
Abstract
Summary
Objective
Perineural invasion of cholangiocarcinoma happens in the early stage of the disease but is often not recognized until its later stages. Research about the behaviour and mechanism of perineural invasion by cholangiocarcinoma is urgently needed for a useful new model. The aim of this work is to establish a novel model to address the problem.
Design
Neural cells and cholangiocarcinoma cells were co-cultured to mimic the neurotropic invasion of cholangiocarcinoma.
Setting
Human embryonic stem cells were induced to form neural cells by glial cell-derived neurotropic factor and retinoic acid; neural cells and cholangiocarcinoma cells were co-cultured in Transwell chamber.
Participants
Human embryonic stem cells and cholangiocarcinoma cells were applied.
Main outcome measures
Paired t-test was used to compare the counts of penetrating cholangiocarcinoma cells in co-culture and control group.
Results
Formation of neurospheres and neural-like cells were observed following induction at 24 and 48 h, respectively; synapses were viewed to protrude from neural-like cell bodies after incubation for 96 h. Forty-eight hours after incubation, immunocytochemical staining of the cells showed that synaptophysin and glial fibrillary acidic protein were expressed in the neuron-like cells and gliocytes-like cells, respectively. The cholangiocarcinoma cells that had penetrated through the Matrigel/polyethylene terephthalate membrane from the upper chamber to the lower chamber of the Transwell in the co-culture group were significantly more numerous than those in the control group (68 ± 8.3/field versus 46 ± 5.7/field, P < 0.05).
Conclusion
The novel model is a valuable tool to study the perineural invasion of cholangiocarcinoma.
Introduction
Cholangiocarcinoma has a poor prognosis; even with radical surgery, the mean survival time of patients is 17 months. 1 One of the critical factors that affects postsurgical prognosis is perineural invasion. A previous study showed that carcinomatous perineural invasion occurred in 66% of patients and gravely affected prognosis. 1 The mechanism underlying this invasion remains to be revealed. Though models have been developed to study drug effects on cholangiocarcinoma in vivo and in vitro, there are few models available to study perineural invasion.2–4 We therefore attempted to establish an in vitro model to investigate the interaction between human neural cells and cholangiocarcinoma cells. In our model we observed the perineural invasion of cholangiocarcinoma cells and their neurotropic growth characteristics.
Materials and methods
The embryonic stem cells (ESCs) were obtained from Sciencell Research Laboratories (San Diego, CA, USA); human cholangiocarcinoma cell line QBC939 was a gift from Prof Shu-Guang Wang (the Third Military Medical University, Chongqing, China). Fetal bovine serum (FBS) and Dulbecco's Modified Eagle's Medium (DMEM) and RPMI Media 1640 were from Life Technologies (Shanghai, China). Glial cell-derived neurotropic factor (GDNF) was obtained from Yuan Tai Biotechnologies (Hunan, China); antibodies for synaptophysin (SYN) and glial fibrillary acidic protein (GFAP) were brought from Maixin Bio (Shanghai, China). Transwell cell culture plates were obtained from Corning Life Sciences – Axygen Inc. (Union City, CA, USA).
Induction of ESCs
The ESCs were cultured in six-well plates with RPMI media 1640 containing 10% FBS, 25 μg/L GDNF and 25 μg/L retinoic acid, at 37°C in an incubator with 5% CO2. After 48 h incubation neuron-like cells were observed under microscopy.
Immunocytochemical staining
Well-developed neuron-like cells were chosen and mounted on slides. After fixation, cells were washed with phosphate-buffered saline (PBS) buffer three times followed by incubation with 3% H2O2 for 10 min, washed once with PBS and blocked with goat serum, then anti-SYN and anti-GFAP antibodies were added, and incubation was carried out overnight at 4°C. After washing three times, the second antibody was added and incubated for 30 min at room temperature. Following washing another time, 3,3′-diaminobenzidine (DAB) was added for staining. Then the slides were mounted with neutral resin for microscopic examination and a location with yellow-brown staining was considered to be a positive site.
Establishment of co-culture model
Suspensions (5 × 104/mL) of human cholangiocarcinoma cells, prepared with RPMI supplemented with 1% bovine serum albumin, were added to the upper chamber, 2 mL of neuron-like cells (5 × 104/mL) in DMEM with 10% FBS were added to the lower chamber of the Transwell and placed in an incubator with 5% CO2 at 37°C for 24 and 48 h, respectively. Medium with 10% FBS but without cells was added to the control groups in the lower chambers. Following staining with haematoxylin, cells were counted under the microscope.
Statistical analysis
SPSS 15.0 software was used in statistical analysis and data were shown by mean-variance (x ± s). Paired t-test was used to compare the counts of the Matrigel/polyethylene terephthalate (PET)-penetrating cells found in the co-culture of human neural and cholangiocarcinoma cells with those of the controls. P < 0.05 was considered statistically significant.
Results
Induction of ESC
After induction with GDNF and retinoic acid for 24 h, human ESCs became neurosphere aggregations with high refraction. Formation of neuron-like cells was observed following incubation for another 24 h. The neuron-like cells appeared like irregular polygons and gliocyte-like cells were irregularly oval. Synapses protruded from the neuron-like cell bodies only after incubation for 96 h (Figure 1).
Glial cell-derived neurotropic factor and retinoic acid-induced human embryonic stem cells are shown to become neuron-like cells and gliocyte-like cells. Formation of neurospheres (a) and neural-like cells (b) were observed following 24 and 48 h, respectively, after induction (×800). The synapses were seen to protrude from neural-like cell bodies after incubation for 96 h (c) (×400)
After incubation for 48 h, immunocytochemical staining of the cells showed that SYN and GFAP were expressed in the neuron-like cells and gliocytes-like cells, respectively (Figure 2).
After incubation with glial cell-derived neurotropic factor and retinoic acid for 48 h, immunocytochemical staining of the cells showed that synaptophysin (a) and glial fibrillary acidic protein (b) were expressed in the neuron-like cells and gliocytes-like cells, respectively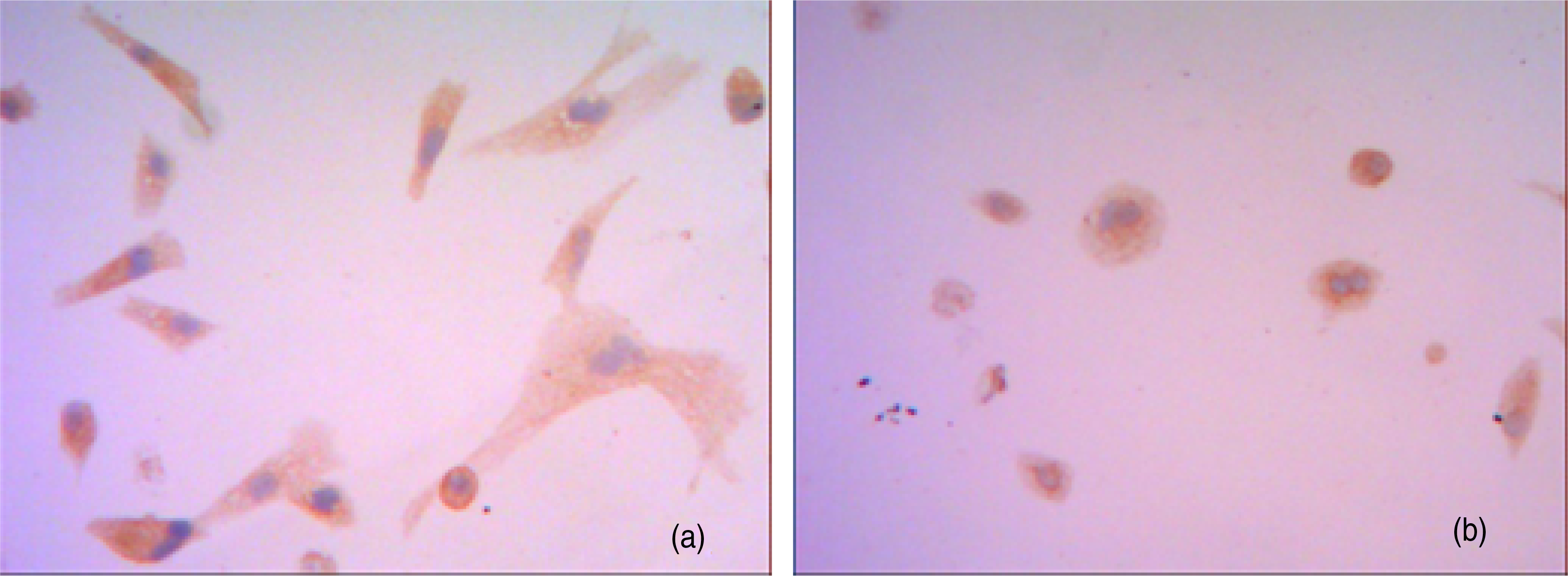
Neural invasion and neurotropic growth of cholangiocarcinoma cells
The cholangiocarcinoma cells in both the co-culture and control groups penetrated through Matrigel/PET membrane. After incubation for 48 h, in the co-culture group cholangiocarcinoma cells aggregated and grew by adhering to neuron-like cells and their synapses, forming cholangiocarcinoma cell masses; some of the cancer cells even penetrated through neural cell membranes and grew inside the cell bodies (Figure 3). The cholangiocarcinoma cells that had penetrated through Matrigel/PET membrane from the upper chamber to the lower chamber of the Transwell in the co-culture group (Figure 3a) were significantly more numerous than those in the control group (Figure 3b) (68 ± 8.3/field versus 46 ± 5.7/field, P < 0.05).
After co-culture for 48 h, cholangiocarcinoma cells in both co-culture group (a) and control group (b) penetrated through Matrigel/polyethylene terephthalate membrane and entered the Transwell lower chambers (×200). The cholangiocarcinoma cells (red arrow) penetrated through neuron-like cell membranes (blue arrow) in the co-culture group and showed characteristics of neurotropic growth (yellow arrow) (a)
Discussion
We successfully established a novel model to study the behaviour and mechanism of perineural invasion of cholangiocarcinoma cells by co-culturing cholangiocarcinoma cells and neural-like cells.
Immunocytochemical staining showed that, after incubation with GDNF and retinoic acid for 48 h, SYN and GFAP, markers of neuron cells and gliocytes, respectively, were expressed in the cells transformed from human ESCs, suggesting that the induced cells had neural characteristics. The accumulated data indicate that retinoic acid delicately controls the differentiation of various stem cells through interaction between diffusion (morphogen-like) gradients, and through the set up of signalling limits by its metabolites. 5 As a vitamin A-derived, non-peptidic, small lipophilic molecule, retinoic acid works as ligands for nuclear retinoic acid receptors, converting them from transcriptional repressors to activators. The levels of retinoic acid in embryonic tissues are maintained by the balance between the synthesis and degradation of the acid. A previous study demonstrated that GDNF can mediate the action of retinoic acid-induced neural cell differentiation. 6
In this study, we observed that more cholangiocarcinoma cells penetrated the Matrigel/PET membrane of the Transwell plate from the upper to the lower chamber in the co-culture group containing cholangiocarcinoma and neural-like cells than in the control group containing only cholangiocarcinoma cell. Our study also demonstrated that cholangiocarcinoma cells formed clusters around neural-like cells and some of them even penetrated the membrane of the neural cells. These data showed that the cultured carcinoma cells had neurotropic growth characteristics, suggesting that they mimicked a very important clinical aspect of cholangiocarcinoma. Hence, this novel model will be a valuable tool to study the perineural invasion of cholangiocarcinoma cells.
In recent years, the mechanisms of differentiation, migration and metastasis of cholangicarcinoma cells remain the focus of research. Data have been accumulated, implicating multiple factors and mechnisms in these processes. Narong and Leelawat 7 observed that by activation of the MEK1/2 pathway, basic fibroblast growth factor induced cholangiocarcinoma cell migration. Overexpression of extracellular matrix protein-1 (ECM1), nuclear protein-1 (NUPR1) and arsenate resistance protein 2 was associated with proliferation and motility of cholangiocarcinoma cells.8–10 Muscarinic acetylcholine receptor M3 (mAChR M3), vitamin D3, L1, urokinase plasminogen activator, immunoglobulin G4 (IgG4)-positive cells, fatty acid-binding protein 5, periostin as well as angiotensin II are factors related to cholangiocarcinoma invasion and metastasis.11–18
Perineural invasion occurs frequently in the early stage of cholangiocarcinoma but is only discovered at its late stages; this occurrence is well correlated with postoperative recurrence and poor prognosis. A variety of factors, such as nerve growth factor, neural cell adhesion molecule, matrix metalloproteinase, acetylcholine and transforming growth factor have been identified to influence the invasion. 19 Our model will help confirm the roles of those factors and explore new mechanisms.
Taken together, the novel model of co-culturing cholangiocarcinoma cells and neural-like cells induced from human ESCs may be a useful tool to study the perineural invasion of cholangiocarcinoma.