
Abstract
Scholars have argued that plant domestication in eastern North America involved human interactions with floodplain weeds in woodlands that had few other early successional environments. Archeological evidence for plant domestication in this region occurs along the Mississippi river and major tributaries such as the Tennessee, Ohio, Illinois, Missouri, and Arkansas rivers. But this region is also known as the prairie peninsula: a prairie-woodland mosaic that was maintained by anthropogenic fire starting as early as 6000 BP. Contrary to conventional wisdom, recent research has shown that bison were also present in the prairie peninsula throughout the Holocene. Recent reintroductions of bison to tallgrass prairies have allowed ecologists to study the effects of their grazing on this ecosystem for the first time. Like rivers and humans, bison create early successional habitats for annual forbs and grasses, including the progenitors of eastern North American crops, within tallgrass prairies. Our fieldwork has shown that crop progenitors are conspicuous members of plant communities along bison trails and in wallows. We argue that ancient foragers encountered dense, easily harvestable stands of crop progenitors as they moved along bison trails, and that the ecosystems created by bison and anthropogenic fire served as a template for the later agroecosystem of this region. Without denying the importance of human-river interactions highlighted by previous researchers, we suggest that prairies have been ignored as possible loci for domestication, largely because the disturbed, biodiverse tallgrass prairies created by bison have only been recreated in the past three decades after a century of extinction.
Introduction
Agriculture grew out of multispecies relationships that transformed ecosystems, but were also shaped by these systems’ previous configurations. The emergence of an agricultural system is one of self-transformation. Following Ingold (2000), we can use the weaving of a basket as a metaphor to understand this process: “the equable form of the spiral basket does not follow the dictates of any design; it is not imposed upon the material but arises through the work itself. Indeed, the developing form acts as its own template” (p. 345, emphasis added). The bodies and behaviors of domesticated plants and animals arise from their deep evolutionary ecology. In order to understand how they came to be domesticated participants in agricultural economies, we must first understand how the earlier landscapes they inhabited served as templates for the later agroecosystems.
Many of our most important crops are descended from annual plants that produce an abundance of small, hard seeds. Crops such as the millets (Setaria italica, Panicum miliaceum, Eleusine coracana, Pennisetum glaucum, Echinochloa spp., Digitaria spp.), sorghum (Sorghum bicolor), amaranth (Amaranthus spp.), quinoa (Chenopodium quinoa), buckwheat (Fagopyrum esculentum), and the lost crops of eastern North America (Chenopodium berlandieri, Hordeum pusillum, Iva annua, Phalaris caroliniana, and Polygonum erectum) all have small seeds with thick, indigestible fruit or seed coats in their wild forms. Anthropologists have struggled to understand why ancient foragers chose to harvest plants that seemingly offer such a low return on labor. Before any mutualistic relationship could begin, people had to encounter stands of seed-bearing annual plants dense and homogenous enough to spark the idea of harvesting seed for food. Large stands reduce the energetic costs of traveling between stands or searching for additional populations, thus leading to higher harvesting efficiency than gathering from many small stands. Today, it is rare to find the wild progenitors of small-seeded crops in large, dense stands outside of urban, pastoral, or agricultural settings. The progenitors of some of North America’s small-seeded crops can also be found in large, dense stands in floodplains (Smith, 1992). Given their affinity for anthropogenic environments that did not exist until the Late Holocene, what kinds of ecosystems did these species evolve to prosper in? In what settings would early foragers have encountered them? We have argued elsewhere that many of the progenitors of small-seeded crops co-evolved with grazing animals and have a suite of adaptations that allow their seeds to be effectively dispersed in dung (Spengler and Mueller, 2019). This is why dense, homogenous stands of crop progenitors can often be found in herd animal corrals, wallows, and rangelands.
In addition to the high labor costs of harvesting small-seeded annuals occurring in small scattered patches, it is also difficult to thresh, winnow, and hull them, processes which are necessary to remove hard seed and fruit coats or chaff and render the grains digestible by humans (Gremillion, 2004). Beyond the mystery of how and why foragers were attracted to small, hard seeds as food lies another problem: how did people first start saving seed and planting? The wild progenitors of many temperate zone crops produce seeds that are dormant without some treatment that mimics conditions in the soil seed bank, including all but one of eastern North American’s lost crops (Baskin and Baskin, 2014; Mueller et al., 2019). If they are harvested and stored in the manner of domesticated seed stock, as little as ten percent of the crop will germinate when planted, an obstacle that early cultivators must have overcome (Abbo et al., 2011; Ladizinsky, 1987). The many difficulties inherent in the use of wild annuals as seed crops have led to decades of speculation that such plants must have been domesticated in an atmosphere of scarcity, and that ancient people would not have chosen to harvest or cultivate such plants unless their other choices were limited (Flannery, 1969; Weiss et al., 2004; Zeder, 2012).
In eastern North America, Indigenous people created a biodiverse domesticated landscape during the mid-Holocene (Smith and Yarnell, 2009). This landscape included agriculture without flood management or irrigation along the Mississippi river and its tributaries, a land management choice that preserved the rich aquatic animal and plant foods of the floodplains. Ancient Indigenous people also extensively burned, which decreased the density of forests and increased the prevalence of fire-loving species. These changes in forest structure and composition increased the abundance of acorns (Quercus spp.), hickory nuts (Carya spp.), chestnuts (Castanea dentata), hazelnuts (Corylus americana), and wild fruits, as well as game, by creating edges and clearings (Delcourt and Delcourt, 2004; Nowacki and Abrams, 2008). Communities in the midcontinent also domesticated at least six annual seed crops, including sunflowers (Helianthus annuus), squashes (Cucurbita pepo var. ovifera) and four others that are now extinct (hereafter “lost crops,” Table 1) While there is no good shorthand geographical term for the region where this agroecosystem emerged, it was centered on the Mississippi river between northern Illinois and Iowa and southern Arkansas and Tennessee, encompassing most of the vast Mississippi watershed between the Appalachian mountains and the eastern Great Plains (Figure 1). Hereafter, we will refer to this region as the midcontinent.
The eastern agricultural complex.
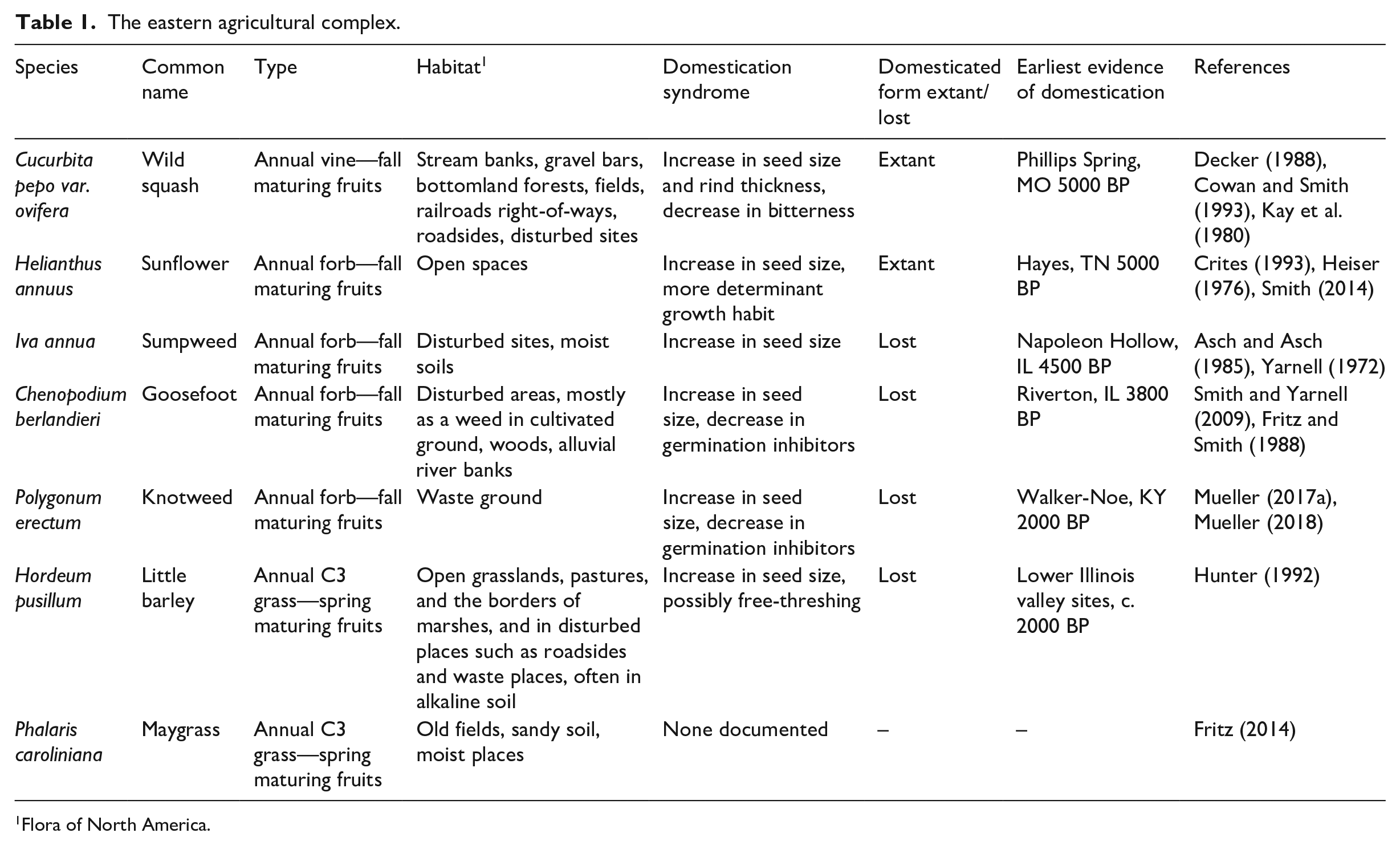
1Flora of North America.
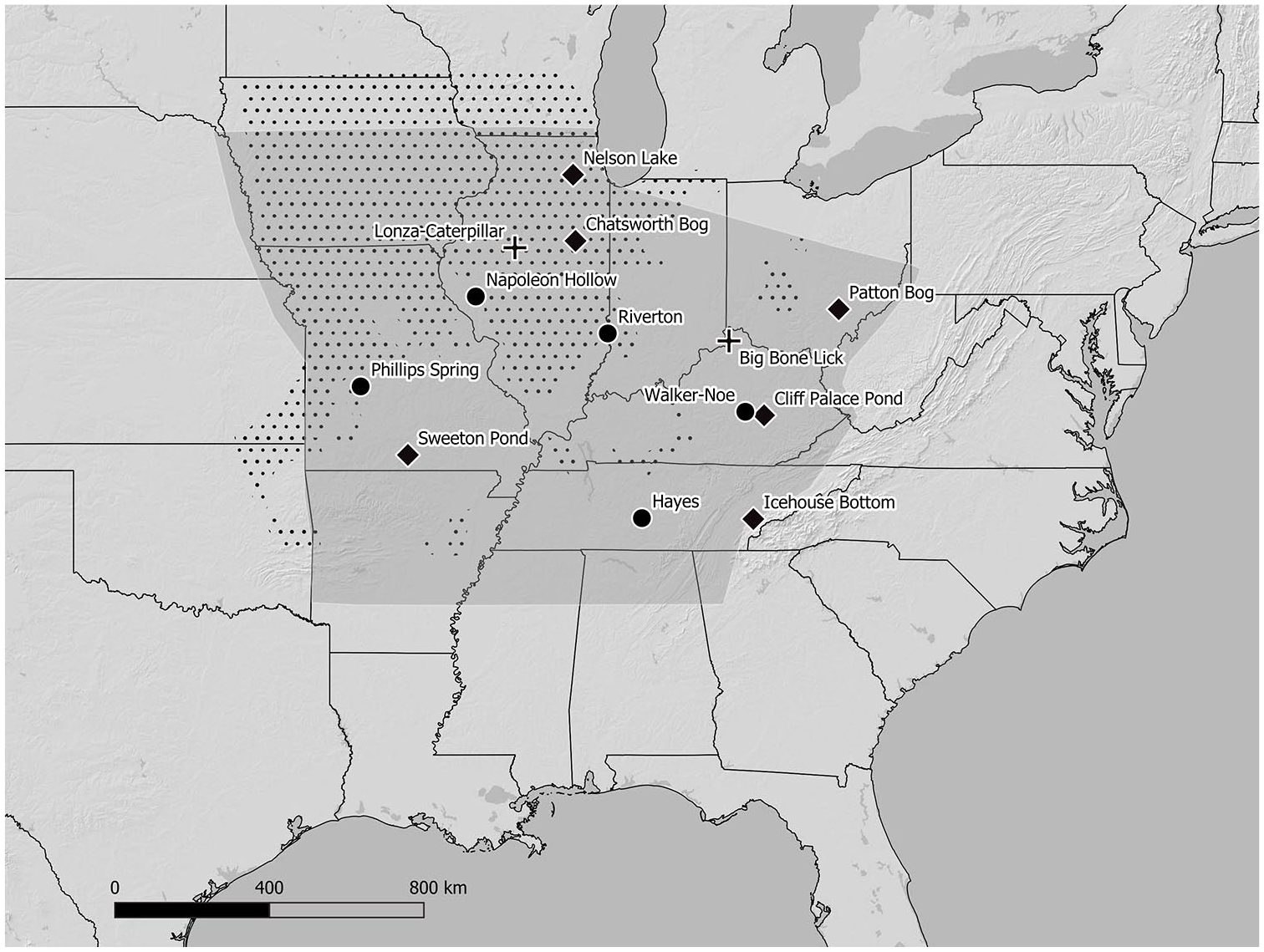
Map of eastern North America. The dotted area is the extent of tallgrass prairie documented by Euro-American settlers, called the prairie peninsula, adapted from Transeau’s (1935) map. The shaded area is the core area where evidence for the cultivation of the eastern agricultural complex (Table 1) is documented in the archeological record. Circles represent the archeological sites with the earliest evidence for plant domestication: Phillips Spring (squashes), Napoleon Hollow (sumpweed), Riverton (goosefoot), Hayes (sunflower), and Walker-Noe (knotweed). See Table 1 for references. Diamonds represent paleo-environmental sites with evidence for anthropogenic fire that are discussed in the text. Crosses are the Holocene bison kill sites east of the Mississippi river that are mentioned in the text.
Compared to other regions, it is especially challenging to understand how and why ancient foragers and early farmers might have shaped ecosystems, because the domesticated subspecies of most of their crops are extinct, and because the landscape where they were domesticated has been transformed by colonial invasion, the genocide or removal of Indigenous people, the clear-cutting of the forests, and the conversion of the prairies to agricultural fields in the past 400 years. There are many different kinds of anthropogenic openings and early successional ecosystems in this region today, including dirt roads and roadsides, agricultural fields, yards and parks, railroad tracks, and pastures for introduced domesticated animals, among many others. These are the kinds of habitats where the progenitors of the lost crops are primarily found (Asch and Asch, 1985, 1978; Cowan, 1978). Most field guides and botanical keys describe them as weedy and well-adapted to human-created early successional ecosystems or “disturbance” (Table 1). This begs the questions: where did they occur before humans modified the ecosystems of eastern North America? Where would late Pleistocene and early Holocene foragers have encountered such weedy plants?
The only non-anthropogenic environments where large stands of lost crop progenitors have previously been documented is within floodplains. In particular, goosefoot (Chenopodium berlandieri), sumpweed (Iva annua), and erect knotweed (Polygonum erectum), as well as wild squashes, are often found on newly deposited sediment close to creeks, rivers, and lakes that flood seasonally. This led scholars working in the Mississippi and Illinois river valleys (Smith, 1992; Struever and Vickery, 1973) to suggest that the earliest cultivation of these species took place on floodplains, where naturally occurring stands provided a dense source of food with almost no labor investment, a dynamic that had previously been suggested to explain domestication in other regions (Sauer, 1952). Rindos (1984) also pointed out that river banks are seasonally disturbed, and therefore, competitive vegetation in suppressed, allowing feral crops or wild progenitors to survive in these disturbed ecosystems longer than they would on a landscape without regular disturbance. He also recognized the utility of floodwaters for seed dispersal purposes.
The widely accepted Floodplain Weed Theory of domestication in eastern North America posits that annual floods created organically enriched clearings and deposited the seeds of water-dispersed species, and that the earliest farmers relied on this yearly inundation to suppress succession (Smith, 1992). Mueller (2017b) and Gremillion et al. (2008) have further suggested that domesticated plant populations became isolated from their wild relatives when people removed these floodplain species from the risky and unpredictable environments in which they had evolved and began to cultivate them in upland clearings created using fire. Previous domestication theorists have focused on the relationship between the lost crops and floodplains because water is the most obvious force of disturbance within the area that archaeologists refer to as the Eastern Woodlands, and thus floodplains provided habitat for crop wild progenitors before widespread human disturbance began. The archeological record suggests that riverine animals were also at the center of pre-Columbian Indigenous subsistence (Styles and Klippel, 1996). In addition to providing food, rivers must have formed important routes of travel, all but ensuring that humans would have repeatedly encountered any plants and animals that lived along them.
Without denying the probable role of floodplains in the domestication of the lost crops, here we will argue that archaeologists have long overlooked another source of disturbance that shaped eastern North American landscapes alongside humans for thousands of years: bison. In discussions of subsistence, they have also often ignored the fact that large swaths of the “Eastern Woodlands” were covered in a prairie-savannah-forest mosaic throughout the Holocene (Figure 1). In recent years, bison have been reintroduced to tallgrass prairie remnants for the first time in over a century. We synthesize studies on the effects of these reintroductions, paleoenvironmental data, and our own observations and to show how human interactions with bison and fire could have given rise to the unique agricultural system that arose in the midcontinent (Figure 1).
A pyropyrrhic victory: the Prairie Peninsula
In 1935, Edgar Transeau (1935) compiled species occurrence data from dozens of botanists to create the iconic map of the “prairie peninsula” of eastern North America (Figure 1). This extension of grasslands into the humid midcontinent had been documented by Euro-American settlers, and went on to puzzle botanists long before ecology emerged as a discipline: “To some it has seemed to be not really prairie but rather deciduous forest with trees absent because of ecological conditions long since terminated . . .to others the Prairie Peninsula represents a pyropyrrhic victory of the Indians attempting to enlarge native pastures, or to capture their grazing meat supply” (p. 423). We can only assume that Transeau saw the anthropogenic prairies as a Pyrrhic victory because they have been so thoroughly erased by Euro-American settlers. Tallgrass prairies are now some of the most endangered ecosystems on earth, and bison were nearly driven to extinction and erased from the midcontinent for hundreds of years (Anderson, 2006). Despite this catastrophic change obscuring our view of the prairie peninsula, today multiple lines of evidence can be marshalled to support the “pyropyrrhic victory” hypothesis. The anthropogenic nature of the now-vanished prairie peninsula and the chronology of prairie retreat and advance over the course of the Holocene is coming into focus.
The prairies of the Great Plains and midcontinent are young ecosystems. This region was covered in glaciers or forests until the early Holocene, when prairies and savannahs expanded from the Rocky Mountains to the Mississippi River. During the dry mid-Holocene Hypsithermal c. 8000-5000 cal BP, grasslands expanded east of the Mississippi river, as the prairie peninsula grew to its full extent (Anderson, 2006). Mesic forests were replaced by fire and drought tolerant oak-hickory forests further east and south (Delcourt and Delcourt, 2004). After the Hypsithermal, as the climate became cooler and more humid, forests should have replaced most prairies east of the Great Plains, as occurred in the upper Midwest (Williams et al., 2009). However, further south, in the midcontinental region where the first domesticated plants were about to emerge, prairies persisted despite climate change in the Late Holocene. At the same time, oak-hickory forests should also have declined, but instead remained dominant in eastern North America until fire suppression began in the 19th century (Delcourt and Delcourt, 2004; Nowacki and Abrams, 2008).
Figure 1 shows the region where the prairie peninsula and the archeological evidence for plant domestication overlap. In the western part of the region, prairies in western Arkansas and northern Missouri were home to both people and bison throughout the Holocene (Cleland, 1965; McMillan, 2012). Even in the Ozark and Ouachita mountains of southern Missouri and northern Arkansas, which were largely forested when Euro-American colonists arrived, anthropogenic burning created landscape heterogeneity and patches of prairie grading into oak savannahs and forests throughout the Holocene (Jones et al., 2017). Recent work at Sweeton Pond in southern Missouri has provided a fine-grained paleoecology of the last 2000 years in the Ozarks, documenting a period of heightened charcoal accumulation from c. 2000 to 1500 cal BP, followed by the establishment local grasslands that persisted until the massive disruptions of the 1400–1500s. With the historically documented arrival (or return) of the Osage in the 1500s, burning and grassland expansion was renewed (Nanavati and Grimm, 2020). Along with similar evidence for grassland formation c. 2500 years ago from Patton Bog in the Hocking Hills of eastern Ohio (Abrams et al., 2014), this sequence provides support for the assertion that even in the most wooded parts of the “Eastern Woodlands,” the Ozark and Appalachian mountains, prairie ecosystems were integral parts of the landscape (Figure 1).
The prairies of central Illinois are especially important to our argument because some of the earliest and best evidence for plant domestication comes from the fringes of what Euro-American settlers called the Grand Prairie, in central Illinois. This vast prairie was flanked by the Wabash and Illinois river valleys, where the earliest domesticated sumpweed and goosefoot have been recovered, and where the best evidence for little barley (Hordeum pusillum) domestication occurs (Asch and Asch, 1985; Hunter, 1992; Smith and Yarnell, 2009) (Figure 1). In the colonial era, the Illinois people who lived on the Grand Prairie used fire to actively maintain a patchy landscape of forest, prairie, and agricultural land, all of which were necessary for their lifestyle (Morrissey, 2019). Historically, bison hunting was also an important part of this lifestyle, in Illinois and elsewhere in the midcontinent (Morrissey, 2016). Yet archaeologists long believed that bison did not cross the Mississippi River until after c. 600 BP (McMillan, 2006; Tankersley, 1987). This belief stemmed from the relative paucity of bison bones in older archeological sites and lack of evidence for bison hunting in areas that were prairies at the time of Euro-American colonization. However, McMillan (2006) compiled a database of archeological and palaeontological occurrences of bison bones and radiocarbon dates to show that bison were present in Illinois throughout the Holocene. Harn and Martin (2006) documented a bison kill site along the Illinois River dating to the Early Woodland period, c. 2500 BP (Lonza-Caterpiller site, Figure 1). Of this rare eastern kill site, they write “Beyond meat, humans seem to have taken little from the carcasses (2006: 53),” a practice that may have been common at kill sites, which perhaps explains why so few bison bones are found at habitation sites (Metcalfe and Barlow, 1992).
The retreat and advance of the Grand Prairie in Illinois has been impressively documented using multiple proxies (Nelson et al., 2006). The authors conclude, “fire may have exerted an important control on the establishment and maintenance of prairie communities in the eastern Prairie Peninsula. . .fire activity became elevated ~500–1000 years prior to the full establishment of prairie, ~6200 BP. Because this change in fire importance did not coincide with any major shifts in our aridity and vegetational records, it is possible that increased fire ignitions resulted in more burns. . .Fires may also have been important for the maintenance of prairies. For example, a marked increase in effective moisture occurred ~5800–5400 BP, as evidenced in change in all proxy indicators at both our sites. This change, however, did not cause woody taxa to expand and prairie taxa to decline. . .” (Nelson et al., 2006: 2534). Studies of Illinois “barrens” (tallgrass prairies along the Illinois river) have ruled out soil as a limiting factor that would have prevented woody vegetation from encroaching on prairies (Kilburn et al., 2009). If neither aridity nor poor soils can explain the presence of prairies in Illinois throughout the Holocene, then anthropogenic fire is the only remaining explanation for their presence, and is also consistent with the ethnographic and historical records of Indigenous communities from this region. In this feat of ecosystem engineering, people and bison were natural partners.
The impact of bison on tallgrass prairies
It has long been recognized that Indigenous people maintained the open canopy oak-hickory forests that were sources of nuts, fruits, and game using fire and other techniques (Abrams and Nowacki, 2008). Increasingly, ecologists also accept that the eastern prairies of the late Holocene were deliberately maintained by people (Anderson, 2006), but current archeological data and theories make it hard to understand why ancient people managed prairies. Bison, the most obvious prairie resource, are not well-represented in the archeological record. Even with the evidence from kill sites that bison were largely butchered in place, the paucity of bones in habitation sites in comparison to later sites and sites further west suggests that bison were not a staple food east of the Great Plains until ~500 years ago (McMillan, 2006, 2012; Tankersley, 1987). Zooarcheological assemblages in this region are instead dominated by forest and river-dwelling animals. Tree nuts were staple foods throughout the Holocene, and the prevailing theory about the beginnings of agriculture is that people took advantage of yearly flooding to cultivate annual plants on naturally disturbed riverbanks. These data and theories suggest that forests and rivers were the foundation of ancient subsistence, and are not compatible with the emerging body of evidence that Indigenous communities intentionally sustained tallgrass prairies on a regional scale, in defiance of climate change and ecological succession.
This disconnect is the result of the centuries long absence of the bison-tallgrass prairie ecosystem. Until the 1990s, no ecologist had seen the biodiverse and heterogenous tallgrass prairies created by grazing bison herds—they had disappeared before the discipline of ecology came into existence. They were also unknown to the generation of botanists and archaeologists who discovered the lost crops of eastern North America. The lost crops are annual forbs and cool season C3 grasses. They are not commonly found in the homogenous prairie remnants that were available for study in the mid-20th century, which are characteristically dominated by perennial warm season C4 grasses that offer little in the way of human food. The following three observations of changes to tallgrass prairies after bison reintroductions led us to initiate fieldwork investigating a potential connection between the domestication of the lost crops and bison:
(1) Reintroduction of bison to tallgrass prairies results in greater species diversity, and in particular, more forbs and annual grasses on a landscape that is otherwise dominated by perennial grasses (Coppedge et al., 1998a). This is primarily because, to a greater extent than other ruminants, bison overwhelmingly eat grasses, and avoid forbs (Coppedge et al., 1998b; Knapp et al., 1999). Knapp et al. (1999) have documented an interesting grazing pattern among the bison of the Konza Prairie. These bison preferentially re-graze the same areas, preferring the younger shoots of grasses and preventing the tallgrasses from fully rebounding: “Thus, within a bison grazing area, forbs are often conspicuously left ungrazed and are surrounded by grazed grasses. . .” (p. 41).
(2) While opening up habitat for annuals, especially forbs, by intensely grazing perennial grass shoots, bison also disperse the seeds of many plants, including some of the lost crop progenitors (Rosas et al., 2008). The lost crops sumpweed and little barley were among the species documented in the dung of free ranging bison in eastern Oklahoma (Rosas et al., 2008). These taxa are even more frequently dispersed by epizoochory (adhesion to fur) than by endozoochory (dispersal in dung). Given the propensity of bison to wallow, these species benefit from being dropped in nitrogen enriched and relatively well-watered clearings using either dispersal mechanism (Coppedge and Shaw, 2000).
(3) The interaction between burning and grazing is also relevant in the context of millennia of anthropogenic burning in eastern North America discussed above. Fulendorf and Engle showed that patchy fires create “grazing focal points” within the prairie, and that bison spend the majority of their time grazing these recently burned areas (Fuhlendorf and Engle, 2001). Bison prefer to create new wallows in these areas, as well (Coppedge and Shaw, 2000). This behavior may increase the probability of dispersing seeds to recently burned locations. Bison dung burns hotter than woody fuels (Crockett and Engle, 1999), creating a potential for a feedback loop where burning leads to intensive grazing, and dung concentrations create good conditions for future burns. Burning may thus have both attracted extremely valuable prey (bison) and created niches for annual seedbearing plants, from the perspective of ancient foragers.
Harvesting the prairie
Taking into account both the paleoenvironmental data and ecological studies of bison reintroductions to tallgrass prairies, we hypothesize that:
(1) Bison would have created patches of habitat for annual forbs and cool season grasses in ancient tallgrass prairies;
(2) Like rivers, bison trails (hereafter, traces) would have provided an easy way for people to travel, and that traveling along a trace would lead to repeated encounters with stands of crop progenitors, and;
(3) It would be possible to carry out efficient harvests of crop progenitor seeds in stands created by bison grazing and wallowing.
To investigate these hypotheses, we conducted fieldwork in the Joseph H. Williams Tallgrass Prairie Preserve (TGPP), a 39,605 acre prairie in the Flint Hills of eastern Oklahoma grazed by 2500 bison and managed by the Nature Conservancy using a patch-burn approach. We chose TGPP for preliminary exploration of our hypotheses because previous research has shown that bison on this preserve dispersed the seeds of two of the lost crops, little barley and sumpweed (Rosas et al., 2008), and that a third, maygrass (Phalaris caroliniana) is also a part of the flora. Unfortunately, two of the lost crops (erect knotweed and goosefoot) do not occur in the TGPP, although close relatives do (Palmer, 2007). For this reason, our discussion here will focus on little barley and maygrass, which set seed in May and June, and sumpweed, which sets seed in October.
We visited TGPP 28–31 May, 18–20 June, 29–30 Sep, and 18–20 Oct 2019. These visits were timed to identify populations of spring and fall maturing lost crops, collect seed from them, and collect bison dung samples during the window in which they might contain crop progenitor seeds. During our first visit we chose three traces to walk (Figure 2) based on coarse grained occurrence data for our species of interest provided by Mike Palmer. We also established a fourth survey area just inside the bison exclusion area and directly adjacent to one of our traces, in order to ascertain how crop progenitor distribution differed in the absence of grazing and trampling (Figures 2, 3b, and 4a).
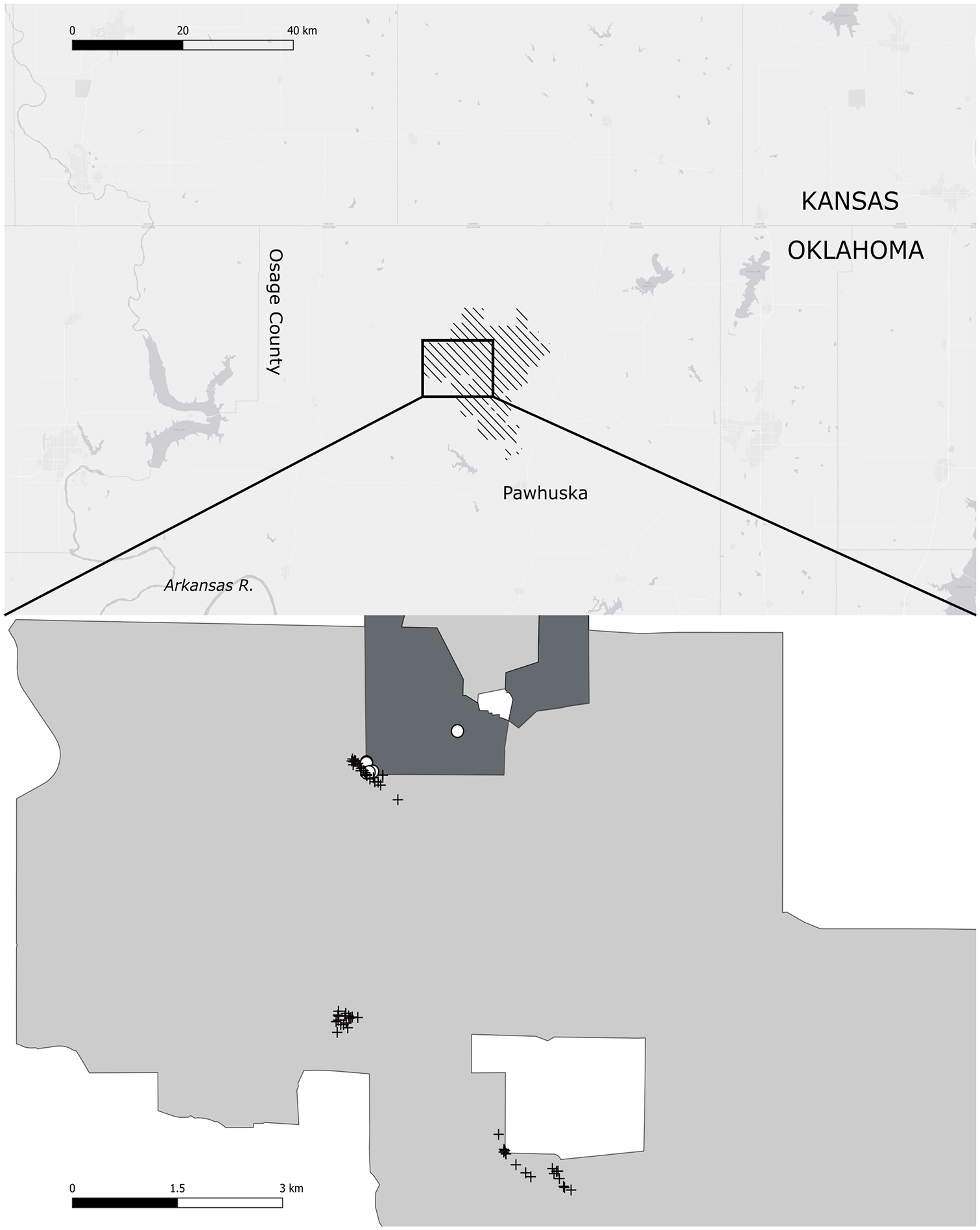
Top: Map showing the location of the Joseph H. Williams Tallgrass Prairie Preserve, in Osage County, Oklahoma. Bottom: close up of the TGPP. Dark grey area is the bison exclusion area, and light grey area is grazed by 2500 bison. Crosses show the locations of dung samples taken at each of our three traces, and white circles show our comparative sites within the ungrazed area.

Observations from spring visits to TGPP. (a) Little barley seeds in a trace. (b) View from inside the bison-grazed area towards the bison exclusion fence. A large wallow full of little barley ends at the fence line. Sumpweed seedlings can be seen in the foreground. (c) A wallow complex with extensive populations of little barely, maygrass, and sumpweed. (d) Bison fur adhering to branches among a dense population of little barley.

Observations from fall visits to TGPP. (a) View along bison exclusion fence, with dense population of sumpweed in the grazed area (right), and sumpweed absent in the ungrazed area (left). (b) Dense popultions of sumpweed on either side of a trace. (c) Harvesting in a large sumpweed population within a wallow. (d) Wild sunflowers and sumpweed in a wallow.
Our observations strongly supported our hypotheses with respect to the cool season annual grasses, little barley and maygrass. Despite the fact that we have never seen little barley described as a common prairie grass, it was the most conspicuous grass along traces and in wallows during both of our spring visits. Large wallows sometimes had the appearance of cultivated fields of little barley (Figure 3c). By June, little barley seeds visibly littered the open soil in the middle of traces (Figure 3a). Clumps of bison fur hung in stands of little barley growing in wallows (Figure 3b), and two out of the 15 dung samples we collected in June contained little barley seeds, providing more evidence for dispersal of this species’ seeds by both endo- and epizoochory. By following traces from wallow to wallow, we were able to harvest from continuous dense stands of little barley during our June visit (Figure 3). In the bison exclusion area, little barley was common along the road and near the buildings, its normal weedy habitat, but was entirely absent >1 m from the road within the prairie.
Maygrass is a much less common grass than little barley and is not considered weedy. Nevertheless, we have previously mostly encountered it in agricultural field margins and along roads, and others have reported similar habitats (Cowan, 1978). Where it does occur in other prairie remnants lacking bison that Mueller has visited, it is as scattered individual plants that are difficult to harvest. At TGPP, we located two large populations of maygrass growing in bison wallows (Figures 3c and 5). We did not observe any maygrass in the bison exclusion area or outside of wallows. Maygrass seeds were present in one of the 15 dung samples we collected in May. Since Rosas et al. (2008) did not document maygrass in bison dung in their study, this represents the first direct evidence for bison dispersal of this species’ seeds. We also observed clumps of maygrass growing out of old bison dung. While this species did not form patches as dense as those we have observed in some anthropogenic settings, the populations were extensive and interspersed with dense patches of both little barely and seedlings of sumpweed (Figure 5).

Wallow dominated by three lost crop species, from top: Black arrow: maygrass; white arrow: little barley; gray arrow: sumpweed.
During our spring visits, it was already clear that sumpweed is very strongly associated with bison wallows from the abundance of seedlings in these settings (Figure 5). A sump, like a wallow, is a depression that is periodically full of water, so it is possible that the common name actually derives from early observations of this plant growing in wallows. Dense, homogenous patches of sumpweed covered large areas within wallows and extended up the traces that connected them (Figure 4b). Populations of sumpweed (and the wallows that contained them) conspicuously ended at the bison exclusion fence (Figure 4a). In October, vast populations of sumpweed were visible from a distance and were exclusively associated with traces, wallows, and river and pond margins (Figure 4c). In the bison exclusion area, like little barley, sumpweed only occurred near the roads (especially in roadside ditches). The dung samples we collected during these visits have not yet been analyzed, but sumpweed seeds were visible to the naked eye when we were washing these samples in the field. As with little barley and maygrass in the spring, we were able to harvest from continuous stands by following traces from wallow to wallow (Figure 4). We also observed wild sunflowers growing in wallows with sumpweed (Figure 4d), and Polygonum ramosissimum (a close relative of the lost crop, P. erectum) growing along traces and on a trampled lakeshore.
Conclusions and directions for future research
All three of our hypotheses were strongly supported by our initial year of field work at TGPP. We observed dense patches of crop progenitors growing along traces and in wallows, but only observed these species along roads and near buildings elsewhere on the preserve. They did not occur in the ungrazed portion of the prairie, but were very conspicuous species in the grazed portion (see Figures 3 and 4). By following recently trodden traces through the prairie, we were able to harvest from continuous stands of little barley and maygrass (in June) and sumpweed (in October). While much more limited in distribution, we also observed a species of Polygonum closely related to the crop progenitor, and wild sunflowers in bison wallows, and did not encounter either of these species in the ungrazed areas. We also found it much easier to move through the prairie on traces than to venture off of them. The ungrazed prairie felt treacherous because of the risk of stepping into burrows or onto snakes, and disorienting because of its homogeneity. These observations support a scenario in which ancient people would have moved through the prairie along traces, where they existed. If they did so, they certainly would have encountered dense of stands of the same species they eventually domesticated. We must stress how surprised we were to find little barley, maygrass, and sumpweed as the three dominant species in many wallows. Although Mueller has spent several years documenting populations of crop progenitors across eastern North America, she has never observed a stand of little barley and maygrass with a dense mat of sumpweed seedlings emerging beneath them except in bison wallows (Figure 5).
These data are experiential and qualitative. In future seasons, we will return to the TGPP and visit prairies within the former prairie peninsula in order to quantify the distribution and abundance of crop progenitors under different management regimes. We also have yet to investigate the relationship between the stands of crop progenitors we observed and the frequency and intensity of fire. The stands of crop progenitors we observed had the appearance of fields, and we would like to compare their productivity to the yields we have been able to achieve in experimental gardens. Finally, the question of how these species went from naturally occurring in harvestable stands in bison wallows to being domesticated by people remains open. We hypothesize that as humans began saving and planting seeds, the relaxation of selection for small seeds and robust seed protections, which are necessary adaptions for plants whose seeds are dispersed in dung, may have been enough to result in the changes we see in domesticated specimens. Thinning stands, which can increase yield, may have selected both for larger and earlier germinating seeds. Finally, opening new habitats and planting seed, when it occurred, would have exerted strong selective pressure by resulting in populations sprung only from seeds with little or no inherent dormancy. We plan to explore these questions through long-term experiments which are already underway.
Further unanswered questions and directions for research lie in the archeological record. Bison bones may be much more common in the Holocene archeological and paleontological record of the prairie peninsula than most archaeologists realize. Dating just one kill site in Illinois (Lonza-Caterpillar) revealed that bison inhabited prairies east of the Mississippi river throughout the Holocene, and that people hunted them (Harn and Martin, 2006). Meanwhile, in Missouri and Arkansas, McMillan reports dozens of undated bison bone finds (McMillan, 2012). Further east, the most famous Holocene bison kill site is Big Bone Lick, Kentucky. The earliest date on the Holocene bison from Big Bone Lick is cal. 700–550 BP (Genheimer and Rieveschl, 2013). However, the 6 m deep Holocene floodplain deposits from which bison bones have been recovered date back to 5000 BP, leaving the question of older bison in the Ohio valley open (Tankersley et al., 2015). Further up the Ohio river, the Patton Bog site provides evidence for burning and the formation of prairies beginning around 2500 BP as far east as the western Appalachian front (Abrams et al., 2014). This is consistent with older paleoecological records from the western Appalachian front (Cliff Pond Palace and Icehouse Bottom, see Figure 1), which also provide evidence for increasing clearance of forests and fire-tolerant species around this time (Delcourt et al., 1986, 1998). More fine-grained paleoecological records in the lowlands and intensive dating of Holocene bison bones are needed to establish when the easternmost prairie peninsula came into existence, and whether or not bison inhabited it.
Stable isotopic methods could be used to directly test our hypotheses by (1) revealing the C3/C4 make-up of the prairies grazed by ancient bison, and; (2) ascertaining whether pre-domesticated archaeobotanical specimens were grown in nitrogen-enriched habitats, like wallows. Ungrazed tallgrass prairies are C4 landscapes dominated by four perennial, warm season bunch grasses (Sorghastrum nutans, Andropogon gerardi, Schizachyrium scoparium, Panicum virgatum). However, the proportions of C3 and C4 species vary along a latitudinal gradient (Tieszen et al., 1997), seasonally, and according to the species composition of the landscape (Tieszen, 1994), which is strongly effected by bison grazing. For example, Widga (2006) interpreted low δ13C values (compared to those expected for animals grazing on a tallgrass prairie) for the Late Holocene bison at Big Bone Lick as an indication that these bison were browsing on woody C3 vegetation. Alternatively, they may have inhabited a patchier and more heterogeneous prairie, like the one at TGPP, that was less dominated by C4 grasses. Increasing both the comparative sample of stable isotope values from modern bison bones and teeth on eastern prairies and the ancient sample from Holocene archeological sites might enable us to reconstruct some aspects of ancient bison diets and the species composition of the prairies or savannas they inhabited. Stable nitrogen isotopes can be used to determine if ancient seeds came from plants that grew in fertilized habitats (Bogaard et al., 2013). Since ancient eastern North America lacked any large domesticated animals, nitrogen enrichment in early crop progenitor specimens would constitute fairly strong evidence that they were collected from bison wallows, or that bison dung was collected and distributed in fields and gardens.
We present these preliminary observations as an invitation to researchers to consider the role of bison as co-creators of heterogeneous landscapes of disturbance, which gave rise to a unique and diverse agroecology in eastern North America. The return of the bison to tallgrass prairies presents an opportunity to witness the ecological effects of their interaction with various kinds of human management. North America’s tallgrass prairies were largely displaced by industrial agriculture, and these ecosystems are generally valued for their contribution to biodiversity, rather than as sources of useful plants and animals for human societies. Resolving the nature of humanity’s deep relationship with bison and the tallgrass prairies could contribute to better restoration and conservation plans for eastern prairies and help prairie managers re-think the potential of these ecosystems by considering the management practices of Indigenous people as models. Indigenous people in the midcontinent created resilient and biodiverse landscapes rich in foods for people by managing floodplain ecosystems rather than using levees and dams to convert them to monocultures, and by using fire and multispecies interactions to create mosaic prairie-savanna-woodland landscapes that provided a variety of resources on a local scale. Rather than beautiful artifacts preserved for the survival of charismatic species, the tallgrass prairies have the potential to be re-integrated into productive human landscapes, a move which would surely make more space for them in American’s breadbasket.
Footnotes
Acknowledgements
We completed this fieldwork thanks to a research grant from the Haus der Kulturen der Welt and the Max Planck Institute for the Science of Human History as part of the project Mississippi. An Anthropocene River. We would like to thank Christoph Rosol and Jesse Vogler for their roles in organizing the research and events of the Anthropocene River project in St. Louis, where we presented early results from this research. We would also like to thank Robert Hamilton and the staff of the Joseph H. Williams Tallgrass Prairie Preserve for facilitating this research, Mike Palmer for providing his species occurrence data, and Sara Laskow and Gayle Fritz for their help and company during our October visit to TGPP.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Haus der Kulteren der Welt and the Max Planck Institute for the Science of Human History