
Abstract
The spatiotemporal progression of dopamine depletion in Parkinson’s disease (PD) provides a special model for assessing dopaminergic effects on neural systems with differential baseline dopamine levels. This study aims at reviewing cognitive effects of dopaminergic stimulation in PD. While considering dopaminergic drugs (levodopa or dopamine agonists), temporal intervals (acute or chronic) and cognitive domains, we found that empirical evidence was almost focused on acute effects of levodopa on executive functions. The paucity of empirical evidence suggests that no meaningful conclusions can be actually drawn and further research is needed in relation to: (1) other cognitive domains; (2) the acute cognitive effects of dopamine agonists, as compared with levodopa; (3) possible differences between cognitive effects of different dopamine agonists; (4) the cognitive effects of chronic dopaminergic therapies. The latter issue is of particular clinical interest considering that many PD patients present a mild cognitive impairment: is this cognitive feature worsened or improved by the prolonged dopaminergic therapy? In addition to the potential risk of inducing dyskinesia and behavioral side effects such as impulse control disorders, also cognitive effects of prolonged dopaminergic treatments should be taken in account by clinicians in order to anticipate or to delay their prescription to PD patients.
Keywords
Introduction
Parkinson’s disease (PD) is a neurodegenerative disorder characterized by motor (bradykinesia, rigidity and resting tremors) and nonmotor symptoms (cognitive impairment, affective and behavioral disturbances, impairment of the autonomic nervous system) [Chaudhury et al. 2006]. Cognitive impairments may be present at an early stage in newly diagnosed drug-naïve patients [Poletti et al. 2012b], with deficits being most prominent in the domains of executive functions, episodic memory and visuospatial functions [Muslimovic et al. 2005]. Prospective studies showed that up to 75–80% of PD patients may eventually develop dementia during the course of the disease, with akinetic-dominant phenotype, early presence of hallucinations and cognitive impairment being the risk factors [Aarsland et al. 2003; Santangelo et al. 2007].
Considering the severe impact of cognitive impairment on the quality of life of PD patients and their families [Schrag et al. 2000], the investigation of factors that may prevent, improve or worsen cognitive impairment represents an important topic in the management of these patients. In this perspective, dopaminergic drugs, representing the gold-standard therapy for motor symptoms of PD patients, may affect their cognitive status [Cools, 2006]. Although both levodopa and dopamine agonists stimulate dopamine receptors, they have different pharmacokinetic characteristics, with levodopa providing a mainly phasic dopaminergic stimulation and dopamine agonists providing a tonic dopaminergic stimulation [Bonuccelli and Pavese, 2006; Poewe et al. 2010]. Furthermore, different dopamine agonists (e.g. pramipexole, ropinirole, pergolide) have distinct receptor binding and pharmacokinetic characteristics, presenting different affinities for dopamine receptors [Perachon et al. 1999].
This review aims at providing an update of empirical evidence on the cognitive effects of dopaminergic drugs on PD patients. Before presenting and discussing findings of empirical studies, the neuropathological bases of cognitive impairment in PD are presented in the following section.
Neuropathological bases of cognitive dysfunction in PD
PD is primarily caused by loss of dopaminergic neurons in the nigrostriatal pathway, reducing dopamine levels in the striatum [Hughes et al. 1992; Kish et al. 1988]. This dopamine depletion has an impact on the functioning of four frontostriatal networks [Alexander et al. 1986; Yeteran and Pandya, 1991] involved in motor, cognitive, affective and motivational aspects of behavior [Chudasama and Robbins, 2006; Owen, 2004]. Two of these circuits have been mainly investigated and have been related in cognitive deficits of PD patients: the ‘dorsolateral’ circuit including the dorsolateral prefrontal cortex (DLPFC), the striatum (dorsolateral caudate nucleus), the globus pallidus (dorsomedial) and the thalamus; the ‘orbital’ circuit including the orbitofrontal cortex (OFC), the striatum (ventromedial caudate nucleus), the globus pallidus (dorsomedial) and the thalamus. Within each circuit, two loops connect the striatum with the prefrontal cortex (PFC): a direct excitatory loop and an indirect inhibitory loop [Alexander et al. 1986; Yeteran and Pandya, 1991].
Frontostriatal circuits are involved in ‘executive functions’, necessary for an appropriate, contextual goal-directed behavior, allowing us to formulate goals with regard to their consequences, to generate multiple response alternatives, to choose and to initiate appropriate actions, to self-monitor the adequacy and correctness of these actions, to correct and modify them when conditions change and finally to persist in the face of distractions [Miyake and Friedman, 2012].
The impairment of executive functions that characterizes most of PD patients from early disease stages [Muslimovic et al. 2005; Poletti et al. 2012b] is not primarily due to a direct neuropathology of PFC, but to reduced dopaminergic striatal stimulation, disrupting the physiological functioning of frontostriatal circuits. Anatomical and neuropathological evidences suggest that the evolving pattern of executive impairment in PD might be explained by considering the spatiotemporal progression of dopamine depletion within the striatum, and in relation to the terminal distribution of its cortical afferents [Cools, 2006; Owen, 2004]. In the early clinical stages of PD the dopamine depletion is greatest in the foremost dorsolateral extent of the head of the caudate nucleus, an area involved in the ‘dorsolateral’ frontostriatal circuit. Executive functions related to this frontostriatal circuit include functions of attentional control, such as working memory, set-switching and planning, and are usually impaired from the early stages of PD [Sawamoto et al. 2008; Rowe et al. 2008]. In the early clinical stages of PD the orbital frontostriatal circuit and the related executive functions, providing a reward-based control of behavior, are mostly preserved [Poletti et al. 2010]. With the progression of disease, the dopamine depletion impairs also the orbital-frontostriatal circuit, probably resulting in an impairment of related executive functions, although these stages of PD have been scarcely investigated by the neuropsychological point of view [Poletti and Bonuccelli, 2012].
Summarizing, temporal and spatial asymmetries of dopamine depletion and their relation with cognition during the progression of the PD-related neuropathology determine the differential cognitive effects of dopaminergic medication on executive functions in PD. The impairment of executive functions represents the core cognitive feature of PD patients and is clearly related to the nigrostriatal degeneration, as suggested by the correlation between the severity of executive dysfunction and the severity of bradykinesia [Domellof et al. 2011; Poletti et al. 2012b], considered the best motor sign of nigrostriatal degeneration [Vingerhoerts et al. 1997]. Although often subtle, deficits may involve other cognitive functions at an early stage, such as memory, language and visuospatial functions [Muslimovic et al. 2005]: these deficits are probably due not only to the indirect effect of executive dysfunction on them, but also to an early cortical neuropathological involvement of posterior regions [Hosokai et al. 2009; Lyoo et al. 2010; Nobili et al. 2011; Pappatà et al. 2011]. With the neuropathological progression of the disease, the widespread cortical diffusion of Lewy bodies [Braak et al. 2003] produces a more severe cognitive impairment, involving several cognitive functions, and often leading to dementia [Aarsland et al. 2003].
Methods
We performed a systematic review of the literature focusing on studies identified in the electronic databases ISI Web of Knowledge, Medline and PsychInfo and published in English language until August 2012. Keywords used for search were ‘Parkinson’s disease’ combined with a term related to dopaminergic drugs (in alphabetical order: ‘apomorphine’, ‘bromocriptine’, ‘cabergolina’, ‘dopamine agonist’, ‘levodopa’, ‘pergolide’, ‘pramipexole’, ‘ropinirole’, ‘rotigotine’) and a term related to cognitive functioning (in alphabetical order: ‘cognition’, ‘executive functions’, ‘language’, ‘memory’, ‘neuropsychology’, ‘prefrontal functions’, ‘visuospatial functions’, ‘working memory’). Studies identified in electronic databases were distinguished on the basis of their methodology in studies investigating acute cognitive effects versus studies investigating chronic cognitive effects. Studies investigating acute cognitive effects adopt a methodology of comparing cognitive performances in ‘on’ and ‘off’ conditions: ‘on’ condition means that patients take their dopaminergic medication and then are tested, while ‘off’ condition means that patients are tested when they have abstained from dopaminergic medication for a minimum of hours (usually at least 12 hours). Studies investigating chronic cognitive effects adopt a methodology of a longitudinal assessment of patients.
Results
The systematic review of electronic databases identified 22 studies designed to assess the cognitive effects of acute dopaminergic stimulation in PD patients and 3 studies designed to assess the cognitive effects of chronic dopaminergic stimulation.
Acute dopaminergic stimulation
The main empirical findings of the 21 studies investigating cognitive effects of acute dopaminergic stimulation on PD patients are summarized in Table 1. A preliminary survey identified two common characteristics among these studies. First, almost all studies investigated the acute cognitive effects comparing performances of patients ‘on’ and ‘off’ dopaminergic therapies. Second, considering the main role played by the dopaminergic systems on executive functions, almost all studies investigated the cognitive effects of dopaminergic therapies exclusively on them
Studies investigating acute cognitive effects of levodopa and dopamine agonists in early and moderate PD patients.

Legend: ↑, improved cognitive performance; ↓, worsened cognitive performance; HC, healthy controls; de novo, newly diagnosed drug-naïve; PD, Parkinson’s disease; WCST, Wisconsin Card Sorting Test; WPT, weather prediction task.
Results of these studies have to be evaluated considering the spatiotemporal progression of dopamine depletion within the striatum. In the early stages of PD the dopamine depletion is greatest (to a maximum of about 90%) in the most dorsolateral extent of the head of the caudate nucleus, producing a dysfunction of the dorsolateral frontostriatal circuit, while the orbital circuit is almost preserved; only in more advanced stages of the disease the orbital frontostriatal circuit is affected by dopamine depletion. This spatiotemporal difference in dopamine depletion at the striatal level explains why the effect of dopaminergic drugs is not linearly correlated with cognition.
One of the first studies on the effects of levodopa on cognitive functions of PD patients demonstrated the enhancement induced by levodopa on performances in executive tasks of verbal and visuospatial working memory and categorization (Wisconsin Card Sorting Test) [Kulisevsky et al. 1996]. These preliminary findings were subsequently confirmed by a series of studies that adopted several executive tasks, most of all included in the Cambridge Neuropsychological Test Automated Battery, showing that the withdrawal of dopaminergic medication in early PD patients has a detrimental effect on set-switching and working memory [Cools et al. 2001, 2002a, 2003, 2010], which are associated with the dorsolateral frontostriatal circuit, whereas it has a beneficial effect on probabilistic reversal learning, associated with the orbital frontostriatal circuit [Cools et al. 2002b, 2006, 2007]. Because the effects of levodopa depend mainly on its ability to elevate dopamine levels in the striatum [Maruyama et al. 1996], the observed different effects on set-shifting and working memory versus reversal learning are most likely due to effects of dopamine in the dorsal and the ventral striatum, respectively, which are known to be connected to different cortical areas via segregated frontostriatal circuits [Cools et al. 2006]. This double dissociation is evident when directly comparing patients ‘on’ and ‘off’ medication and is in line with the ‘dopamine overdose hypothesis’, first formulated by Gotham and colleagues [Gotham et al. 1986], which suggests that the administration of dopaminergic medication to PD patients may replete dopamine-depleted circuits, but overdose relatively intact ones. Indeed, other recent studies confirmed that in the early stages of PD, the treatment with levodopa has a beneficial effect on DLPFC-related executive functions, including attention, set-shifting, working memory and planning [Beato et al. 2008; Fera et al. 2007; Hanna-Pladdy and Heilman, 2010; Mollion et al. 2003; Molloy et al. 2006; Pascual-Sedano et al. 2008] but has a detrimental effect on OFC-related executive functions, that provide a reward-based control of behavior, as evidenced by poor performances in tasks of decision making under ambiguity and reversal learning [Jahanshahi et al. 2010; Rowe et al. 2008].
In advanced PD, when the dopamine depletion affects also the orbital frontostriatal circuit, levodopa is expected to have beneficial effects also on the executive functions related to this frontostriatal circuit; this prediction is actually not sustained by empirical evidence because no studies assessed OFC-related executive functions in advanced PD patients, probably due to the frequent association with dementia in these later stages, and since severe motor impairment often hampers the neuropsychological assessment and the identification of specific cognitive deficits in these patients [Poletti and Bonuccelli, 2012].
As underlined at the beginning of this section, the majority of studies on the effects of dopaminergic drugs on the cognitive status of PD patients focused on executive prefrontal functions, while very few studies investigated other cognitive functions. The enhancement effect of levodopa involves not only functions that are influenced by executive functions [Martin et al. 2003; Vanderploeg et al. 1994], such as prospective memory and verbal learning [Costa et al. 2008; Mattis et al. 2011], but also other functions poorly influenced by executive functions, such as semantic priming [Anqwin et al. 2009], i.e. the faster recognition of a target word when it is preceded by a related prime word compared with an unrelated word. Interestingly, a recent empirical trend highlighted the influence of the dopaminergic striatal system on the hippocampus and the related episodic memory system [Morcom et al. 2010; Shohamy and Adcock, 2010]; therefore, PD appears to be a good empirical model to be adopted in future studies to investigate the relationship between the dopaminergic system and different memory systems [Edelstyn et al. 2010; Foerde and Shohamy, 2011].
The majority of studies on cognitive effects of dopaminergic drugs used levodopa, providing a phasic stimulation; in recent years some studies have begun to investigate also the cognitive effects of dopamine agonists, providing tonic dopaminergic stimulation. A study found that pergolide, a D1/D2 agonist, had no cognitive effects (on episodic verbal memory and executive functions) on PD patients, similarly to levodopa [Brusa et al. 2005]; the same research team reported that pramipexole, a D2/D3 agonist, produced a significant impairment of short-term verbal memory, attention and executive functions, while levodopa did not, in a group of early/mild PD patients [Brusa et al. 2003]. Differently, a study reported that both pergolide and pramipexole improved performance accuracy on verbal and visuospatial working memory tasks in a sample of newly diagnosed drug-naïve PD patients with low baseline performance [Costa et al. 2009]; finally, a recent study [Drijgers et al. 2012] reported any acute cognitive effect of pramipexole in a sample of 23 pramipexole-naïve PD patients.
Other studies investigated the effects of apomorphine and levodopa on the performances of a group of PD patients in visual–spatial and visual–object working memory tasks, compared with performances during ‘off’ phase [Costa et al. 2003; Muller et al. 2002]: apomorphine worsened reaction times in both visual–spatial and visual–object working memory tasks, while levodopa improved accuracy and reaction times in both visual–spatial and visual–object tasks.
Chronic dopaminergic stimulation
A different issue regards the chronic cognitive effect of dopaminergic drugs on PD patients. While negative effects of levodopa on motor functioning are well known (e.g. dyskinesia [Poewe et al. 2010], it is unclear whether the prolonged chronic therapy with dopaminergic drugs, usually taken for many years, has beneficial (protective), neutral or detrimental effects on the cognitive status of PD patients. Indeed, the systematic review of literature found only three studies that investigated this issue (see Table 2): one study [Kulisevsky et al. 2000] followed 20 de novo PD patients for a period of 24 months of treatment with levodopa (10 patients) or pergolide (10 patients; to these patients levodopa was added after 6 months). Both treatments were associated with significant improvements in motor scores and in all cognitive tasks at the first follow up evaluation (until 18 months after the baseline assessment) but, while improvement in motor scores persisted, improvements in some tasks of executive functioning and of long-term memory were not sustained at the final 24-month examination. Another study [Rektorova et al. 2005] assessed the cognitive functions of 41 PD patients in treatment with levodopa before and after 8 months of an add-on therapy with pramipexole or pergolide: any difference was found between cognitive performance at the baseline and after the therapy with dopamine agonists. A similar finding was reported by a study [Relja and Klepac, 2006] that evaluated a sample of 16 medicated PD patients during 12 months of treatment: patients treated with levodopa alone and patients receiving pramipexole as add-on therapy to levodopa did not cognitively differ at the baseline and at the follow-up neuropsychological assessment.
Studies investigating chronic cognitive effects of levodopa and dopamine agonists in PD. The order is chronological.
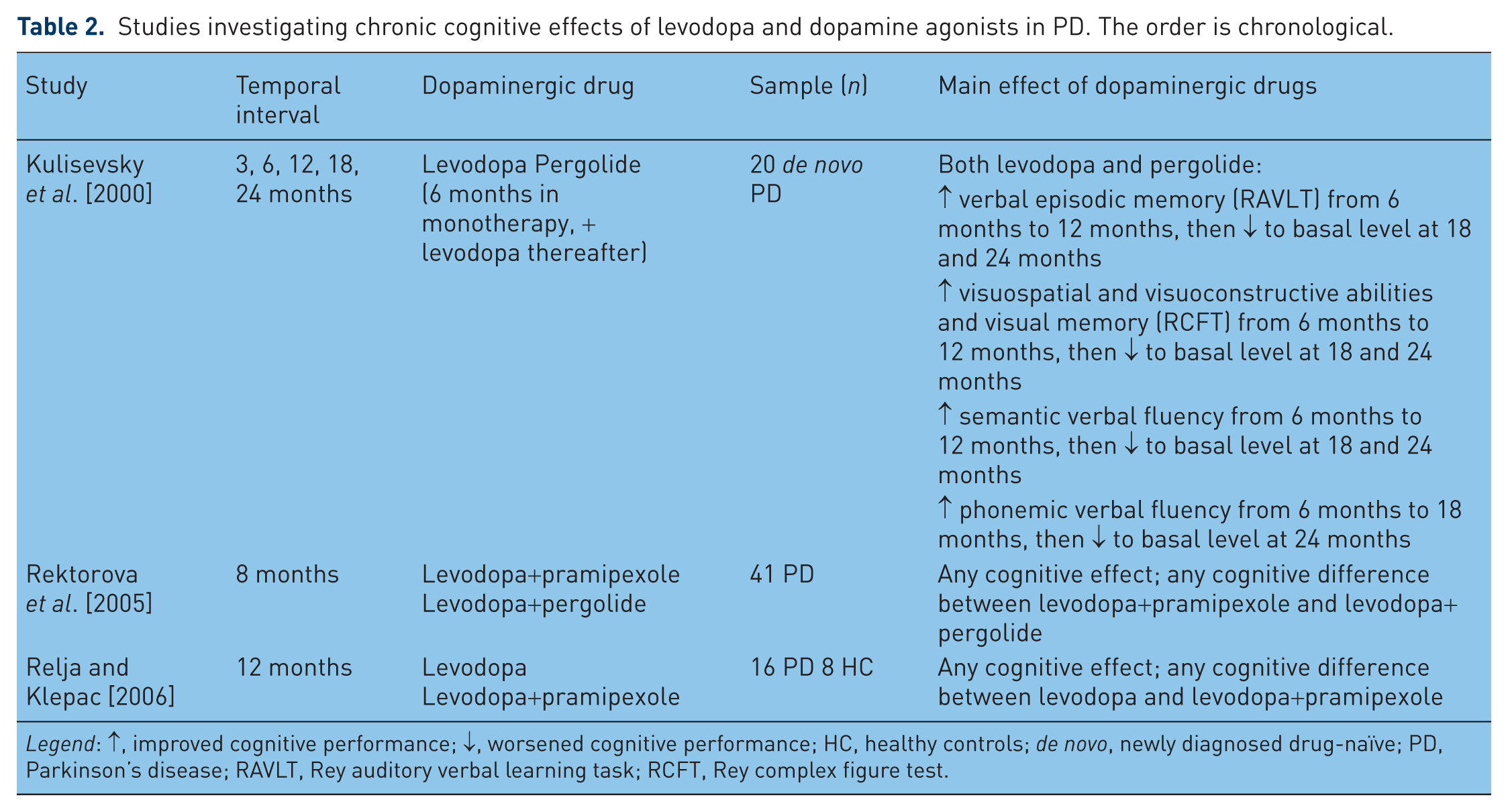
Legend: ↑, improved cognitive performance; ↓, worsened cognitive performance; HC, healthy controls; de novo, newly diagnosed drug-naïve; PD, Parkinson’s disease; RAVLT, Rey auditory verbal learning task; RCFT, Rey complex figure test.
These findings preliminary showed that: (1) chronic dopaminergic stimulation at least do not have negative mid-term effects on cognitive functions of PD patients; (2) levodopa and dopamine agonists do not have differential mid-term effects on cognitive functions of PD patients. In these studies patients were followed only for brief periods (from 6 months to 2 years), while dopaminergic drugs may be taken by PD patients for many years: this suggests that the long-term effect of chronic dopaminergic stimulation with levodopa or dopamine agonists on cognitive functions of PD patients is actually almost unknown.
Discussion
This article aimed at reviewing empirical evidence on the cognitive effects of dopaminergic drugs in PD. The study of cognition in patients with PD is of particular interest because the spatiotemporal progression of dopamine depletion during the course of the disease provides a special model for assessing dopaminergic effects on neural systems with differential baseline dopamine levels. The interaction between degrees of dopamine depletion (dorsolateral versus orbital frontostriatal circuits; left hemisphere versus right hemisphere) and different dopamine replacement therapies may produce different cognitive profiles at different stages of the disease: this complex clinical picture could partially explain why findings of studies on cognitive functions of PD patients are usually heterogeneous also within the same cognitive domain.
Considering different possibilities of empirical investigation of cognitive effects of dopaminergic drugs in PD in relation to drug (levodopa or dopamine agonist), temporal interval (acute or chronic) and cognitive domain, we found that empirical evidence is almost focused on acute effects of levodopa administration on prefrontal executive functions.
Acute cognitive effects: levodopa
Reviewed empirical findings are compatible with neuroanatomical and neurochemical models of dopaminergic frontostriatal systems and their cognitive functions [Chudasama and Robbins, 2006; Owen, 2004]. All of these models commonly propose that, in early PD patients, the withdrawal of dopaminergic medication has a detrimental effect on cognitive functions associated with the dorsolateral loop, and a beneficial effect on the cognitive functions associated with the orbital loop; this pattern has been recently confirmed and better specified by a study that matched behavioral performances of PD patients ‘on’ and ‘off’ dopaminergic drugs and fMRI findings in healthy subjects in a simple selection task [MacDonald et al. 2011]. Findings confirmed that ventral striatum and the related orbital frontostriatal circuit is involved in learning new stimulus–stimulus associations and its functioning is impaired in early PD stages by dopaminergic drugs; on the other hand, dorsal striatum and the related dorsolateral frontostriatal circuit is involved in the assimilation of new and relevant information for the production of more accurate selections, for example shifting attention to more salient stimuli, and its functioning is enhanced in early PD stages by dopaminergic drugs.
This double dissociation involving cognitive effects of dopaminergic drugs is therefore evident when directly comparing patients ‘on’ and ‘off’ dopaminergic medication and was first suggested by the ‘dopamine overdose hypothesis’ [Gotham et al. 1986, 1988], stating that the administration of dopaminergic medication to early PD patients may replete dopamine-depleted circuits (including the dorsal striatum), thus improving performances in tasks related to the dorsolateral loop while ‘overdosing’ relatively intact circuits (including the orbital loop). As levodopa mainly elevates dopamine levels in the striatum [Hornykiewicz, 1974; Maruyama et al. 1996], these differential effects are likely due to opposing effects of levodopa in the dorsal and the ventral striatum, which are connected to different cortical areas via segregated frontostriatal loops [Alexander et al. 1986].
The neurocomputational model of frontostriatal circuitry functioning in PD [Frank et al. 2004] proposed that basal ganglia modulate the selection of actions under consideration in the PFC. Two main projection pathways from the striatum travel up to the cortex through the thalamus via different basal ganglia output structures. The subthalamic nucleus provides a self-adaptive, dynamic control signal that temporarily prevents the execution of any response, depending on decision conflict [Frank, 2006]. The direct frontostriatal ‘orbital’ pathway is excitatory and the indirect frontostriatal ‘orbital’ pathway is inhibitory. Transient changes in dopamine levels that occur during positive and negative feedback loops have opposite effects on the D1 and D2 (dopamine) receptors, which are relatively segregated in the direct and indirect pathways, respectively [Hernandez-Lopez et al. 2000]. Dopamine bursts during positive reinforcement activate the direct pathway and deactivate the indirect pathway, driving learning so that reinforced responses are subsequently facilitated. Conversely, decreases in dopamine result in negative feedback, or deactivation, of the direct pathway and activation of the indirect pathway. Thus, unreinforced responses are subsequently suppressed or avoided. This model predicts a stronger processing of positive rewards in medicated PD patients, since levodopa increases dopaminergic bursts and facilitates an excitatory activity in the direct pathway of the cortico-striato-thalamo-cortical loops. Otherwise, medicated PD patients should show a decreased ability to learn through the mechanism of reward omission. This is because levodopa prevents dips in dopaminergic systems, which disturbs the inhibitory activity of the indirect pathway in the cortico-striato-thalamo-cortical loop. Unmedicated patients should show the opposite pattern, learning sufficiently from negative feedback to avoid harm, while showing impairment in learning from positive reinforcement. This neurocomputational model has been empirically confirmed by administrating a probabilistic selection task to PD patients ‘on’ and ‘off’ dopaminergic medication [Frank et al. 2007]: levodopa altered the patients’ tendency to learn from positive versus negative outcomes, without modifying conflict-induced slowing.
The tonic/phasic model of dopamine system regulation [Grace, 2000; Goto and Grace, 2005] proposed that the nucleus accumbens (NAcc) is believed to regulate goal-directed behavior because it receives convergent synaptic inputs from limbic structures and the PFC. Thus, the NAcc is located such that contextual information from the hippocampus and emotional information from the amygdala, could be integrated with actions programmed in the PFC [Grace, 2000]. Electrophysiological experiments in rats showed that tonic and phasic dopamine release selectively modulates hippocampal and prefrontal cortical inputs through the D1 and D2 receptors, respectively. In addition, D1 activation and D2 inactivation in the NAcc produces behaviorally selective effects (learning versus set-shifting of the response strategy) that correspond to specific afferents. These results suggest that the dynamics of dopamine release regulate the balance between the limbic and cortical drives through activation and inactivation of specific dopamine receptor subtypes in the NAcc, and this regulates goal-directed behavior [Goto and Grace, 2005].
These results are also consistent with empirical results on the detrimental effects of dopaminergic medication on reversal learning in patients with mild PD [Cools et al. 2006], as described in the inverted U-shape model of Cools [Cools, 2006] describing differential effects of dopaminergic drugs on functions of the orbital and of the dorsolateral prefrontal circuits along the PD disease progression.
A subsequent fMRI study [Cools et al. 2007] clarified the neural mechanism underlying this impaired reversal learning caused by dopaminergic therapy in PD patients: PD patients who were ‘on’ or ‘off’ levodopa medication had their brain activity measured by fMRI while performing a probabilistic reversal learning task able to activate the ventral striatum and the orbital frontostriatal circuit. fMRI data showed a role of the NAcc in the dopaminergic modulation of reversal learning in patients with mild PD. Reversal learning was accompanied by an increased NAcc activity only when patients were ‘off’ their dopaminergic therapy. Upon resuming therapy, reversal learning was disrupted due to changes in the functioning of the NAcc. Further studies are necessary to address the pharmacological mechanisms underlying the medication-induced reversal impairment; in particular, studies in patients with severe PD accompanied by a loss of dopamine in the NAcc, will reveal whether the levodopa-induced deficits in patients with mild PD depend on the level of dopamine depletion in the NAcc. Whereas other accounts of the medication-induced impairment do not require the NAcc to be intact [Frank et al. 2004], it is possible that the impairment could be abolished during progression of the disease. Therefore, in an overdosed orbital loop, the dopaminergic replacement therapy prevents the dips in those dopaminergic systems that support the ‘no go’ learning through the indirect pathway of the cortico-striato-thalamo-cortical loop. This phenomenon likely causes dysfunctional reward processing, which impairs learning from reward omission [Frank et al. 2007]. Moreover, considering that the phasic-acting levodopa needed to restore dopaminergic bursts effaces dopaminergic dips during reinforcement learning, while tonic-acting dopamine agonists should impair both dopaminergic bursts and dips, the question remains as to whether levodopa and dopamine agonists have different effects on reinforcement learning.
Acute cognitive effects: dopamine agonists
Few studies were specifically designed to assess acute cognitive effects of dopamine agonists in comparison with levodopa and between different dopamine agonists. As regards pergolide and pramipexole, their positive effect on working memory performances of de novo PD patients [Costa et al. 2009] is in line with the inverted U-shape curve model [Cools, 2006], stating that dopaminergic stimulation in early disease stages replaces the functioning of the dorsolateral frontostriatal circuit, primary involved in working memory; indeed, dopamine agonists had a more beneficial effect in those patients with lower baseline performances, indirectly indicating the presence of a more severe nigrostriatal damage. As regards different findings between cognitive effects of pergolide (neutral) and pramipexole (detrimental) in early medicated patients [Brusa et al. 2003, 2005], considering that patients had similar age and disease duration in the two studies (58 versus 57 years; 2.6 versus 2.5 years of disease duration), the different characteristics (respectively D1/D2 versus D2/D3 agonist) of these drugs probably may have played a role. Moreover, the longer mean disease duration (5 years) of patients evaluated by Drijgers and colleagues [Drijgers et al. 2012], in comparison with previous studies, could partially explain the finding of a neutral effect of pramipexole.
Overall, it could be concluded that whereas the acute effects of levodopa on cognitive functions at different stages of PD seem to be established and well described by the inverted U-shape curve model [Cools, 2006], no meaningful conclusions can be drawn at this time in relation to the acute effects of dopamine agonists on cognition, as compared with levodopa, and the differential effects of different dopamine agonists on cognition. However, dopaminergic receptors are differently represented in the human brain [Bonuccelli et al. 2009], are differently involved by phasic and tonic stimulation [Deleu et al. 2012] and are differently involved in cognition [Takahashi et al. 2012]; considering that different dopamine agonists have different effects on dopamine receptors, with ergolines (bromocriptine, pergolide, lisuride and cabergoline) stimulating D1 and D2 receptors and nonergolines (pramipexole, ropinirole and rotigotine) stimulating D2 and D3 receptors [Bonuccelli et al. 2009] at least different categories of dopamine agonists (ergolines versus nonergolines, i.e. D1/D2 versus D2/D3 agonists) are deemed to have probably different cognitive effects on PD patients, that have to be investigated in future studies.
Chronic cognitive effects
Whereas the acute effects of levodopa on prefrontal executive functions at different stages of PD, especially at early stages, seem to be established and well described by current models of dopaminergic systems, no meaningful conclusions can be drawn and further empirical research is needed in relation to the cognitive effects of prolonged dopaminergic therapies. This issue is of particular clinical interest considering that since the time of clinical diagnosis of PD many patients present a mild cognitive impairment: is this cognitive feature worsened or improved by the prolonged dopaminergic therapy? In addition to the potential risk of inducing dyskinesia and behavioral side effects such as impulse control disorders [Weintraub et al. 2010], also cognitive effects of prolonged dopaminergic treatments should be taken into account by clinicians in order to anticipate or to delay their prescription to PD patients, possibly adopting other drugs with possible effects of neuroprotection and cognitive enhancement, as the selective monoamine oxidase type-B inhibitor rasagiline [Elmer et al. 2006; Hanagasi et al. 2011; Jenner and Langston, 2011].
Future directions
In addition to the clinical issues delineated previously, other issues should be investigated in future studies.
First, although dopamine systems are mainly involved in prefrontal executive functions and the main cognitive effects of dopaminergic drugs are expected to involve them, other cognitive domains have to be investigated from this perspective even if cognitive effects are probably more subtle and difficult to be identified; for example, preliminary empirical evidence suggests that dopaminergic systems are involved, at least with a modulatory effect, in episodic memory [Shohamy and Adcock, 2010], but any conclusion can be actually drawn. Also within the domain of prefrontal executive functions more studies are needed: indeed most studies focused on functions of the orbital and of the dorsolateral frontostriatal circuits, while functions of the ‘anterior cingulate’ frontostriatal circuit (including the anterior cingulated cortex [ACC], the striatum [ventromedial caudate nucleus, ventral putamen] the nucleus accumbens, the olfactory tubercle, the globus pallidus [rostromedial] and the thalamus) have been scarcely investigated in PD: this circuit has been involved in motivated behavior, considering that its damages clinically result in apathetic syndromes [Bonelli and Cummings, 2007]. Apathy is a common neuropsychiatric feature also in PD patients [Starkstein et al. 2009] and has been associated with cingulate anatomic reductions and functional deficits [Benoit and Robert, 2011; Kostic and Filippi, 2011] and with executive impairment [Poletti et al. 2012a], but the role of nigrostriatal dopaminergic deficit on apathy in PD and the potential role played by dopaminergic drugs are actually almost unknown and deserve further empirical investigation.
Second, in addition to the main dopaminergic dysfunction, other neurotransmitters are dysfunctional with different degrees in PD, including acetylcholine, serotonin and norepinephrine [Baloyannis et al. 2006; Bohnen et al. 2006; Guttman et al. 2007], although their role in cognitive dysfunction is partially unknown [Calabresi et al. 2006; Marsh et al. 2009; Scholtissen et al. 2006] and deserves further empirical investigation.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.
Conflict of interest statement
Ubaldo Bonuccelli has been on advisory boards for GlaxoSmithKline, Lundbeck, Novartis and UCB, received honoraria for speeches at meetings from Boehringer Ingelheim, GlaxoSmithKline, Novartis, grants from the Regione Toscana Health Authority and intellectual property rights from Sperling and Kupfer for a book authorship. Michele Poletti has no conflicts of interest to declare.