents in Africa ca. AD 1700 indicate that a relatively small portion of the African continent had been evidentially transformed fully to anthromes by this time (Figure 1c), however the evidence for ecological transformations is difficult to pinpoint in time and space (Ellis et al., 2010, 2021). Garrity et al. (2012: 49) largely attribute farming systems in modern Africa to postcolonial-era rural development programmes and economic structural adjustment policies, but there is a considerable body of research that finds deep connections between modern agricultural practice and precolonial farming systems (Altieri et al., 2015; Edwards et al., 1993; Samberg et al., 2013; Wright et al., 2019). Domesticated landscapes are boundless and recursive, in a state of constant co-evolution between gradients of wild, feral, domesticated and domesticate-sympathetic species, and require deep understandings of the subtle ecological drivers that form landscapes. Given the imprecision of proxy records to understanding past landscape formations, ecological theory provides a solid foundation to inform the impact of agriculture on landscapes.
Ecologies of fear
This review critically applies historical ecological theory to understand niche formation in the context of human technological developments beginning with the introduction of farming systems across the African continent. The Darwinian model of natural selection among organisms is predicated on differential rates of reproduction based on fitness within a trophic system (Darwin, 1859). According to Darwin and over a century and a half of subsequent evolutionary biological research, organisms better adapted to an environmental context have a better chance of reproducing compared to those who are not as well adapted. A critical component of the trophic system is the tense relationship between predators and prey – the former balance the reproductive success of the latter to maintain quasi-equilibrium when viewed intergenerationally (Brown et al., 1999; Rosenzweig and MacArthur, 1963). ‘Prey’ in all their various forms have evolved to avoid predation and adopt elusive strategies to maximize their reproductive fitness. Within the context of the broader landscapes they inhabit, this behavioural dynamic is mitigated by the ‘ecology of fear’ in which there is a continuum between N-driven (population size) versus μ-driven (fear) systems (Brown et al., 1999; Rosenzweig and MacArthur, 1963). In μ-driven systems, the predator reduces the number of prey mostly by fear when driving them out of preferred habitat into suboptimal foraging patches rather than by killing them. Humans have had deep histories of entanglement in the evolutionary ecologies wherever we go and have profoundly influenced the biology of a wide range of species based on size- or trait selective harvesting techniques and defaunation dating deep into the Pleistocene (Sullivan et al., 2017). On a regional level, Thompson et al. (2021a) apply this model to fire-sculpted ecologies created by foragers in northern Malawi (Figure 3) after 92,000 years ago (see also, Thompson et al., 2021b). However, fear-based ecologies are not the exclusive domain of hunter-gatherers.
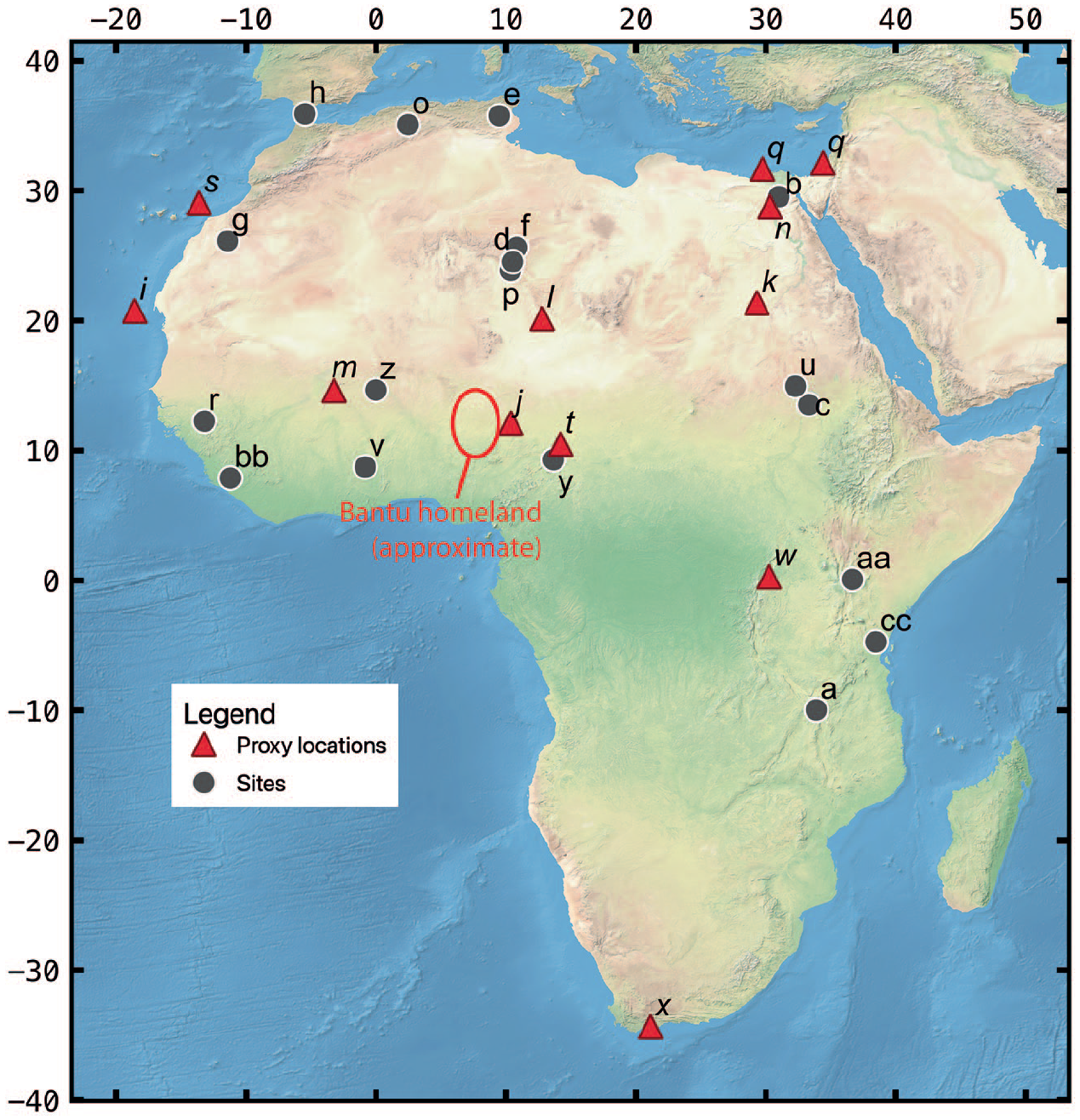
Locations of human or palaeoecological reconstruction discussed in order of the text: (a) Northern Malawi Pleistocene foragers (Thompson et al., 2021b); (b) Fayum Depression (Linseele et al., 2014); (c) Jebel Moya (Barich, 2014; Brass, 2018; (d) Takarkori (Dunne et al., 2018) and Wadi Teshuinat (Mercuri, 2008); (e) Tunisian Maghreb (Mulazzani et al., 2016); (f) Mezzak Plateau (Gallinaro and di Lernia, 2018); (g) Zemmur region (Borrell et al., 2018); (h) El-Khil and Kaf That el-Ghar caves (Martínez-Sánchez et al., 2018a, 2018b, 2021; (i) GeoB7920-2 sediment core (Dupont and Schefuß, 2018); (j) Lake Tilla (Salzmann et al., 2002); (k) Selima Oasis (Haynes et al., 1989); (l) Segedim Depression (Schulz, 1994); (m) Ounjougou (Huysecom et al., 2004); (n) Sai Island (Florenzano et al., 2019); (o) Ifri Oudadane (Morales et al., 2013) and Ifri n’Etsedda (Linstädter et al., 2016); (p) Tin-a-Hanakaten (Aumassip, 1984); (q) Nile River runoff proxies (Blanchet et al., 2015; Hennekam et al., 2014; (r) Gajiganna (Höhn et al., 2021); (s) Canary Islands (von Suchodoletz et al., 2010); (t) Mandara Mountains (Diaz et al., 2018); (u) Butana Group (Out et al., 2016; Winchell et al., 2017; (v) Kintampo Group (Logan and D’Andrea, 2012) (w) Wandakara (Russell et al., 2009); (x) Gourits River terraces (Damm and Hagedorn, 2010) (y) Garoua (Wright et al., 2017); (z) Iron Age Burkina Faso sites (Höhn and Neumann, 2012) (aa) Laikipia Plateau (Muiruri et al., 2022); (bb) Gola Forest (Kandeh and Richards, 2011); (cc) Usambara Mountains (Msuya et al., 2008; Munishi et al., 2008) and coastal Kenyan sites (Culley et al., 2021; Wright, 2007). Palaeoecological sites are connoted in italicized font and red triangles.
Human systems of food production have long been recognized as having transformative effects on the environment (Aschmann, 1973; Helbaek, 1960; Montgomery et al., 1973; Sherratt, 1980). Land clearance for planting or pasture, water diversion for irrigation and terracing represent three of the myriad ways in which humans changed the natural land surface agriculture to cultivate food. The knock-on effects of these activities include enhanced soil erosion and salinity, eutrophication, deforestation, chaparral growth and eventual urbanization. In addition, the selective removal of non-productive species (called ‘weeds’ or ‘vermin’ or even ‘predators’) is foundational to the construction of a niche that is good for the perpetuation of farming systems. Modern ecological studies frame these relationships as human-wildlife conflicts or interactions (e.g. Manfredo and Dayer, 2004), but structural adjustment of wildlife relative to a human presence is as old as the niches within which we evolved (Sullivan et al., 2017).
Examining the underpinnings of the conversion of the landscape to one in which natural (non-anthropogenic) processes predominated more of the African continent’s functional ecology to one in which farming systems (plant and animal agriculture) are the dominant drivers requires a reframing of understanding trophic webs. The question is whether farming systems of the Iron Age, or earlier, conceptually represent an initial phase of the Anthropocene, which geologists have largely resolved is the epoch in which we are currently living (Zalasiewicz et al., 2021). Oft-cited counter arguments to an early Anthropocene before AD 1950 are that population and impact levels of early farmers were too low to induce recursive, continental-scale anthropogenic effects (Steffen et al., 2007), and natural variability in solar radiation and atmospheric CO2 concentrations account for pre-industrial climate conditions (Steffen et al., 2011). However, early farming techniques involved far more land clearance units per capita compared to Industrial-Era agriculture (Ruddiman, 2007; Ruddiman and Ellis, 2009) and methane production from early farming practices were likely significant on a global scale (Fuller et al., 2011). The incomplete nature of the archaeological record, particularly in Africa, impedes precise scaling of changes in land cover (Kay and Kaplan, 2015; Morrison et al., 2021), but the anthrome ecologies of today have clear historical antecedents.
The introduction of new trophic dynamics into an ecological system undergoing extrinsically driven change (e.g. aridification, burning, salinization, grazing) accelerates the race towards reaching a tipping point in which the ecology enters a new biofunctional state (Andersen et al., 2009; Folke et al., 2004; Kinzig et al., 2006). Behaviourally Modified Trophic Cascades (BMTC) are the indirect ecological impacts of predation or disturbances that occur outside the zone where direct impacts are detected (Schmitz et al., 2004). Agricultural technologies, particularly from by plant-cultivating Iron Age farmers, were accompanied by more settled residential practices. Humans are alpha predators with advanced lethal technologies that other members of the animal kingdom take exceptional care to avoid (Frid and Dill, 2002; Zbyryt et al., 2018). Thus, farming settlements were places for many species of wildlife to avoid, while other sympathetic species such as mice, scavenging carnivores and certain species of monkeys are attracted to these zones. In the case of the introduction of domesticated plants or animals into a novel ecosystem, the direct impacts of land clearance and husbandry as well as indirect impacts related to human settlement and land investiture associated with agriculture are agents of significant landscape transformation akin to what is called the Anthropocene, but may be difficult to detect in the palaeoecological record (Braje and Erlandson, 2013; Ellis, 2011; Ruddiman, 2013). In order to link socio-ecological processes for the purpose of assessing when, where and how anthromes were created, we must rely on inferences drawn from places with long archaeo-ecological records from which substantial landscape transformations have been documented and may have been influenced by novel forms of human technology.
Neolithic pastoral ecologies of northern Africa
There is probably no single ecosystem on Earth that witnessed an ecological transformation in the pre-Industrial Era at a scale substantially comparable to that which took place in the Sahara during the Holocene. At 10 ka, the Sahara was a complex mosaic of savannah and forests and hosted thousands of lakes (Hély et al., 2009). This was the so-called AHP – a period of extended high rainfall across the tropical northern latitudes that was the result of a more northerly placement of the ITCZ relative to today (DeMenocal and Tierney, 2012; Gasse, 2000). Due to chronic undersampling, prehistoric precipitation levels are difficult to estimate, but vegetation reconstructions indicate that the ITCZ was between 5° and 7° north of its present position (Hély et al., 2014). Simulation models suggest overall precipitation may have been as much as 25% (Kutzbach and Liu, 1997) to 40% (Prell and Kutzbach, 1987) higher than at present, which is consistent with proxy data (Gasse, 2000; Tierney et al., 2017). The mosaic landscape was discontinuously dotted with wetlands and small lakes (Quade et al., 2018), which was a significantly more attractive biome for human habitation than present conditions. However, the Sahara is vast and topographically diverse, and there were many places within the Sahara during the AHP that had ecologies relatively similar to those that are found today (Hildebrand et al., 2018; Zerboni and Nicoll, 2019).
The timing and geography of the termination of the AHP is a matter of debate with some proxy evidence indicating an abrupt transition from wet to dry conditions (e.g. Bloszies et al., 2015; Bristow et al., 2018; Collins et al., 2017; DeMenocal et al., 2000; Salzmann and Hoelzmann, 2005) and some that indicate a slower, step-wise transition (e.g. Francus et al., 2013; Kröpelin et al., 2008; Lézine et al., 2005; Neumann, 1989; van der Lubbe et al., 2017). Climate models consistently underestimate the speed with which many regions of the Sahara became arid relative to the proxy record (Claussen et al., 2017). Poor parameterization of climate models accounts for some of the discrepancy and most recently the problems are overcome by structuring in ‘tipping points’ triggered by changes in land-atmospheric feedbacks (Hopcroft and Valdes, 2021), but the pace of aridification in some portions of northern Africa appears to have exceeded orbital forcing alone.
Proxy evidence shows a general north-to-south termination of the AHP due to the failure of the summer monsoon to progressively penetrate inland relative to Atlantic Ocean moisture sources (Gasse, 2000; Tierney et al., 2017). Concurrent to changes in the moisture balance, there is a progressive shift in plant community composition from tropical woodland/grassland mosaics that characterized the AHP to herbaceous, xerophilic communities beginning after 8 ka, but accelerating between 7.4 and 7 ka (a detailed review of the evidence is provided in Hildebrand et al., 2018). In the Nubian aquifer of the southeastern Sahara, the presence of evaporitic sediments indicates aridity beginning ca. 5.7 ka, reaching modern conditions by 4.5 ka (Pachur and Hoelzmann, 2000). Lake evaluation indexes, analyses of δ18O isotopes and clay mineral characterization from central Mediterranean sapropels, capturing runoff from Libyan, Algerian and Tunisian drainageways, converge on a general interpretation that termination of the AHP initiated after 6.5 ka with the most intense phase of aridification occurring between 5.5 and 5 ka (Drake et al., 2018; Wu et al., 2017). Across western Africa, a time-transgressive termination of the AHP is identified in proxy records from 20.8°N to 5.5°N dating from ca. 5.5 to 3 ka (DeMenocal et al., 2000; Shanahan et al., 2015). The dominant driver of the transition from humid to dry conditions was changes in orbital conditions, which displaced the ITCZ from a more northerly to a more southerly position.
However, the termination of the last AHP is comparatively remarkable because the tempo of the ecological transition was so variable and spatially discontinuous, particularly in the Sahara (Collins et al., 2017; Itambi et al., 2010; McGee et al., 2013). A bivariate bagplot of a 3000-year running average of 3 million years (Myr) of orbital precession (Berger and Loutre, 1991) vs. a northern African humidity index based on elemental analyses of a Mediterranean sediment core (Grant et al., 2017) demonstrates that the period from 8.5 to 5 ka is outside the fence of disparity of the 5993 data points analysed (Figure 4). A Cook’s Standard Distance bivariate analysis likewise demonstrates that the terminal AHP in the Holocene was an outlier in relation to normalized climate parameters over the last 3 Myr (Figure 5). Other outlying climate conditions occurred within the last ca. 1 Myr, some of which are tied to well-known non-anthropogenic forcing factors (e.g. Mount Toba eruption of 74 ka) or were during extreme climatic anomalies within the geologic record (Marine Isotope Stage [MIS]6, MIS5e). It is important to underscore that the discordant spatial-temporal records of slow versus rapid termination of the AHP confound a simplistic interpretation of broad-brush regional statistical data.
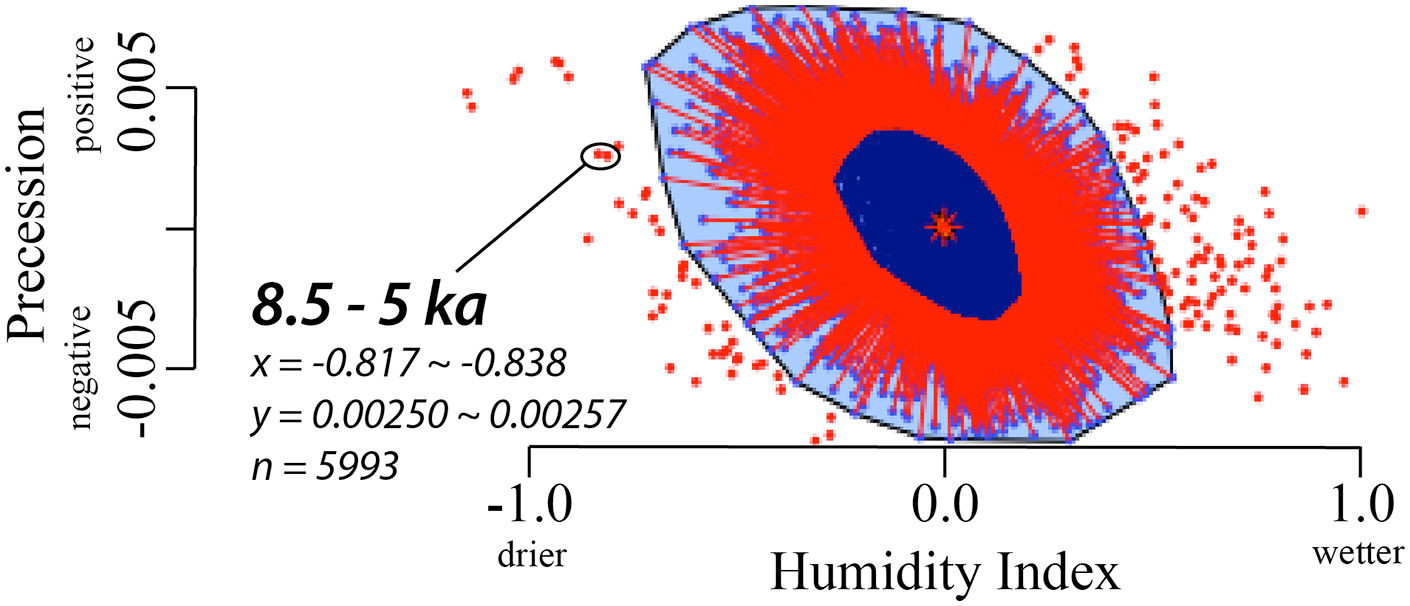
Bivariate bagplot run of 3000-year running averages over 3 Myr of orbital precession (Berger and Loutre, 1991) versus a northern African humidity index (Grant et al., 2017) in aplpack in R. Values outside the fence (>2.5-σ) are classified as outliers. Data and R code for the statistical model is available at https://doi.org/10.6084/m9.figshare.16964947.v1.
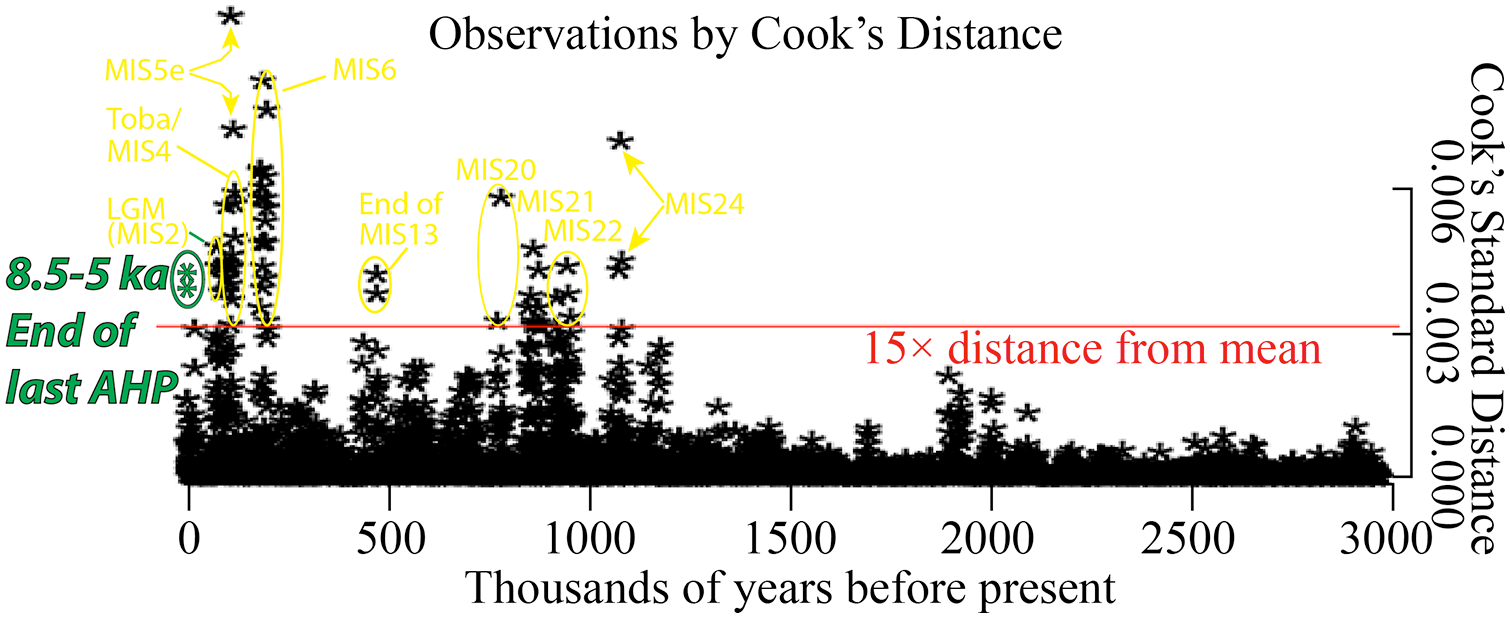
Cook’s Standard Distance run of 3000-yr running averages over 3 Myr of orbital precession (Berger and Loutre, 1991) versus a northern African humidity index (Grant et al., 2017) in carData in R. Outlier values relative to 15× distance to the mean value are identified. Greater Cook’s Standard Distance indicates greater disparity between orbital and humidity indexes. Data and R code for the statistical model is available at https://doi.org/10.6084/m9.figshare.16964947.v1.
Termination of the last AHP is only one event of many in which there is disparity between orbital forcing and atmospheric conditions governing precipitation across northern Africa, but there is at least one potential extrinsic forcing mechanism that differs from previous threshold-crossing events. In this regard, the introduction of domesticated animals must be considered when examining tipping points in the ecology of northern Africa and creation of a novel anthropogenic ecosystem. There is some debate on the timing, but it is generally accepted that domesticated animals, including goats (Capra hircus), sheep (Ovis ovis) and cattle (Bos taurus) arrived in the Fayum Depression from southwestern Asia by 7.35 ka (Figure 3; Linseele et al., 2014) with potentially earlier occurrences in the surrounding circum-Nile River and Western Desert regions dating as far back as 8.1 ka (Barich, 2014; Brass, 2018; Osypinska et al., 2021). Evidence for dairying and intensive wild cereal foraging has been identified from organic residues in ceramics and palaeobotanical remains from ca. 8 ka at the site of Takarkori in SW Libya, spreading eastward thereafter (Dunne et al., 2018; Mercuri et al., 2018). Neolithic technological complexes are dated to 7.4 ka in the Tunisian Maghreb (Mulazzani et al., 2016), on the Messak plateau of southwestern Libya from 8.3 ka (Barich, 1998; Gallinaro and di Lernia, 2018), the Zemmur region in Western Sahara ca. 6.4 ka (Borrell et al., 2018), the El-Khil and Kaf Taht el-Ghar caves of northern Morocco from 7.4 ka (Martínez-Sánchez et al., 2018a, 2018b, 2021). Based on the available evidence, there is a general westward and southward expansion of Neolithic economies from North African points of origin that transpires over the ensuing 2000–3000 years (di Lernia, 2013; Wright, 2017).
During the incipient phases of livestock herding in the Sahara, the direct footprint of pastoral occupation was small because occupations at most sites were ephemeral (Marshall and Hildebrand, 2002). There are indirect forms of evidence such as an increase in the numbers of Steinplätze (fireplaces/hearths) (Gabriel, 1987; Gallinaro, 2018) and population modelling based on gross analysis of radiocarbon ages (Manning and Timpson, 2014) suggesting that the numbers of sites occupied by pastoralists during the terminal phases of the AHP may have been equivalent to those of fisher-hunter-gatherers from the preceding zenith of the AHP. With equivalent numbers of humans living on a landscape, equivalent ecological impacts should be expected. However, two factors separate the early and middle Holocene phases of Saharan occupations: (1) the climate was in a precessionally driven state of aridification due to the southward displacement of the ITCZ, and (2) livestock economies were unprecedented features of this landscape.
BMTC theory argues that there will be recursive effects to the ecology of a region with the introduction of a novel, significant trophic element into a system, which is called ‘turnover’ (Turner and Gardner, 2015). Trophic systems rely on relative stability of population dynamics, but when a new species or selective pressure is introduced, there can be a radical reorganization of the system. Within the context of savannah-forest mosaics, fire produces an open landscape (Scherjon et al., 2015), which are habitats that animals who have evolved within such mosaics would tend to avoid. Grazing animals are more vulnerable to predation in open grasslands. The avoidance of open land as landscapes revegetate following a fire allow floral succession to unfold – primary colonizing mosses yield to short grass, which yields to tall grasses, which yields to savannah-forest mosaic (Hoffmann et al., 2012).
The evidence for fire across the Sahara is found throughout the Holocene (Baumhauer et al., 2004; Marlon et al., 2013) and likely extends deep into the Pleistocene (Hoag and Svenning, 2017). However, charcoal particles counted from an offshore sediment record adjacent to the western Sahara show high (>6 × 103 particles cm−3) levels of charcoal during the AHP, which significantly increases between 7 and 6 ka (10 × 103 particles cm−3) (Dupont and Schefuß, 2018). From 6 to 3 ka, charcoal levels averaged 4 × 103 particles cm−3, which is attributed to an overall reduction in available fuel load for burning (Dupont and Schefuß, 2018). A charcoal record from Lake Tilla in northeastern Nigeria also finds continuous burning throughout the Holocene with only slight variability (Salzmann et al., 2002). Neolithic sites at the Selima Oasis in Sudan (Haynes et al., 1989), Segedim depression in northern Niger (Schulz, 1994) and Ounjougou in southern Mali (Huysecom et al., 2004) likewise have charcoal records indicative of landscape-scale fires.
Due to their long, continuous presence in the region, fires on the Saharan landscape alone could not have induced a threshold-crossing event. The presence of grazers in the region also has a deep evolutionary history. But, ecological turnover is normally catalyzed in multiple ways, and in this instance is argued to have been forced by a combination of an altered landscape of fear in which livestock were encouraged to graze in areas that would have previously been avoided in concert with natural (orbital) climatic changes. Allowing livestock to fodder in a drying or recently burned grassland would have promoted the growth of shrub vegetation at the expense of grass, which reduced vegetation transpiration into the atmosphere and enhanced dust entrainment and albedo. Livestock preferentially feed on grass relative to scrub vegetation (de Leeuw, 1979; McNaughton, 1985), and as they forage across an area, nascent sprouted grass resources will be depleted, which reduces the overall species diversity and leaves shrubs to predominate an area’s floral ecology (Huang et al., 2018). Within the context of a drought, seed germination is retarded in favour of herbaceous sprouters, which further encourages the growth of shrubland at the expense of grasslands (Parra and Moreno, 2018). Such a phenomenon reduces the plant functional diversity of a region, and a qualitative reduction in this metric is the best-fit scenario for present-day climate models that explain the end of the AHP (Groner et al., 2018; Hopcroft and Valdes, 2021).
In locations where proxy data have been collected, there is ample evidence of rapid ecological turnover in concert with the introduction of domesticated livestock in areas that were in the process of undergoing an orbitally-induced transition to more arid conditions. For example, there is a shift from grassy woodlands to shrubland documented at Sai Island, Sudan and adjacent Nile River regions concurrent with increasing presence of livestock grazing during the middle Holocene (Florenzano et al., 2019). The spread of psammophilous shrub vegetation at Wadi Teshuinat in the Libyan Sahara at the expense of grasses is attributed to aridification of the region in combination with selective grazing by herbivores, especially after 4.9 ka (Mercuri, 2008). The arrival of domesticated animals at Ifri Oudadane, Morocco at 7.3 ka correlates to a decrease in arboreal pollen and grasses and an increase in maquis (shrub) vegetation (Morales et al., 2013). At nearby Ifri n’Etsedda, Morocco between 7.4 and 6.8 ka (Linstädter et al., 2016), and Tin-a-Hanakaten, Algeria at 7.2 ka (Aumassip, 1984), similar turnovers to non-arboreal pollen taxa at the expense of arboreal pollen are documented when domesticated animals are first identifiable in the archaeological record. On a broader scale, sediment proxies from the Nile River between 8.7 to 6 ka show abrupt transitions between grassy and shrub-dominant conditions within the context of a slow decline in precipitation (Blanchet et al., 2015; Hennekam et al., 2014). On the other hand, woody savannah biomes such as Mege, in the floodplain environment of Gajiganna in what is now northeastern Nigeria, have undetectable changes in the charcoal composition associated with the introduction of livestock after ca. 3.8 ka and only minor changes with Iron Age settlement after 2.4 ka (Höhn et al., 2021). The persistence of generally wetter biome until ca. 0.95 ka and different land use practices of the Chad Basin compared to surrounding areas show that there is no one-size-fits-all model to landscape formation in relation to early agricultural practices.
While overall numbers of pastoralists were undoubtedly low in the incipient phases of domestication, the knock-on effects of localized threshold crossing events can accumulate to impact a region’s ecology. Zerboni and Nicoll (2019) argue that erosion of desert pavements from trampling since the Neolithic has enhanced aeolian production, which is a general marker of the Anthropocene as expressed in northern Africa. Rapid upticks in dust deposition in marine sediment records across the eastern mid-Atlantic during the Holocene are consistent with a rapid and large-scale retreat in vegetation and lowering of lakes sufficient to produce an aeolian sediment source (Egerer et al., 2017, 2018), which temporally correlates with the introduction of livestock into the region. Anomalous dust flux detected in a 180 ka sediment core in the Canary Islands between 8 and 7 ka is interpreted as a potential footprint of the expansion of pastoral economies into the western Sahara, despite a lack of direct evidence for a significant archaeological presence (von Suchodoletz et al., 2010). A similar uptick in aeolian quartz is also noted in middle to late Holocene ocean sediment cores adjacent to the western Sahara, which is attributed to potential human disturbances associated with the introduction of livestock (Pokras and Mix, 1985). On the site level, there is a significant increase in aeolian activity coeval with the arrival of domesticated animals and decline in arboreal cover at Tin-a-Hanakaten, Algeria (Aumassip, 1984). An accelerated period of erosion in a pedo-sedimentary sequence from the foothills of the Mandara Mountains in northern Cameroon occurs during the Late Holocene, which corresponds to increased archaeological settlement of the region (Diaz et al., 2018). Model simulations indicate that increased dust flux reduced convective (summer monsoon) rainfall during the middle Holocene by approximately 13% in the Sahara and 58.6% in the Sahel (Messori et al., 2019; Thompson et al., 2019). Orbital parameters alone insufficiently explain such anomalies in the long sedimentary records. The spatial discontinuity of the termination event as captured by models (Hopcroft and Valdes, 2021) and the proxy records (Adeleye et al., 2021) support the notion that local disturbance ecologies, potentially associated with human technologies, could have accelerated the tempo and magnitude of some threshold crossing events during the terminal AHP in northern Africa.
Late Holocene human ecologies of Africa
While middle Holocene landscape transformations in arid and semi-arid northern Africa associated with the introduction of domesticated animals represents one potential avenue of anthrome construction, the coeval evolving relationship between humans and plants is another facet of niche construction to explore. Changes to the composition of plant communities within biomes are foundational to inducing broad-scale ecological succession. Climate variability will exert selective pressure on different plant taxa, depending on how adapted to the given environment those species are. However, it is a well-established ecological tenet that trophic systems are in continuous disequilibrium relative to changing climates and balances between prey and predators will fluctuate according to their evolutionary fitness (Davis, 1986). Within the context of anthropogenic ecologies this disequilibrium has presented opportunities for highly social niche constructors like our species to maximize food security. This is because our ability to strategically plan and communicate knowledge intergenerationally has resulted in ecological succession that has, to this point, provided a net benefit to our evolutionary fitness. However, the broader context of niche construction and ecological turnover beyond the scope of our immediate habitats are only now becoming apparent.
Intensive human exploitation of plants in Africa can be traced to the early Holocene (before 10 ka) at sites in the Libyan Sahara (Dunne et al., 2016; Mercuri et al., 2018). During the AHP, northern Africa was populated by ceramic-using forager-fisher-gatherers who practiced high residential mobility in the early phases, but transitioned to ‘delayed return’ modes of subsistence during the terminal phases of AHP (Gatto and Zerboni, 2015; McDonald, 2016; Stojanowski and Knudson, 2014). As the landscape transitioned to more arid conditions, increasingly transhumant forms of subsistence took root in the form of animal pastoralism (see previous section). Traditionally, it has been argued that animal pastoralism preceded the use of domesticated plants (Marshall and Hildebrand, 2002), but recent archaeological investigations demonstrated that intensive uses of plant resources, including wild ancestors of sorghum (Sorghum arundinaceum) by 10 ka the Nile River Valley (Venkateswaran et al., 2019) and domesticated cereals and pulses (Hordeum vulgare, Triticum monococcum/dicoccum, Triticum durum, Triticum aestivum/durum, Lens culinaris and Pisum sativum) appearing by 7–6 ka at Ifri Oudadane, Morocco (Morales et al., 2013), in the Butana Group at the confluence of the Blue and White Nile rivers (Out et al., 2016; Winchell et al., 2017). Forms of pearl millet (Pennisetum glaucum) were domesticated by 4.9 ka in the western Sahara (Burgarella et al., 2018) and spread outward after 4.5 ka (Ozainne et al., 2014), but non-domesticated taxa were utilized by foraging populations for many millennia prior (Kahlheber and Neumann, 2007; Mercuri et al., 2018). The domestication of African rice (Oryza glaberrima) has been traced archaeobotanically to between 10 and 2 ka from the Inland Niger Delta region following a long process in which climate change and selective propagation culminated in the ultimate domestication of this specific species (Cubry et al., 2018). Prior to the ultimate domestication of any plant or animal, the wild ranges of the organisms are widespread and artificial culling and propagation activities are multilocal. Plant foraging creates an opportunity for niche construction in which the harvesting of edible plants resulted in modification of the environment favourable to humans, which reciprocally intensified over time ultimately resulting in domestication of select species in certain locations. However, the impacts of niche construction are not exclusively confined to emergent domesticate loci.
The Iron Age of Africa is associated with a significant transformation of landscapes into which it spread, beginning after 5–4.5 ka in the borderlands region of northern Nigeria and Cameroon where the Bantu language family has its roots and spreading throughout sub-Saharan Africa by 2 ka (Henn et al., 2008; Koile et al., 2022; Prendergast et al., 2019; Russell et al., 2014). In the initial phases, Iron Age farmers engaged in mosaic farming practices with adoption of crops like pearl millet, yams (Dioscorea rotundata) and African rice along with household livestock rearing while continuing to harvest wild or managed forest and aquatic resources (Lane, 2004; Neumann et al., 2012). Until recently, limited evidence of anthropogenic disturbance ca. 4 ka in the rainforests of central Africa and phylogenetic analyses of Bantu languages had indicated that early Iron Age farmers initially avoided rainforest environments, preferring instead savannah ecosystems (Grollemund et al., 2015), however more recent phylogeographic analyses have called this hypothesis into question finding evidence for Bantu language expansion into the central Congo rainforests by 4.4 ka (Koile et al., 2022). The spread of southeast Asian domesticates such as manioc (Manihot esculenta), bananas (Musa sp.), yams (Dioscorea sp.) and taro (Colocasia esculenta) are believed to have originated from the northeast of the continent and spread into tropical climes of Africa between 2.5 and 1.5 ka (Power et al., 2019). The spread of the Iron Age from its homeland in west-central Africa to the east and eventually south took the form of genetic replacement of indigenous hunter-gatherer groups in Malawi (Lipson et al., 2022), whereas post 3 ka admixture between extant pastoralists and hunter-gatherers with late-coming Iron Age farmers in Kenya, Uganda and Botswana call into question the ‘package migration’ model of farming, iron and Bantu languages into southern Africa espoused by mid-to-late 20th century archaeologists (Wang et al., 2020). Instead, farming, land clearance patterns and related technologies are assumed to have spread more syncretically, manifesting themselves in regionally distinct manners (Crowther et al., 2018).
Mixed foraging/farming practices during the Iron Age were retained deep into the Holocene in the northern Congo Basin (Bleasdale et al., 2020), at Kintampo sites in central Ghana (Logan and D’Andrea, 2012) and in the riparian coastal regions of Kenya (Culley et al., 2021; Wright, 2007), while other regions, particularly in southern Africa, experienced a more complete transformation into fully domesticated landscapes (Badenhorst, 2011; Davies et al., 2022). The degree to which iron production is associated with charcoal production and landscape clearance is hotly debated (Bayon et al., 2012; Garcin et al., 2018; contra Giresse et al., 2018), nevertheless either through direct or indirect land management processes, the transformation of most of sub-Saharan Africa into some form of an anthrome before the Industrial Era follows trajectories documented elsewhere on Earth (Ellis et al., 2020, 2021), largely entrenched by 3 ka (Stephens et al., 2019). Although the spatial extent of African lands directly under cultivation by 3 ka has been modelled to have been relatively low (<1%; Cao et al., 2021; Kay and Kaplan, 2015), the knock-on effects of land clearance for food and technological production in prehistory have been estimated as exceeding those of today (Kaplan et al., 2011; Ruddiman and Ellis, 2009).
Niche construction in the African context involved significant amounts of land clearance in order to maximize opportunities for desirable species at the expense of myriad economically non-advantageous plants. However, this form of niche construction had ecological impacts that are identifiable in regional proxy records. For example, a study of δ13C and δ15N from terrestrial leaf waxes from Lake Wandakara, Uganda shows acceleration of deforestation and eutrophication ca. 1 ka attributed to human agricultural land management practices (Russell et al., 2009). Soil induration/ferralitic crust formation and aeolian activation correlate to land clearance across western Africa (Da Costa et al., 2015). Extensively disturbed ferrosolic agricultural landscapes have been measured as having reduced biodiversity (Brussaard et al., 2007; Padonou et al., 2015). Long sediment core records consistently attribute upticks in erosion and sedimentation particularly with Iron Age land clearance for cultivation (Bayon et al., 2012; Garcin et al., 2018; Neumann et al., 2010). These correspond to terrestrial records, such as in the Gourits River Basin in Western Cape Province of South Africa, where initial settlement of herders after 1.6 ka degraded the natural vegetation cover and correlates to the initiation of a period of sediment erosion, followed by a more substantial period of erosion following European settlement of the region beginning 0.4 ka (Damm and Hagedorn, 2010). Furthermore, on a global scale, agricultural farmlands are net contributors of atmospheric carbon as they reduce the carbon pool, which has added to the rise of temperatures associated with global warming (Lal, 2001).
Research conducted in the middle Benoué River Valley of northern Cameroon provides a longitudinal view of landscape succession concurrent with the construction of settlement mounds. There are a total of 39 documented settlement mounds north of the present-day city of Garoua (Wright et al., 2017). Stable isotopic reconstruction of the landscape around the site of Langui-Tschéboua demonstrates a significant reduction in the prevalence of woody (C3) vegetation before and after mound construction (Figure 6; Wright et al., 2019). There are also contrasting ecological impacts between highland and lowland contexts with the isotopic ecology of the former region retaining more tree cover following human settlement contemporaneous to lowland contexts (Figure 7; Wright et al., 2019).
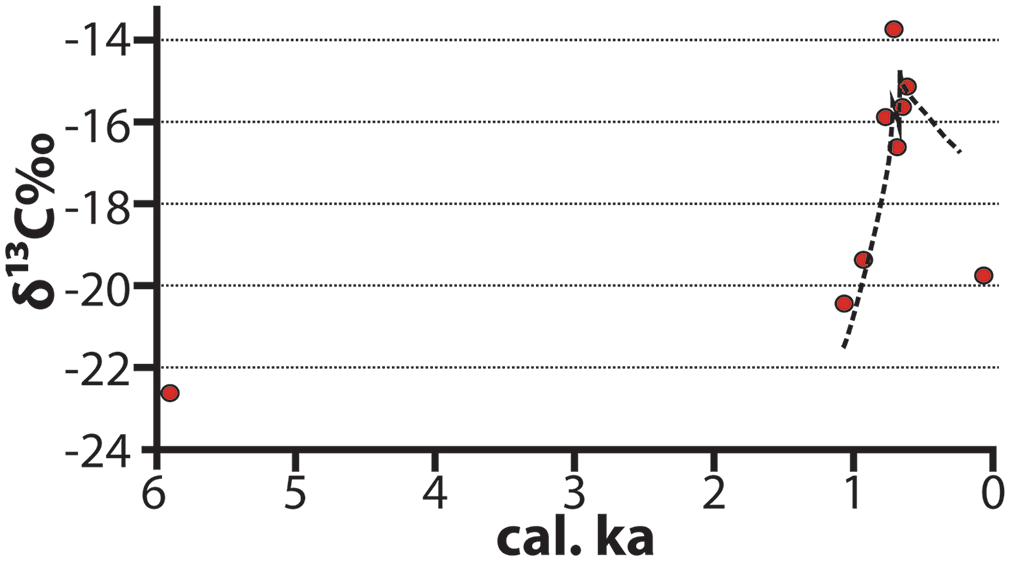
δ13C isotopic profile of Langui-Tschéboua settlement mound, Benoué River Valley (Wright et al., 2019). The regression line is an exponential fit of values from the occupation levels of the mound. The earlier proxy (5.9 ka) is from the underlying alluvial substrate.
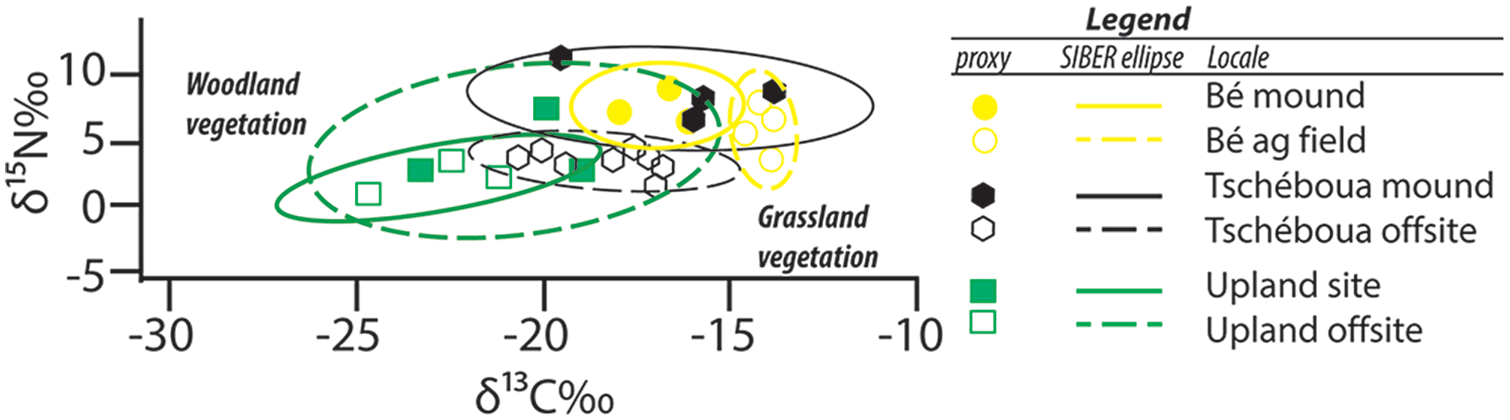
δ15N‰ (vs. AIR) and δ13C‰ (vs. VPDB) for samples collected in onsite (filled shapes) and offsite (open shapes) samples for Bé (yellow circles), Langui-Tschéboua (black triangles) and GRA-1 (green squares) (Wright et al., 2019). Standard ellipses generated in the SIBER package in R (Jackson et al., 2011) at the 95% (2-σ) confidence interval.
Across western Africa, the identification of anthropogenic parklands resulting from clearance of woody vegetation occurs in a variety of ecological contexts and can be particularly difficult to identify in riparian settings particularly when modelled with natural climate variability (Heinrich and Moldenhauer, 2002; Höhn et al., 2021). Early Iron Age sites from northern Burkina Faso show widespread cultivation interspersed with long intervals of fallowing from ca. 2 to 1 ka after which shorter fallow periods and more intensive livestock production as well as decreasing precipitation resulted in the creation of a more open (grassy) landscape (Höhn and Neumann, 2012). A landscape succession study on the Laikipia Plateau of central Kenya similarly shows a transition from a closed forest ecosystem at 2.3 ka to an open woodland/grassland environment by 1.6 ka (Muiruri et al., 2022) also within a generally drying climate regime (Taylor et al., 2005). Maintenance of an open landscape using fire and grazing is attested by charcoal and fungal spores after 0.9 ka (Muiruri et al., 2022) and conforms with recent studies on rangeland/parkland creation and mechanisms of perpetuation (Phelps et al., 2020; Western et al., 2021). The alteration of habitat, either through direct forms of land clearance, coppicing and husbandry or as a result of indirect ecologies of fear from increasing sedentism on mounds or culturally bounded rangelands, has profoundly transformed the landscape of virtually all of sub-Saharan Africa, beginning ca. 5–4.5 ka, accelerating after 2–1 ka to the point where current ecosystem functionality has been utterly dependent on humans for at least the last millennium.
Farming is not intrinsically deleterious to a flourishing ecological system – adverse impacts to biodiversity are contextual and circumstantial (Muthee et al., 2022). For example, a 7-year study of shifting cultivation plots in Ivory Coast found profound short-term reductions in microbial communities during periods of cultivation, but once fallowed, bacterial flora in ferrolitic soils recovered to pre-disturbance levels within the cycle of a single rainy season (Persiani et al., 1998). A similar comparative study of arthropod diversity in southern Benin found high occurrences of species richness on formerly cultivated (but presently forested) plots (Lachat et al., 2006). A broad survey of coincident non-human vertebrates and human populations show strong positive correlations between archaeological sites and endemic species richness (Fjeldså and Burgess, 2008). Studies of regions that receive high amounts of precipitation consistently demonstrate that while deforestation and soil erosion associated with land clearance and agriculture diminish biodiversity, flora and fauna inhabiting these regions are resilient provided there are sanctuaries for refugee populations.
Furthermore, many anthropogenic activities related to agriculture have been documented as enhancing ecosystem biodiversity and resilience. For example, studies from the Central Rift Valley of Kenya (Marshall et al., 2018) and Tanzania (Storozum et al., 2021) find long-term augmentation of fertility within cattle enclosures compared to non-enclosure areas from Pastoral Neolithic contexts lasting thousands of years beginning after ca. 4 ka. Studies across western Africa consistently find soil ‘ripening’ within the context of abandoned villages and settlement mounds (a.k.a. tombondu), which have been compared to the formation of terra preta dark earth soils of the Amazon Basin (Fairhead and Leach, 2009). Pastoralists are critical for the redistribution of nitrogen and phosphorus around the landscapes of eastern Africa and the soil and plant communities of the region have come to depend on them (Augustine, 2003).
Traditional human land management practices can be important components of maintaining and augmenting biodiversity. The promotion of agroforestry such as honey harvesting, provisioning of tree byproducts such as fruits or nuts and carbon sequestration services have significantly increased landscape resilience in the places where they are practiced (Muthee et al., 2022). A comparative study of managed and unmanaged agricultural lands in Burkina Faso demonstrates that farmers were crucial agents in conserving cornerstone plants for ecosystem functioning (Nacoulma et al., 2011). Areas of former human settlement within the Gola Forest, Sierra Leone have higher densities of extent plant and bird species compared to areas that have no evidence of former occupation (Kandeh and Richards, 2011). Herders such as the Maasai of eastern Africa are critical agents in conservation of predatory cats due to the deep cultural connections they have with them (Goldman et al., 2010). Domestication of wild plant species and arboriculture by farmers in the Usambara Mountains, Tanzania have been documented as a means of conserving local plant diversity, which has recursive effects enhancing ex-situ microbial and faunal biodiversity (Msuya et al., 2008; Munishi et al., 2008). In prehistoric Fayum, limited, selective coppicing of tamarisk (Tamarix sp.) yielded only minimal disturbances to overall species abundance of different woodland communities (Marston et al., 2017), similar to the effects reported in the Chad Basin discussed above (Höhn et al., 2021).
Conclusion
Agriculture and its associated human settlement practices have transformed the African landscape on an unprecedented scale in the geologic record, but it did not emerge out of nowhere. If the Anthropocene is marked by the supplanting of natural geologic formation processes by artificially (human-) driven processes, there will be no Golden Spike to pound to mark its onset in Africa. Historical ecologists recognize that anthropogenic landscapes are the result of long and cumulative processes of human intervention in the life cycle of plants and animals (Armstrong et al., 2017) and as the continent that has the longest human settlement history, Africa hosts the oldest anthropogenic landscapes. Over time, the process of landscape domestication has shifted from selective culling of non-consumable plants via coppicing, burning and weeding and propagation of consumables in the early Holocene to active protection of productive species (e.g. goats, patches of fertile soil in which favourable plants germinate) in the middle to late Holocene to one in which there is wholesale clearance of flora and fauna for industrial scale agriculture in the modern era.
Fundamentally, whether agriculture enhances or diminishes overall biodiversity depends on the scale of land clearance, complexity of the ambient ecological composition of the landscape and the degree to which farmers consciously and actively promote conservation of endemic plant and animal species (Bhagwat et al., 2008; Brussaard et al., 2007; Tscharntke et al., 2005). In some cases, human intervention into landscape processes result in a net gain for biodiversity, but there are many instances in which biodiversity incurs a resulting net loss. In other words, conscious self-removal of humans as the centrepieces of a μ-driven (fear-based) landscape has proven to be critical for preserving elements of the natural world needed for robust ecosystem functionality.
Ecological systems that have undergone systematic co-evolutionary adaptation to human activity, which presently includes the vast majority of the earth’s surface and oceans, are now utterly dependent on humans to sustain them in their present formulations. Just as pre-agricultural forms of anthropogenesis related to burning and coppicing altered BMTCs via novel recasting of landscapes’ ecologies of fear, plant cultivation reorganized relationships between microbial and plant and animal communities. These impacts are not limited to archaeological sites – they cascade across biogeographic regions and ecospheres (biosphere, hydrosphere, lithosphere, atmosphere). These processes are ongoing and confound precise ordination of an Anthropocene epoch using ecological biomarkers because they occur fluidly across time and space.
Footnotes
Acknowledgements
I have had the distinct privilege of having hundreds of conversations about this topic with innumerable friends, colleagues and students over the years, and I have learned immensely from each and every one. In particular, I have gained the most from my conversations with farmers and pastoralists in various parts of Africa whose deep knowledge of the land they live on has inspired me to dig deeper and deeper to understand it even a fraction as well as they do. I thank three anonymous reviewers of this paper, editor Robert Costanza and commenters who have looked at fragments or draft versions over the years.
Funding
The author received no financial support for the research, authorship, and/or publication of this article.