
Abstract
Peptide-based vaccines, utilizing defined B- and T-cell epitopes, have emerged over the past two decades as promising alternatives to conventional vaccine platforms. Their key advantages, including excellent safety profiles, precise immunological targeting, and ease of manufacture, position them as a versatile tool in modern immunology. However, the widespread clinical application of these vaccines is significantly challenged by inherent limitations, most notably their low inherent immunogenicity and susceptibility to enzymatic degradation in vivo, leading to structural instability and short half-lives. This review systematically examines the innovative strategies being developed to overcome these critical obstacles. It delves into advances in rational vaccine design, highlighting the pivotal role of bioinformatics and computational tools for precise epitope selection and the engineering of sophisticated multi-epitope constructs that elicit broader immune responses. Furthermore, we provide a comprehensive analysis of novel drug delivery systems, such as a diverse range of nanoparticle carriers (e.g., liposomes, polymer-based), and review the mechanism of action of next-generation adjuvants. These technologies are crucial for enhancing antigen presentation, protecting the peptide payload, and promoting robust, durable cellular and humoral immunity. By synthesizing current literature from databases like PubMed and Google Scholar, this review offers a detailed overview of recent progress in peptide vaccine development against viral pathogens, drawing on key findings from both preclinical and clinical studies. It highlights key findings from preclinical and clinical studies and provides insights into the mechanistic actions and future potential of these precision vaccines in combating evolving viral threats.
Plain language summary
Vaccines are our most powerful tool for preventing infectious diseases. A new and promising type of vaccine uses small, specific pieces of a virus, called peptides, rather than the whole virus. These peptide-based vaccines offer important benefits: they are very safe, can be precisely designed, and are simpler to produce than traditional vaccines. However, they also face challenges—they are often not strong enough to stimulate a strong immune response, can break down quickly in the body, and may not work equally well in all people. This review explores the latest scientific strategies to overcome these limitations. We look at how researchers are using computer modeling to select the most effective viral peptides, combining multiple peptides to improve effectiveness, and packaging them into nanoparticles or using special immune boosters to enhance their delivery and potency. By examining recent studies, this article provides a comprehensive overview of the advances and future potential of peptide vaccines in the fight against viral infections.

Background
Designing a vaccine is one of the most impactful public health strategies, contributing to the global elimination of diseases such as smallpox and polio. 1 Today, many existing vaccines are based on inactive (killed) or attenuated pathogens or toxins secreted by pathogens. 2 In contrast to mRNA or virus-like particle (VLP) vaccinations, which depend on intracellular antigen production or structural imitation, peptide vaccines function by directly administering specific antigenic sequences, providing high accuracy but frequently necessitating adjuvants or delivery mechanisms to augment immunogenicity.3,4 In recent years, our growing understanding of the immune system has significantly improved vaccine development strategies. 5 One of the most important advancements is the application of bioinformatics tools, which enable researchers to rapidly identify and analyze specific microbial protein antigens. 6 These computational methods, particularly reverse vaccinology, are now widely used to design recombinant and peptide-based vaccines with greater efficiency and accuracy.5,6
Peptide-based vaccines offer several advantages over whole-protein vaccines. They are easier and cheaper to produce, are more efficiently absorbed and processed by antigen-presenting cells, and do not require complex folding into a tertiary structure to be effective. These vaccines contain short, immunogenic fragments of pathogenic proteins that are capable of stimulating a potent immune response. 7
Search strategy and inclusion criteria
This review aims to find peer-reviewed papers centered on peptide-based vaccine development targeting viral diseases. Investigations were conducted across the PubMed, Scopus, and Google Scholar databases. A tailored search strategy was established and modified for each database utilizing combinations of the following keywords: (“peptide vaccine” OR “epitope vaccine” OR “synthetic peptide”) AND (“virus” OR “viral infection” OR “viral immunology”). The screening process and data extraction were performed by a single investigator from January to May 2025.
Variables for analysis were chosen based on the PECO (Population, Exposure, Comparator, Outcome) framework, tailored to the context of vaccine development research. This encompassed the host population type (e.g., animal models, human subjects), vaccine formulation and delivery mechanisms (exposure), reference or control formulations (comparison), and documented immunological or protective responses (outcome). In the preliminary screening process, titles and abstracts were evaluated, and duplicate entries were eliminated. 8
The peptide as a vaccine
Recent advances in vaccine design tools, such as high-throughput sequencing and immunoinformatics, have enabled the identification of immunodominant epitopes within pathogens that can effectively stimulate the immune system. 9 One of the earliest examples of this concept was demonstrated in 1963 using the tobacco mosaic virus as a model for synthetic peptide-based antigen design. Furthermore, the development of protein sequencing techniques in 1977, along with Merrifield’s solid-phase peptide synthesis method introduced in 1963, significantly contributed to the growth of peptide-based vaccine research. 10 Unlike mRNA or VLP vaccines, which rely on intracellular antigen synthesis or structural mimicry, peptide vaccines act through direct administration of well-defined antigenic sequences, offering high precision but often requiring adjuvants or delivery systems to enhance immunogenicity.
The first clear demonstration that synthetic peptides can elicit a protective immune response was reported in 1982 against the Hand, Foot, and Mouth Disease (FMDV). 11 Short peptide vaccines, composed of minimal T- or B-cell epitopes, are easy to synthesize but often poorly immunogenic. To overcome the limited immunogenicity of short peptide vaccines, synthetic long peptide (SLP) vaccines have been developed. These SLPs are preferentially internalized by APCs and processed via both MHC class I and class II pathways, thereby eliciting coordinated CD8+ and CD4+ T-cell responses.12,13 Another promising strategy in peptide-based vaccination is the use of peptide-pulsed dendritic cell vaccines. 14 These vaccines involve ex vivo loading of autologous dendritic cells with viral or tumor-associated antigenic peptides, followed by re-infusion into the patient to stimulate antigen-specific T-cell responses. 14 Multiple clinical trials have demonstrated the safety and immunogenicity of dendritic cell-based vaccines in settings involving viral infections and cancer.14,15
Approaches in peptide vaccine synthesis and design
The most common method for synthesizing peptides is solid-phase peptide synthesis.10,16 In this technique, amino acids are added one by one to a growing peptide chain that is attached to a solid support (resin). 10 First, a protected amino acid is linked to the resin through its carboxyl group. Then, its protecting group is removed to expose the amino end. 10 The next amino acid is then activated and added. 10 This cycle of deprotection and coupling continues until the full peptide sequence is built. 16 At the end, all protecting groups are removed, and the complete peptide is detached from the resin.10,16 Following synthesis, the crude peptides are typically purified using high-performance liquid chromatography to ensure purity and functionality. 17
Mass spectrometric techniques, particularly electrospray ionization coupled with tandem mass spectrometry (ESI-MS/MS), provide a powerful and efficient approach for identifying pathogen-derived peptides that are naturally presented by HLA molecules.18,19 This approach, commonly referred to as immunopeptidomics, allows for the comprehensive profiling of naturally processed peptides presented by HLA molecules. This strategy plays a critical role in the development of peptide-based vaccines by enabling the selection of immunologically relevant epitopes. 19
Another promising approach in vaccine development is reverse vaccinology, which leverages bioinformatics tools and peptide databases to identify potential antigenic peptides. Compared to traditional experimental methods, computational screening provides a faster and more cost-effective strategy for discovering vaccine candidates. Numerous therapeutic peptides have been successfully designed using computational algorithms and later validated experimentally.7,20 Despite its advantages, reverse vaccinology relies heavily on in silico predictions, which require rigorous in vitro and in vivo validation to ensure clinical relevance. 21 Table 1 summarizes some commonly used databases and their applications in designing peptide-based vaccines through reverse vaccinology.
Some bioinformatics tools in the field of reverse vaccinology.
In the reverse vaccinology workflow, the process begins with the identification of pathogen-derived target proteins, which are then analyzed using immunoinformatics tools to predict potential B-cell and T-cell epitopes. Epitopes with the highest predicted antigenicity, immunogenicity, and safety profiles (i.e., non-toxic and non-allergenic) are shortlisted for further evaluation.22,23 Subsequently, selected peptide vaccine candidates are tested in animal models to assess their ability to elicit robust immune responses. These preclinical results guide the optimization and selection of the most promising candidates for advancement into human clinical trials (Figure 1). 21
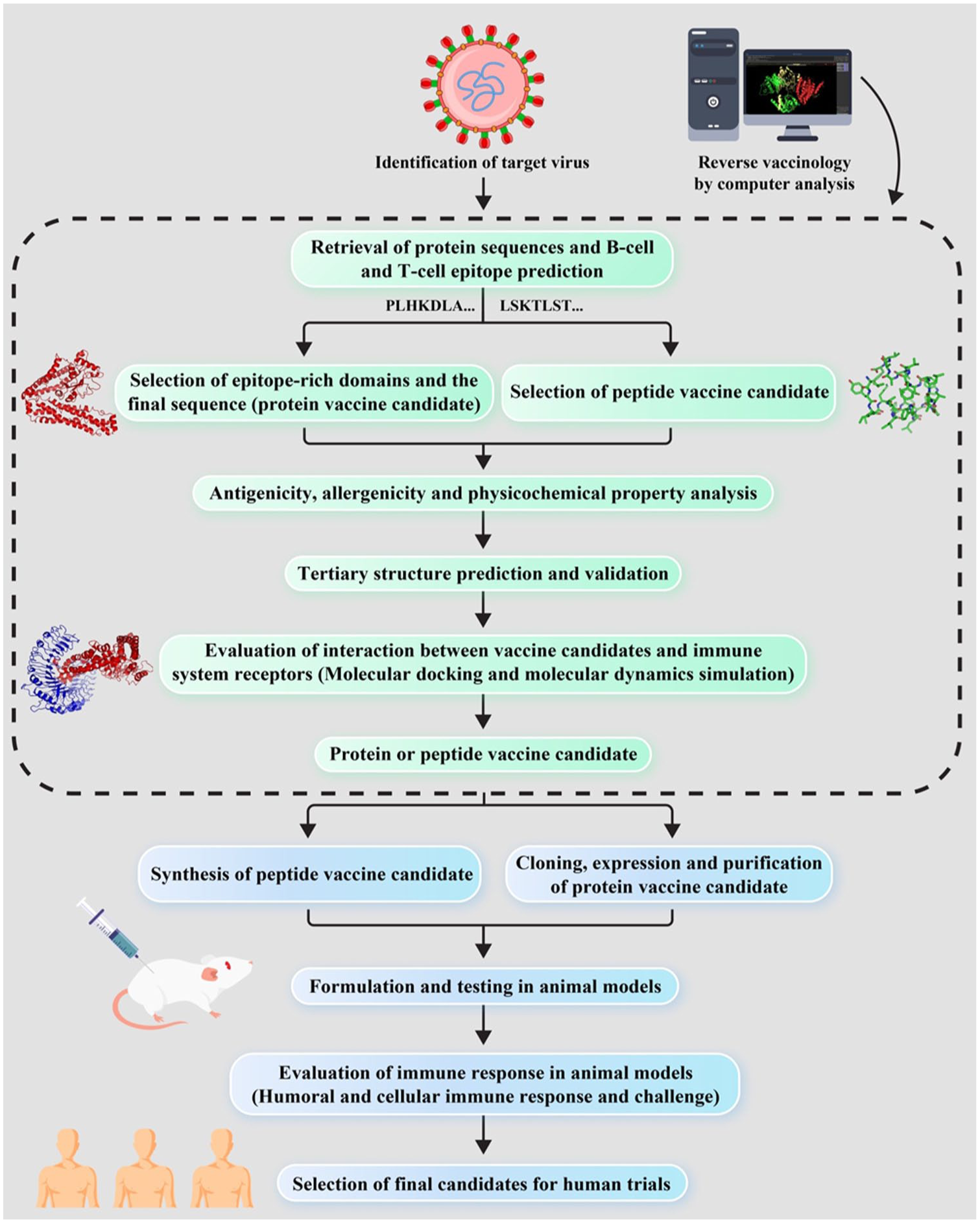
Flow diagram of design strategy, representing the steps of the construction of the peptide-based vaccine.
Mechanism of peptide vaccines
Mechanism of peptide vaccines in inducing cellular immunity: activation of cytotoxic T-lymphocytes
Numerous studies have demonstrated that peptide-based vaccines can effectively activate cytotoxic T-lymphocytes (CTLs). In this pathway, peptides derived from intracellular proteins are processed by the proteasome, a multi-subunit protease complex, into short antigenic fragments. These peptides are then translocated into the lumen of the endoplasmic reticulum (ER) via the transporter associated with antigen processing (TAP). 33 Within the ER, they are loaded onto major histocompatibility complex class I (MHC I) molecules. 34 The peptide–MHC I complexes are subsequently transported through the Golgi apparatus to the cell surface, where they are presented to CD8+ T cells, triggering CTL responses. 35
A major challenge in eliciting strong cellular immunity using exogenous peptide vaccines compared to naturally processed endogenous antigens is their susceptibility to degradation by extracellular proteases. 36 This rapid enzymatic degradation can significantly limit the stability and immunogenicity of the vaccine peptides, thereby reducing their efficacy in stimulating CTLs (Figure 2). 37

Schematic representation of the main pathways of the immune response stimulated by the peptide vaccine.
Mechanism of peptide vaccine to induce a humoral immune response
Peptide-based vaccines can stimulate humoral immune responses by activating B cells in a T-cell-dependent manner. Following vaccine administration, exogenous peptide antigens are taken up by professional antigen-presenting cells (APCs), such as dendritic cells or B cells, and processed within endosomal compartments. 38 These peptide fragments are then presented on the cell surface bound to major histocompatibility complex class II (MHC II) molecules. 38 CD4+ T helper (TH) cells recognize the peptide–MHC II complex and become activated, subsequently providing essential co-stimulatory signals and cytokines to B cells. B cells that have internalized the same antigen via their surface immunoglobulin receptors also present processed peptides on MHC II. Upon receiving help from activated TH cells, these B cells undergo clonal expansion, somatic hypermutation, class-switch recombination, and ultimately differentiate into long-lived plasma cells capable of secreting high-affinity antigen-specific antibodies (Figure 2).38–40
Strategies to improve peptide vaccine immunogenicity: Adjuvants
Peptide-based vaccines, despite incorporating optimal B- and T-cell epitopes, often exhibit limited immunogenicity and therefore require the use of adjuvants to achieve stronger immune responses. 41 Adjuvants are broadly defined as agents that boost the immune system’s response to an antigen. The most historically used adjuvant is aluminum-based compounds (such as aluminum hydroxide or phosphate, collectively referred to as Alum), which have been in use since the 1920s. 42 Although the exact mechanism of Alum remains incompletely understood, recent studies suggest it functions in part by activating dendritic cells and promoting antigen uptake and presentation. 43
However, alum has limitations, including local inflammation, erythema, subcutaneous granulomas, and occasional hypersensitivity reactions. 44 Moreover, it tends to skew the immune response toward a Th2 phenotype, which may not be optimal for vaccines that require cell-mediated immunity. 44
To address these limitations, attention has shifted to toll-like receptor (TLR) agonists as more potent and versatile adjuvants. 45 TLRs are pattern-recognition receptors that activate dendritic cells and drive Th1-type immune responses, which are more effective in combating intracellular pathogens and in cancer immunotherapy. 46 Notably, TLR agonists have been shown to induce higher-avidity T-cell responses than Alum. One of the most prominent examples is monophosphoryl lipid A (MPLA), a lipopolysaccharide (LPS)-derived TLR4 agonist that is typically extracted from Salmonella minnesota Re595 and has been detoxified to reduce toxicity while maintaining TLR4 stimulation capacity and has been approved for human use in licensed vaccines such as Cervarix. 47 Clinical trials have confirmed its favorable safety and efficacy profile, making it a valuable component in modern adjuvant systems. 48
Overall, while traditional adjuvants like alum have historically enhanced peptide vaccine responses, their limitations highlight the need for modern alternatives such as TLR agonists, which offer stronger and more targeted immune activation
Peptide vaccine delivery systems and their immunological impact
The route of administration and the formulation of peptide-based vaccines critically influence peptide stability, biodistribution, and antigen processing. 49 Due to their inherently low immunogenicity and susceptibility to enzymatic degradation, peptides require a specific route of administration and formulation to ensure peptide stability, biodistribution, and antigen processing. 49 Therefore, optimized peptide vaccines are now increasingly designed in multifunctional drug delivery platforms such as nanoparticles, liposomes, VLP, and hydrogels, which can enhance cellular uptake, ensure sustained antigen release, and simultaneously provide immunostimulatory adjuvants.49–51 These strategies not only protect the peptides but also improve lymph node targeting and cross-presentation via MHC class I pathways.49–51 A summary of current delivery approaches for peptide vaccines is provided in Table 2.
Methods of delivery of peptide vaccine.

CPPs, PLGA, poly lactic-co-glycolic acid.
Cell-penetrating peptides
Protein-based vaccines are predominantly processed via the endocytic pathway and presented on MHC class II molecules, leading primarily to the activation of CD4+ T helper cells rather than CTLs. 52 Therefore, to elicit robust CD8+ T-cell responses, it is essential to develop strategies that enable cross-presentation of protein antigens through the MHC class I pathway. 52 One promising approach involves the use of cell-penetrating peptides (CPPs), short cationic or amphipathic molecules capable of translocating across cellular membranes and delivering macromolecules directly into the cytosol. 52 CPPs have been widely applied in drug delivery and DNA transfection, as well as in peptide- and protein-based vaccines. 52
In preclinical models, antigen conjugation to CPPs has been reported to enhance peptide vaccine potency—up to 25-fold in one study—by promoting antigen uptake and cross-presentation by dendritic cells, while novel CPPs derived from the S4 domain of the shaker-type potassium channel have further enhanced MHC I presentation and CD8+ activation when fused to tumor-specific epitopes.53,54 Moreover, CPP-based constructs have shown efficacy in antiviral vaccine platforms, inducing both humoral and cellular immune responses against infections such as Human Immunodeficiency Virus (HIV) and HBV, and providing long-term protection in tumor models like human papillomavirus-16 (HPV-16). 52
Compared to conventional nanoparticle carriers such as poly lactic-co-glycolic acid (PLGA) and liposomes, CPPs facilitate more rapid intracellular delivery; however, they often benefit from co-formulation with stabilizing agents to improve antigen retention and prolong immune activation. 55
Nanoparticle-based platforms
A promising delivery strategy for peptide vaccines involves nanoparticles. These particles act as both carriers and immune modulators by protecting the antigen, enabling targeted delivery to antigen-presenting cells, and enhancing cross-presentation to strengthen the immune response. In addition, nanoparticles can provide multivalent antigen display and intrinsic adjuvanticity, thereby amplifying both humoral and cellular immune responses. 56 Recent studies emphasize that nanoparticle adjuvants can not only enhance antigen uptake but also directly act as immunostimulatory agents by promoting dendritic cell maturation and cytokine production, without the need for additional adjuvants. This dual role has made them highly attractive in both veterinary and human vaccine design. 57
Among the various nanoparticle platforms, self-assembling protein nanostructures have attracted considerable attention. 58 One prominent example is apoferritin (AFt), a natural cage-like protein composed of 24 subunits, which self-assembles into a highly stable and biocompatible nanocage. 58 This structural organization enables the precise loading and presentation of peptide antigens on the surface, as well as the potential co-delivery of immunostimulatory molecules, such as TLR agonists. 59 Ferritin-based platforms have been successfully used in preclinical models for influenza and SARS-CoV-2 (COVID-19) vaccines, demonstrating robust enhancement of both humoral and cellular immune responses. Recent studies have demonstrated that self-assembling peptides can organize into nanostructures such as fibers, micelles, and hydrogels, which not only enable multivalent antigen display but also act as precisely defined chemical adjuvants. 60 These systems have been shown to induce potent immune responses even in the absence of additional adjuvants, while maintaining high stability, biocompatibility, and reducing the required antigen dose. 60
Glycoconjugates and lipopeptide formulations
Conjugation of peptides with oligosaccharides, such as mannose or trehalose-based structures, significantly improves antigen uptake by dendritic cells via receptor-mediated endocytosis, particularly through C-type lectin receptors.61,62
Moreover, the inclusion of lipid moieties, such as Pam3Cys or other lipoamino acids, promotes strong Th1-biased immune responses and facilitates efficient delivery of the antigen to lymphoid tissues. 63 Compared to peptides alone, these glyco- and lipopeptide constructs elicit higher antibody titers and stronger T-cell responses, highlighting their potential in the development of next-generation peptide vaccines.64,65
Heat-shock proteins as immunomodulators and antigen carriers
Heat-Shock Proteins (HSPs), such as HSP60, are highly conserved molecular chaperones recognized by the immune system. 66 HSP60 interacts with TLR4 on antigen-presenting cells, bridging innate and adaptive immunity by promoting maturation and activation of dendritic cells and enhancing T-cell responses.66,67
In vaccine design, HSP60-derived peptides, such as HSP60-p458, have demonstrated effective carrier properties for bacterial antigens, leading to sustained protective immunity, particularly in aged animal models. 68 Recent studies further support the role of HSP60 in enhancing cellular immunity by regulating mitochondrial metabolism in memory T cells. 69 Specifically, HSP60 has been implicated in driving glutaminolysis in CD4+ T cells and fatty acid oxidation in CD8+ T cells following T-cell receptor (TCR) stimulation, thereby augmenting their effector functions. 69 These findings highlight the multifaceted potential of HSP60 as both an immunomodulatory agent and a peptide carrier, offering valuable applications in the development of next-generation peptide vaccines, particularly for infections and cancer.70,71
A diverse array of delivery platforms has been engineered to overcome the inherent limitations of peptide-based vaccines, each distinguished by its unique mechanism of action, manufacturing complexity, and stage of development. VLPs provide a safe and highly immunogenic platform for dense antigen presentation, effectively stimulating both CD4+ and CD8+ T-cell responses.3,4 Liposomes offer notable versatility, as they can be co-loaded with peptides and TLR agonists to simultaneously support CD4+ responses and facilitate cross-presentation for robust CD8+ T-cell activation, often serving as their own adjuvant. Similarly, polymeric particles, such as those composed of PLGA/PLA, enable prolonged antigen release and are particularly effective at promoting CD8+ T-cell immunity through cross-presentation. 55 The class of protein nanoparticles, including ferritin and self-assembling peptides, allows for multivalent antigen presentation, fostering a balanced CD4+ and CD8+ response. 72 For localized and sustained delivery, hydrogels act as depots that enhance the priming of both T-cell lineages and can be tailored for mucosal or systemic delivery, though their chemistry, manufacturing, and controls (CMC) standardization and late-phase safety data remain underdeveloped. 73 CPPs consistently enhance CD8+ T-cell responses by targeting cross-presentation in Batf3+ dendritic cells; however, while their CMC is straightforward, their stability and pharmacokinetic profile pose challenges, and human clinical data is still nascent. 74 Finally, microneedles present a logistically advantageous strategy by efficiently targeting the dense network of skin antigen-presenting cells, which enables potent systemic and mucosal-adjacent immunity with a reduced antigen dose and enhanced thermostability that can circumvent cold-chain requirements. 75 However, their CMC challenges are primarily centered on ensuring the reproducibility of matrix casting and coating processes.
Liposomes
Liposomes have emerged as a versatile and biocompatible delivery platform for peptide-based vaccines, enabling efficient encapsulation, protection from enzymatic degradation, and targeted delivery to antigen-presenting cells.49,51,76
Recent studies have demonstrated that liposomal formulations can significantly enhance the induction of cytotoxic CD8+ T-cell responses, particularly in the context of viral infections. 76 Furthermore, the incorporation of lipid moieties such as covalently attached fatty acids into peptide structures has been shown to improve membrane integration, uptake by dendritic cells, and subsequent antigen cross-presentation, thereby enhancing the immunogenicity of peptide vaccines.77,78
Virus-like particles
Another advanced delivery strategy for peptide antigens employs VLPs non-infectious, non-replicating nanostructures that self-assemble from viral capsid proteins and mimic natural virions in terms of size (20–200 nm) and geometry, without containing viral genomes.79,80 These VLPs enable dense, multivalent display of peptide epitopes that enhance lymphatic drainage and efficient uptake by dendritic cells. After internalization, peptides are cross-presented through MHC class I and II pathways, thereby eliciting robust CD8+ cytotoxic and CD4+ helper T-cell responses.79,80 Several VLP-based vaccines, such as those targeting hepatitis B virus (HBV) and HPV, have received regulatory approval and have demonstrated excellent safety and immunogenicity in humans.79,81 Additionally, many chimeric VLP platforms incorporating exogenous viral or tumor peptides are currently in preclinical or clinical stages, offering promising avenues for emerging infectious disease and cancer vaccines.79,81
Among these VLP platforms, Papaya Mosaic Virus (PapMV)-derived VLPs represent a notable example, as they combine delivery and innate immune stimulation within a single nanoparticle system. PapMV-derived VLPs are rod-shaped protein assemblies composed of PapMV coat protein surrounding non-coding single-stranded RNA. 72 These nanoparticles efficiently engage endosomal TLR7/8 pathways, acting both as antigen carriers and innate immune activators. 72 Antigenic peptides fused or chemically coupled to PapMV frequently result in strong humoral immunity, notably inducing Immunoglobulin G (IgG) responses with Th1 bias (e.g., IgG2a isotype), when targeting epitopes from hepatitis C virus (HCV), influenza, or other pathogens.72,82 PapMV-based vaccines have also demonstrated cross-presentation to CD8+ T cells, supporting cytotoxic T-cell activation.72,83 Chimeric PapMV VLPs currently in development include those displaying exogenous HCV epitopes and have shown potent immunogenicity in preclinical models. 83
Polymeric particles
Degradable polymeric particles composed of poly lactic-co-glycolic acid (PLGA) or polylactic acid (PLA) have emerged as highly effective carriers for peptide antigens. 84 These FDA-approved polymers enable controlled and sustained antigen release, ranging from several days to weeks or even months, thus serving as built-in antigen depots that gradually expose the immune system. 84 Upon uptake by antigen-presenting cells, PLGA particles are internalized and trafficked through the endosomal compartment, where acid-mediated erosion facilitates antigen release and efficient cross-presentation via MHC class I and II pathways. 85 This promotes robust activation of both CD8+ cytotoxic and CD4+ helper T-cell responses. 85 Furthermore, co-encapsulation of peptide antigens with innate immune stimulators (such as TLR agonists or invariant Natural Killer T cell (iNKT) agonists) greatly enhances the magnitude and quality of cellular immunity, including therapeutic models of cancer and infectious disease. 86 Particle size, polymer composition, and surface modifications can be optimized to fine-tune release kinetics and dendritic cell targeting. 87
Self-assembling peptide hydrogels
Self-assembling peptide hydrogels represent a novel class of biomaterials designed for sustained local release of peptide antigens and immune stimulators. These hydrogels form nanofibrous scaffolds that act as both delivery depots and immune modulators. By providing prolonged antigen exposure and recruiting immune cells to the injection site, they enhance antigen uptake, dendritic cell activation, and T-cell priming. Certain hydrogels also exhibit inherent adjuvant activity, further supporting a robust cellular and humoral immune response. 73
Non-invasive routes
Non-invasive administration routes such as intranasal and transdermal microneedle-based delivery are gaining attention for peptide vaccines. Intranasal delivery can target mucosal immunity and facilitate uptake by the nasal-associated lymphoid tissue, making it an ideal approach for addressing respiratory pathogens. 88 Meanwhile, dissolvable microneedles painlessly penetrate the skin barrier, releasing peptide antigens directly to epidermal antigen-presenting cells, including Langerhans cells and dermal dendritic cells, which enhances systemic and local immune responses.88,89
Taken together, innovative delivery systems such as liposomes, VLPs, polymeric particles, CPPs, and hydrogels have greatly improved the stability, uptake, and immunogenicity of peptide vaccines. Alongside non-invasive routes like intranasal and microneedle delivery, these approaches overcome the main limitations of peptides and open the way for effective next-generation vaccines.
Peptides in the prevention of virus infection
As of August 2024, more than 350 interventional clinical trials involving peptide-based vaccines were registered on ClinicalTrials.gov. These trials were identified using the search term “peptide vaccine” with filters for interventional studies (Phases I–IV). Most studies were in early stages, including approximately 250 in phase I and over 150 in phase II, while only 10 studies had progressed to Phase III, of which three focused on SARS-CoV-2. Several factors limit the progression of peptide-based vaccines to Phase III trials. 90 Genetic diversity in HLA molecules can reduce population coverage, while peptides often exhibit low immunogenicity, requiring the use of adjuvants, whose regulatory approval can be challenging.76,90,91 Manufacturing and scalability issues, along with the complexity of trial design and endpoints, further impede development.76,91 Financial constraints and market considerations also restrict late-stage studies.76,91 Several clinical trials investigating peptide-based vaccines against viral infections are summarized in Table 3.
Worldwide clinical trials of peptide vaccines against viral infections.

In the following sections, we explore viral components—such as structural and non-structural proteins—that have been used as targets in peptide vaccine design to elicit protective immune responses.
HIV
Antiretroviral therapy (ART) effectively suppresses HIV, delays disease progression, and improves quality of life, yet it cannot eradicate the virus. 92 Consequently, developing an effective prophylactic or therapeutic HIV vaccine remains a high priority. Multiple components of the HIV proteome have been explored as peptide vaccine targets, including the CD4-binding site, Nef, and Envelope (Env) glycoproteins.93,94 For example, the HIV-1 fusion peptide 8 (FP8), an 8-amino acid epitope located within the highly conserved fusion peptide region of glycoprotein 41 (gp41), plays a critical role in viral entry by mediating membrane fusion. Its structural conservation across diverse HIV strains makes it a vulnerable site of neutralization, and FP8-based immunogens have demonstrated the ability to elicit both humoral and cellular immune responses. Similarly, Nef-derived antigens have shown promising immunogenicity, leading to the activation of cytotoxic T lymphocytes (CTLs) and robust antibody production. 93 Furthermore, peptides derived from Env proteins, particularly gp41, have been shown to induce strong humoral responses in animal models. 94
Several clinical trials have evaluated peptide-based therapeutic vaccines in HIV-infected individuals. Vacc-4x, a peptide vaccine targeting conserved domains of HIV p24, capsid protein of HIV, reduced viral load set point and enhanced HIV-specific T-cell responses in patients undergoing ART interruption in a phase II trial. 95 Similarly, intradermal administration of short HIV Group-specific Antigen (Gag) capsid (p24)-like peptides elicited durable CD4+ and CD8+ T-cell responses lasting over 7 years. 96 These findings underscore the potential of peptide vaccination to induce long-lasting cellular immunity even in chronically infected patients.
Building upon these findings, nanoparticle- and dendritic cell-based delivery systems are now being explored to improve antigen presentation, enhance immune activation, and achieve more potent and durable therapeutic effects. Preclinical evaluation of vaccines based on multiple minimal T-cell epitopes combined with cationic liposomal adjuvants (CAF01) indicates safety in animal models and the capacity to elicit broad Th1/Th17 and CTL responses. 97 Dendritic cell-based approaches further improve antigen delivery, enhance immunological synapse formation with T cells, and leverage nanotechnology to potentiate cellular immunity. 98
Furthermore, engineered membrane proximal external region (MPER) peptide vaccines targeting a conserved region of gp41 near the viral membrane that is critical for integration and delivered via liposomal formulations have been shown to induce MPER-specific antibody responses in mice, with limited neutralizing activity, highlighting the potential and current limitations of MPER-targeted approaches. Liposome-based vaccines combining FP and MPER domains focused antibody responses on the N-terminal MPER region, highlighting the importance of lipid membrane presentation for effective humoral immunity. 99 Moreover, assessment of different crosslinkers between peptide antigens and carrier proteins has demonstrated that linker length and chemical properties can significantly affect antigenicity and antibody responses in fusion peptide-directed vaccines, emphasizing the importance of chemical design in optimizing vaccine efficacy. 100
Advanced computational approaches are being applied to optimize vaccine design. Immuno- and bioinformatics-assisted multi-epitope vaccines targeting B- and T-cell epitopes across the complete HIV genome show potential for broad population coverage, strong predicted immune responses, and stable TLR interactions. Broad strategies for peptide vaccination continue to focus on identifying protective HIV epitopes and optimizing delivery and adjuvant systems to enhance antibody responses. Ongoing research continues to explore these and other conserved HIV epitopes in the design of peptide-based vaccines aimed at inducing broad, long-lasting protective immunity.101,102
West Nile virus
West Nile Virus (WNV), the causative agent of West Nile fever, is a positive-sense single-stranded RNA virus from the Flaviviridae family. 103 Although vaccine development has progressed, no human vaccine has yet been licensed. 103 Several candidates, including live-attenuated, recombinant protein, inactivated, and viral vector-based vaccines, are in preclinical or early clinical stages. 103 Key WNV antigens such as pre-membrane protein (PrM), envelope protein (E), non-structural protein 3 (NS3), and non-structural protein 4B (NS4B) are under investigation, with domain III of the envelope protein (E-DIII) being a prominent subunit vaccine target. 104 E-DIII-based vaccines—especially when conjugated to VLPs or adjuvants—have shown strong neutralizing antibody responses and IFN-γ secretion in preclinical studies.103,104
Chimeric peptide vaccines, such as the p458-Ep15 construct—comprising Ep15, a 15-amino-acid peptide epitope, fused to Heat Shock Protein 60 (Hsp60) have demonstrated complete protection in murine models by inducing both neutralizing antibodies and interferon-gamma (IFN-γ) responses. 105 Computational approaches have further identified a minimal set of conserved T-cell epitopes (three CD8+ and one CD4+) covering approximately 97% of the global population. 106 Immunoinformatics analyses have suggested other B- and T-cell epitopes such as QINHHWHKSGSSSIG, FLVHREWFM, NPFVSVATANAKVLI, and NAYYVMTVGTKTFLV as candidates for multi-epitope vaccine design. 107
Moreover, T-cell-based vaccine platforms are under exploration. For example, single-chain HLA-A2 MHC class I trimers carrying immunodominant WNV peptides have successfully elicited protective CD8+ T-cell immunity in mice. 68 Together, these studies highlight the promise of combining humoral and cellular immunity in designing next-generation WNV vaccines.
Influenza virus
In 2009, a new H1N1 influenza virus triggered the first pandemic of the 21st century. 108 Influenza protective antigens, including hemagglutinin (HA), neuraminidase (NA), matrix protein 1 (M1), matrix protein 2 (M2) (especially the extracellular domain M2 or M2e), and nucleoprotein (NP), play a key role in universal vaccine design. M2e is highly conserved across influenza A subtypes, and animal studies consistently show M2e-based vaccines confer cross-protection via non-neutralizing mechanisms such as antibody-dependent cellular cytotoxicity (ADCC) and T-cell immunity.109,110 Vaccination with HA-derived peptides that activate ADCC has also been shown to provide partial protection, increasing survival and reducing viral load in mice despite not inducing neutralizing antibodies. 111
The most clinically advanced candidate is Multimeric-001 (M-001), composed of conserved linear epitopes from HA, NP, and M1. 112 Phase I/II trials demonstrated robust CD4+ T-cell responses and acceptable safety, although they did not enhance standard neutralizing antibody titers post-seasonal flu vaccine booster.112–114 Recent nanoparticle vaccines, such as OMN/OVX313, which construct a heptameric nanoparticle platform with repeated epitopes of M2e and nucleoprotein (NP), have shown cross-protection and improved survival in mice compared to M2e alone. An H5N1-based M2e tetrameric peptide vaccine demonstrated cross-protection against lethal H7N9 infection in mice, highlighting the potential of M2e-targeted peptide vaccines against diverse influenza A subtypes. 115 Heterosubtypic M1 and NP epitopes fused with hemokinin-1 enhanced immune responses and cross-protection against diverse influenza A strains. 116 However, some modified M158-66 peptides can induce non-relevant T-cells and even worsen pathology after viral challenge, highlighting the need for careful epitope selection. 117 Vaccine-induced antigen-specific regulatory T cells (Tregs) have also been reported to dampen antiviral immunity, but the use of adjuvants like Cytosine-phosphate-Guanine (CpG) can reduce these Tregs and strengthen protective responses. 118
Computational approaches have also been applied to design peptide vaccines, such as HA-derived peptides for H1N1, where in silico modeling identified candidates that induce strong humoral responses and may react with multiple viral subtypes. 119 Recent in silico studies have identified conserved peptides in the HA stem region, such as HA2 88–107, which contain multiple B- and T-cell epitopes with high population coverage and strong predicted binding to common HLA alleles. 120 These in silico approaches complement experimental peptide vaccine strategies, providing candidates with high population coverage and predicted efficacy. 120 Recent reviews emphasize that peptide vaccines targeting conserved viral regions can induce robust cellular immunity and may offer cross-protection against highly variable influenza strains, supporting their role in universal influenza vaccine strategies. Moreover, a new mRNA-lipid nanoparticle vaccine named Fluaxe, encoding chimeric hemagglutinin constructs (cHA heads fused with HA stalks) plus full-length M2, elicited broad cross-reactive antibodies, T-cell memory, and protection against diverse influenza A and B viruses in mice.109,110,121,122
Human cytomegalovirus
Human Cytomegalovirus (CMV), a member of the β-herpesvirus family, is highly prevalent in the global population and poses a significant threat to immunocompromised individuals such as neonates, organ transplant recipients, and people who are infected with HIV.123,124 Congenital CMV infection is a leading cause of neurodevelopmental disorders in children. Current antiviral treatments suffer from limitations such as poor oral bioavailability and dose-related toxicity, highlighting the need for novel prophylactic strategies, including peptide-based vaccines.125,126
Several studies have explored peptide vaccines designed to induce immune responses against CMV. 127 These vaccines often consist of CTL epitopes, such as the phosphoprotein 65 (pp65) peptide, covalently linked with T-helper epitopes like pan-DR epitope (PADRE) or tetanus toxoid sequences.127,128 In a phase I clinical trial conducted by La Rosa et al., peptide vaccines composed of PADRE-pp65 or tetanus-pp65, with or without the CpG-based adjuvant PF03512676, were administered to healthy HLA-A*0201-positive adults. 127 The combination of peptides with the CpG adjuvant induced robust pp65-specific CD8+ T-cell responses in most participants. 127 In contrast, the response rate in the absence of adjuvant was only about 30%. Adverse events were generally mild to moderate, including local skin reactions and flu-like symptoms. 127 Preclinical studies using HLA-A*0201-transgenic mice also confirmed that peptide combinations (PADRE or tetanus fused with pp65) could elicit cellular immune responses without adjuvants, although CpG significantly enhanced immunogenicity. 127
In recent years, the field has shifted toward more advanced platforms. Moderna’s mRNA-1647 vaccine, which encodes CMV glycoprotein B (gB) and the pentamer complex, has demonstrated potent neutralizing antibody and T-cell responses in phase I/II trials, with sustained effects for up to 3 years and a favorable safety profile. Additionally, vectors such as Modified Vaccinia Ankara, which encodes multiple CMV antigens (pp65, Immediate-Early protein 1 (IE1), Immediate-Early protein 2 (IE2)), have shown strong T-cell immunogenicity in transplant recipients, reducing CMV-related events. 129 These findings suggest that multi-epitope peptide formulations targeting both structural (gB, pp65) and non-structural (IE1, IE2) proteins could provide broader immune protection. 129
A SLP vaccine (PEP-CMV), comprising pp65 epitopes, was recently evaluated in a phase I trial in pediatric and young adult patients (ages 3–35) with gliomas or medulloblastomas. 130 The vaccine was generally well-tolerated (mostly grade 1–2 adverse events), and a significant increase in IFN-γ-producing T cells was observed. 130 However, progression-free survival (PFS) and overall survival (OS) remained limited (median PFS: 2.5 months; OS: 6.5 months), possibly due to the aggressive nature of the underlying malignancies. 130
Peptide vaccines conjugated to the non-toxic diphtheria toxin derivative Cross-Reacting Material 197 (CRM197), containing epitopes from pp65 and glycoprotein H (gH), induced dendritic cell maturation via the p38 mitogen-activated protein kinase pathway and elicited stronger humoral and cellular immune responses than inactivated CMV vaccines in murine models, highlighting the potential of carrier-conjugated peptide platforms. 131
Earlier studies have highlighted the potential of peptides derived from CMV glycoproteins UL130 and UL131, which are encoded by genes in the virus’s unique long region and are essential for viral entry into mucosal epithelial cells. 132 Early peptide-based approaches demonstrated the ability to induce neutralizing antibodies capable of blocking viral entry, providing initial proof-of-concept for targeting mucosal entry pathways in peptide vaccine design. 132
Clinical studies in CMV-seronegative individuals, including patients with end-stage renal disease, have demonstrated that peptide vaccination can elicit robust T-cell responses, and in these studies shared TCR motifs associated with CMVpp65 recognition were observed among responders, suggesting that TCR profiling could serve as a predictive biomarker for vaccine responsiveness and providing a rational basis for the design of next-generation peptide vaccines with enhanced efficacy. 133
Building on these insights, broader challenges for translating peptide vaccines into protection against congenital CMV remain. Clinical endpoints should focus on the reduction of fetal infection or disease severity, not solely on sterilizing immunity. 134 Vaccines must also elicit protective maternal antibodies to complement T-cell responses, guiding rational design of next-generation peptide vaccines for maximal cellular and humoral immunity, particularly in women of childbearing age. 134
Respiratory syncytial virus
Respiratory syncytial virus (RSV) is an enveloped negative-sense RNA virus that poses a serious threat, particularly to infants and elderly individuals. Several RSV antigens, notably the F (fusion) and G (attachment) glycoproteins, have been targeted in vaccine development. While vaccine constructs combining F and G proteins have induced robust immune responses in animal models (e.g., mice and primates), immunization with full-length recombinant F or G proteins has been associated with vaccine-enhanced disease upon subsequent RSV exposure, highlighting the need for safer alternatives.135,136
Epitope-based peptide vaccines represent a promising strategy by recapitulating critical neutralization epitopes while minimizing the risk of immunopathology associated with full-length proteins. In pioneering work, Michael W. Steward and colleagues screened a combinatorial solid-phase peptide library with an RSV-neutralizing monoclonal antibody, MAb 19, which recognizes a conformational epitope on the F protein. 137 They identified mimotopes (e.g., S1S: HWSISKPQ) that bound MAb 19 efficiently. When these were delivered as multiple antigen peptides and used to immunize BALB/c mice, the resulting anti-peptide antibodies inhibited MAb 19 binding and neutralized RSV infection in vitro. 137 Following viral challenge, vaccinated mice demonstrated markedly reduced lung viral loads (~98% reduction) and significantly less cellular infiltration compared to controls. 137
Yusibov and colleagues developed an alternative delivery platform using plant-virus-derived particles. 138 A 21-amino-acid G protein peptide (aa 170–190) was fused to the Alfalfa mosaic virus coat protein to create recombinant VLP (VMR RSV). 138 Administered to human dendritic cell cultures and non-human primates, these particles elicited potent CD4+ and CD8+ T cell responses and robust humoral immunity without inducing enhanced disease. 138
Additionally, Momtaz et al. applied an immunoinformatic approach to the matrix 2 protein 1 of RSV (RSV M2-1 protein), identifying T-cell epitopes with high antigenicity and broad population coverage. These epitopes showed promising potential to induce robust and safe cellular immune responses, highlighting M2-1 as a novel target for peptide-based vaccine design. 139
Building on these approaches, Alnajran et al. designed a multi-epitope peptide vaccine incorporating B-cell, CD4+, and CD8+ T-cell epitopes from the F and G proteins using a reverse vaccinology approach. The designed vaccine displayed high antigenicity, low instability, minimal allergenicity, and low predicted toxicity. In silico immune simulations suggested strong humoral and cellular responses, including increased IgG, IgM, interleukin-2 (IL-2), and IFN-γ, while molecular docking and dynamics analyses indicated stable binding to TLR1 and TLR4, supporting its potential efficacy and safety. 140
Recently, Naqvi et al. used a reverse vaccinology approach to predict and experimentally evaluate CD4+ and CD8+ T-cell epitopes from RSV F, G, and small hydrophobic protein (SH) proteins. 141 Selected epitopes induced strong cellular and humoral responses in BALB/c mice, including elevated IgG, IFN-γ, and cytotoxic T-cell activity. 141 Importantly, these epitopes were highly conserved across RSV strains and showed no significant homology with human proteins, supporting their potential as safe and broadly protective peptide vaccine candidates. 141
In summary, peptide-based RSV vaccines targeting multiple viral antigens, delivered through innovative platforms or designed via immunoinformatics and reverse vaccinology, show strong potential to induce broad and safe immune protection. These strategies provide a promising avenue for next-generation RSV vaccine development.
HFMD
Hand, Foot, and Mouth Disease (HFMD) is a highly contagious illness that primarily affects children under the age of 10. 142 Although it is often self-limiting, serious complications such as meningitis, encephalitis, acute flaccid paralysis, and neuro-respiratory syndrome may occur.142,143 The main causative agents are Enterovirus A71 (EV-A71) and Coxsackievirus A16 (CV-A16), although other emerging strains such as CV-A6, CV-A8, and CV-A10 are increasingly implicated in outbreaks, raising concerns about co-infections and genomic recombination.142,143
Currently licensed whole-virus, inactivated EV-A71 vaccines have shown high efficacy, achieving over 90% protection in clinical trials, particularly against EV-A71-associated disease and neurological complications.144,145 However, they face challenges: scalability concerns in manufacturing centers, potential for virus escape, limited durability, and lack of cross-protection against non–EV-A71 strains.143,144
To overcome these limitations, epitope-based and peptide vaccine strategies are being pursued. One promising target is the EV-A71 SP70 neutralizing epitope. In mouse models, a diphtheria toxoid conjugate of this epitope provided approximately 80% passive protection and elicited cross-neutralizing antibodies against multiple EV-A71 sub-genotypes, including B2, B5, C2, and C4. 146 While displaying the SP70 epitope on an adenovirus type 3 vector induced strong humoral responses and provided approximately 70% protection in mice, it was slightly less effective than a diphtheria toxoid conjugate. To improve efficacy, researchers have developed multi-epitope constructs. For instance, the fusion protein MTLNE, which combines three EV-A71 neutralizing epitopes (SP55, SP70, and SP28), successfully induced specific IgG responses in mice, demonstrating the promise of multi-epitope vaccines. 146
More advanced strategies are also emerging, such as incorporating conserved epitopes into VLP platforms to achieve broader protection across different enterovirus serotypes. Additionally, mRNA-based vaccines are under exploration, offering rapid design flexibility and the potential to cover multiple HFMD-associated enteroviruses. 147
Overall, while current inactivated EV-A71 vaccines have proven effective against EV-A71 infection, their limitations highlight the need for next-generation approaches. Epitope-based, multi-epitope, VLP, and mRNA strategies offer promising avenues to achieve broader, longer-lasting, and more scalable protection against HFMD.
Epstein–Barr virus
Epstein–Barr Virus (EBV) is associated with multiple human malignancies across healthy and immunocompromised populations. While prophylactic vaccines offer promise, no candidate has yet achieved sufficient efficacy against initial EBV infection or asymptomatic carriage. An effective EBV vaccine should ideally prevent primary infection or, at the very least, reduce viral load and transmission. 148 A recombinant gp350 subunit vaccine demonstrated safety and immunogenicity in phase I/II trials and was able to reduce the incidence of infectious mononucleosis, but failed to prevent asymptomatic EBV infection. Beyond glycoprotein 350 (gp350), other glycoproteins such as glycoproteinsH/glycoproteinsL (gH/gL), glycoproteins 42 (gp42), and gB are also important for viral entry and are increasingly being incorporated into multivalent vaccine platforms. 148 For example, a recent study demonstrated that a nanoparticle-based vaccine presenting multiple EBV glycoproteins elicited robust neutralizing antibody responses and protected against EBV challenge in humanized mice. 148
Latency-associated antigens, including Epstein–Barr nuclear antigen 1 (EBNA1), Latent membrane protein 1 (LMP1), and Latent membrane protein 2 (LMP2), are being explored for therapeutic and prophylactic vaccines due to their consistent expression in EBV-associated malignancies. EBNA1, in particular, is expressed in all EBV-infected proliferating cells and elicits both CD4+ and CD8+ T-cell responses. 149 A synthetic polypeptide-based vaccine containing multiple EBNA1 and LMP-derived epitopes has shown promise in inducing strong cellular immune responses and reducing tumor growth in animal models. 149
Recent strategies highlight the importance of mucosal immunity, as EBV primarily enters via oral and oropharyngeal mucosa, and suggest that vaccines inducing local IgA and tissue-resident T cells may provide better protection. 150 DNA prime–protein boost approaches are being explored to enhance both humoral and cellular responses, while VLP vaccines can present multiple EBV antigens simultaneously, eliciting strong immunity even without adjuvants. 150
Computational approaches have also been applied to design epitope-based EBV vaccines, selecting conserved CD8+ and CD4+ T-cell epitopes as well as B-cell epitopes to maximize population coverage and minimize potential autoimmunity. 151 Immunoinformatic and systems biology approaches have been used to predict and validate peptide vaccines, involving 3D modeling, molecular docking, and molecular dynamics simulations to ensure stable peptide-MHC interactions and identify optimal candidates for further experimental evaluation. 152
Collectively, these diverse strategies, ranging from subunit to epitope-based vaccines, and from humoral to cellular immunity, are designed with the aim of providing broad and effective protection against EBV infection and its associated malignancies.
Herpes simplex virus 1
Herpes simplex virus 1 (HSV-1), a neurotropic alphaherpesvirus, causes ocular infections such as blepharitis, conjunctivitis, and herpetic stromal keratitis. 153 Current antiviral therapies, including acyclovir, can suppress symptoms but do not eliminate latent infections, allowing the virus to persist and reactivate. 153 Clinical HSV vaccine trials have largely focused on recombinant glycoprotein antigens (e.g., gD, gC, gB), aiming to induce neutralizing antibodies. 154 While some formulations elicited immune responses, efficacy has been limited, especially in HSV-1 seropositive individuals.153,154
Innovative vaccines are now being designed with the aim of stimulating potent cellular immunity, particularly multifunctional CD8+ T cells, which can reach immune-privileged sites such as the cornea and trigeminal ganglion (TG), which are critical for controlling HSV-1 latency.153,155 For example, a vaccine comprising three peptide epitopes derived from HSV-1 glycoprotein C, helicase, and tegument proteins elicited strong interferon-γ+ CD8+ T-cell responses capable of penetrating both the cornea and TG.153,155 Similarly, AG 702, a heat shock protein–based vaccine, was evaluated in a phase I dose-escalation study and found to be safe and well-tolerated, although detailed immunogenicity data from later phases remain limited. 156
Recent advances in HSV-1 vaccine development focus on epitope-based strategies to enhance cellular immunity. A promising candidate, ΔgD-2, induces strong ADCC responses and offers full protection in animal models.157,158 Additionally, in silico-designed multiepitope mRNA vaccines targeting HSV proteins like gB and infected cell protein 0( ICP0) have shown high immunogenicity. 159 These findings underscore the potential of peptide- and epitope-based vaccines to provide effective and durable protection against HSV-1 infection.
Hepatitis B virus
HBV infection remains a significant global health burden, leading to chronic liver disease, cirrhosis, and hepatocellular carcinoma (HCC). Although current prophylactic vaccines have reduced HBV incidence, they do not treat established chronic infections or HBV-related malignancies, highlighting the need for effective therapeutic vaccines. 160 Effective viral clearance during acute HBV infection relies on coordinated CD4+ T helper and HBV-specific CTL responses, which are often impaired or exhausted in chronic infection. The multifunctional HBV X protein (HBx) plays a critical role in viral replication and oncogenesis and remains expressed in infected hepatocytes and tumor cells, making it an attractive target for immunotherapy.161,162 Therapeutic vaccine candidates incorporating HBx-derived CTL epitopes combined with universal T helper epitopes such as PADRE have demonstrated enhanced immunogenicity and safety in preclinical models.163,164 VLPs presenting multiple HBx CTL epitopes and PADRE have elicited strong cellular immune and antitumor responses in vivo, surpassing single-epitope vaccines. 165
Hepatitis B surface antigen (HBsAg) remains central to prophylactic vaccine formulations due to its capacity to induce neutralizing antibodies.166,167 Structural studies confirm that the α determinant (residues 121–160) of HBsAg, particularly lysine and cysteine residues within this region, are essential for maintaining conformational epitopes required for effective antibody recognition. Chemical modification or mutation of these residues reduces antigenicity and vaccine efficacy.166,167
Earlier studies explored PLGA-based microencapsulation of synthetic HBsAg peptides (e.g., S121/48), which induced strong and durable anti-HBs IgG1 responses in vivo and sustained T-cell memory for over 30 weeks. 168 HBc-based VLPs engineered to display foreign epitopes, formulated with Montanide ISA 720, have also shown good safety and potent humoral and cellular responses in phase I trials. 169 In addition, in silico–predicted HBc and HBsAg epitopes have been validated by IFN-γ Enzyme-Linked ImmunoSpot (ELISPOT) in chronic and resolved HBV patients, revealing comparable epitope recognition but stronger functional responses in certain HLA subgroups, and identifying three novel immunogenic peptides. 170
Emerging therapeutic vaccine designs increasingly utilize multi-epitope constructs that target HBV-specific CTL, helper T cell, and B-cell responses to overcome immune tolerance and achieve functional cure of chronic infection. These approaches include peptide-based, VLP-based, and nucleic acid vaccines currently under preclinical and early clinical investigation.171,172
Hepatitis C virus
HCV is a positive-sense single-stranded RNA virus from the Flaviviridae family. Chronic HCV infection is a major global health concern, leading to progressive liver disease, cirrhosis, and HCC, particularly in low- and middle-income countries where access to direct-acting antivirals is limited. 173 Despite the availability of effective antiviral therapies, the high cost and risk of reinfection have renewed interest in the development of a prophylactic or therapeutic vaccine. 174
The HCV genome encodes three major structural proteins, E1 and E2, and several nonstructural proteins (NS2, NS3, NS4A, NS4B, NS5A, and NS5B), which play critical roles in viral replication and immune evasion. 175 The high genetic variability of the E2 envelope glycoprotein, especially within its hypervariable region 1 (HVR1), remains a major obstacle to developing broadly neutralizing antibody-based vaccines. 176 Cellular immunity, particularly virus-specific CD4+ and CD8+ T-cell responses, plays a central role in the control and clearance of HCV infection. 177 However, persistent infection is often associated with T-cell exhaustion and impaired effector function, particularly of CD8+ T cells. 177
To overcome this, several vaccine candidates have been designed to stimulate robust and multispecific T-cell responses. 178 One such candidate, IC41, includes four human leukocyte antigen A2 (HLA-A2)-restricted CTL epitopes and three promiscuous CD4+ helper T-cell epitopes formulated with poly-L-arginine. 178 It demonstrated the induction of Th1/Tc1-type immune responses in preclinical and early-phase human trials. 178 Another candidate, VAL-44, is a multiepitope peptide vaccine incorporating sequences from HCV non-structural proteins (NS5A, NS4B, and core). It has been shown to induce stronger CD4+ and CD8+ T-cell responses and higher IFN-γ secretion compared to shorter epitope constructs.179,180 Additionally, the protein 7 (p7) of HCV, an ion channel involved in viral assembly and release, has been explored as a vaccine target. 181 Nanoparticle formulations of peptides derived from p7 of genotypes 1a and 1b demonstrated enhanced immunogenicity and the capacity to induce both CD4+ and CD8+ T-cell responses in murine models. 181
Reverse vaccinology approaches have identified conserved regions in E2 and other structural proteins suitable for synthetic peptide vaccines, capable of eliciting targeted immune responses in preclinical studies. 182 Clinical trials of patient-specific (personalized) peptide vaccines and IC41 in patients resistant to interferon-based therapy demonstrated safety and induction of virus-specific T cell and antibody responses, highlighting their therapeutic potential.183,184 These findings collectively support the design of HCV vaccines that can elicit broad cellular immune responses as a promising strategy toward viral control and prevention.
COVID-19
Coronaviruses are notable pathogens affecting both humans and animals. At the end of 2019, a novel coronavirus was identified in Wuhan, Hubei Province, causing pneumonia. The disease rapidly evolved into a global pandemic. In February 2020, the disease was officially named COVID-19 by the World Health Organization, and the causative virus was named COVID-19. Following infection, SARS-CoV-2 induces both humoral and cellular immune responses, which have been shown to persist for at least 1 year post-infection.185,186 Given the crucial role of T cells in durable immunity, vaccination remains the most promising strategy to control the pandemic, particularly for pre-exposure prophylaxis in the general population. 186
Most licensed vaccines focus on the spike (S) protein to elicit neutralizing antibodies. Although currently available vaccines have shown remarkable success in reducing the burden of infectious diseases, some of them may have limitations in terms of long-lasting or broadly protective immunity. Therefore, peptide-based vaccines are being explored to precisely target specific epitopes from viral proteins and induce tailored T-cell and antibody responses.187,188 CoVac 1, a multi-peptide vaccine containing epitopes from spike, nucleocapsid, membrane, envelope, and open reading frame 8 (ORF8) proteins, adjuvanted with a TLR1/2 agonist, elicited robust multifunctional CD4+ and CD8+ T-cell responses in a phase I trial, surpassing those observed after infection or vaccination with authorized vaccines and remaining effective against variants of concern.189,190 Poly personalized epitopes COVID-19 (poly PEPI-SCoV-2), comprising nine 30-mer peptides from all four structural proteins, induced strong Th1-biased CD4+ and CD8+ T-cell responses across diverse HLA genotypes, with projections suggesting multi-antigenic responses in 98% of individuals.191,192 Together, these multi-epitope vaccines highlight the potential of T-cell–mediated strategies to achieve broad, variant-resistant immunity.190,192
Complementing multi-epitope approaches, studies on single conserved epitopes have shown promise. Conserved S2 peptides can induce cross-neutralizing antibodies and protect against multiple variants, indicating potential for universal vaccine development. Additionally, conserved epitopes from the membrane and nucleocapsid proteins may further enhance T-cell immunity. Nevertheless, caution is necessary, as some conserved spike regions may elicit potentially pathogenic autoantibodies, emphasizing the need for careful epitope selection to balance efficacy and safety in universal vaccine design. 193
In summary, peptide-based vaccines offer a promising strategy to induce broad and durable T-cell immunity against COVID-19, with conserved epitopes providing potential protection across variants, while careful epitope selection is essential to ensure safety.
Limitations
Despite the promising advances outlined in this review, several critical challenges and limitations must be acknowledged to provide a balanced perspective. First, a significant portion of the evidence presented, including many of the innovative delivery platforms and epitope designs, is derived from in silico analyses or preclinical studies in animal models. The translational gap between these findings and clinical efficacy in humans remains substantial, as murine immune responses do not fully recapitulate human immunity. Secondly, the heterogeneity of human immune responses, governed by the extreme polymorphism of HLA alleles, presents a major hurdle for universal peptide vaccine design; an epitope that is immunodominant in one individual may be non-immunogenic in another. Furthermore, the manufacturing and regulatory pathways for complex peptide-based formulations, particularly those involving novel adjuvants or delivery systems like nanoparticles, are more intricate and less established than for conventional vaccines, posing significant hurdles for large-scale production and clinical approval. Therefore, while the field is rapidly evolving, these limitations underscore the need for more robust clinical validation and tailored strategies to overcome biological and technical barriers.
Conclusion and future
In recent decades, extensive efforts have been devoted to developing innovative therapeutic and preventive strategies against viral infections. VLP and mRNA vaccines have been successful in clinical trials because of structural similarity with antigens. In addition, peptide-based vaccines, with a focus on epitope precision and safety, bring new possibilities to fight against infectious diseases. Combining new adjuvant design and delivery technologies may help close the immunogenicity gap that exists now. 194 Their advantages over conventional vaccines include reduced risk of pathogenicity (due to lack of in-host replication), precise antigen specificity, chemical stability, ease of large-scale synthesis, and cost-effectiveness. 194 Immunoinformatics plays a critical role in accelerating vaccine discovery by enabling the rapid prediction of B- and T-cell epitopes, thereby minimizing the time and expense of early-stage development.36,195 Advances in adjuvant and drug delivery technologies, such as nanoparticles, liposomes, or biodegradable polymers, have overcome the traditional shortcomings of peptide vaccines, namely poor in vivo stability and low immunogenicity, while enhancing targeted drug delivery and antigen presentation.36,195 Despite these advancements, no prophylactic peptide-based vaccine against viral infections has reached full market approval to date.196,197 However, several are currently in late-stage clinical trials, and therapeutic peptide vaccines (especially in oncology) have shown promising translational progress.196,197 Notably, multiepitope peptide formulations against viral diseases and personalized peptide vaccines tailored by HLA genotyping show great potential.196,197 Moreover, despite promising preclinical outcomes, peptide-based vaccines still face regulatory challenges, such as ensuring long-term safety, optimizing delivery platforms, and standardizing immune response evaluation criteria. 7 These factors complicate approval processes and require tailored frameworks by regulatory agencies. 7 Looking forward, the next generation of immunotherapeutics will likely hinge on personalized peptide vaccines, fine-tuned to individual HLA haplotypes and immune profiles. 198 This evolution is enabled by artificial intelligence and deep learning, which are able to optimize epitope selection, predict immune responses, and simplify vaccine design, potentially revolutionizing immunotherapy development and reducing reliance on animal testing in the early stages. 198 The future of peptide vaccine development will depend on an integrated strategy that incorporates precise epitope design via immunoinformatics, advanced delivery systems (e.g., nanoparticles, liposomes), potent yet safe adjuvants, and personalization tailored to patient-specific HLA profiles, all enhanced through artificial intelligence-driven computational approaches.198–200 Such a coordinated method holds promise for eliciting robust humoral and cellular immune responses across diverse populations and disease targets.198–200