
Abstract
Drivers of RNA interference are potent for manipulating gene and protein levels, which enable the restoration of dysregulated mRNA expression that is commonly associated with injuries and diseases. This review summarizes the potential of targeting neuroglial cells, using RNA interference, to treat nerve injuries sustained in the central nervous system. In addition, the various methods of delivering these RNA interference effectors will be discussed.
Introduction
Elucidated by Fire et al. in 1998 1 and hailed as the ‘Breakthrough of the year’ in 2002, 2 RNA interference (RNAi) can be employed as a potent mechanism for overwriting DNA-based regulation in cells, thereby enabling manipulation of cellular functions and behaviour. This was a huge revelation at that time because RNAs have always been thought to be inferior to DNAs and only function to convert genetic information into proteins. 2 It turns out that RNAs are more than that and are capable of executing RNAi, which is a natural phenomenon that exists even across various kingdoms (in plants, fungi and animals) 3 for regulating gene transcription and protein translation. In addition, the mechanism of RNAi is well-established and more information regarding this process can be found in these excellent reviews.3–6
Although its role has not been fully explored to date, it is clear that RNAi is rightfully an evolutionarily conserved defence mechanism against molecular invaders, such as viruses and transposable elements (TEs), where foreign double stranded RNA (dsRNA) were often introduced.3,7 More recently, it has also been recognized that RNAi plays an important role in development, 8 such as neurogenesis,9,10 axonal outgrowth11–13 and myelination 14 in the CNS. Given these huge responsibilities in ensuring the smooth progress of such crucial cellular processes, it is almost certain that RNAi is involved in maintaining cell/tissue homeostasis as well. Indeed, using RNAi microarray analysis, studies are revealing that microRNAs (miRNAs), naturally occurring components for regulating RNAi, are commonly dysregulated following nerve injuries6,15,16 and during disease progression. 14 Therefore, with the goal of either enhancing tissue regeneration or reverting the dysregulated nucleic acid levels to normalcy, effectors of RNAi such as small interfering RNAs (siRNAs), miRNAs, short hairpin RNAs (shRNAs) and antisense oligonucleotides (ASOs) are useful as therapeutics for treating deficiencies in the CNS.
Traumatic nerve injuries often lead to prolonged disruption of the functional coherence of the tissue. This phenomenon, besides partly attributed to the lack of regeneration in neurons, also stems from the inhibitive status of the seemingly inconspicuous glial cells. 17 Being the most abundant but underappreciated cell type in the CNS, these supportive cells are starting to rack up interests from researchers in recent years given their crucial roles in maintaining an optimal environment for neurons to function. Correspondingly, there are also increasing evidences that prudent regulation of their gene and protein expressions following traumatic injuries can promote functional recovery. This suggests that glial cells should also be considered as potential treatment candidates.
Here, we review currently known nucleic acid materials that are capable of eliciting RNAi, as well as methods for efficient therapeutic delivery. In addition, the possibility of targeting glial cells for treating CNS nerve injuries will also be discussed.
Drivers of RNAi
RNAi is fundamentally triggered by the presence of short sequences of RNA strands, which may be single or double stranded and contains around 19 to 29 nucleotides, within the cell. Currently, the predominantly known drivers of RNAi are siRNAs, miRNAs, shRNAs and ASOs. These nucleic acid sequences in their mature form can be easily obtained through commercial means and despite being so similar in their size, each of them have their own uniqueness and efficiency in achieving the intended outcome.
SiRNA
SiRNAs form the main arm of RNAi and are, in design, the most potent molecules to elicit a transient single gene target knockdown once it interacts with the cell’s machinery that is responsible for eliciting RNAi.3–6 Functional siRNAs were observed to be within 21 to 23 base pairs in length and can easily be synthesized in vitro by cleaving long dsRNAs into a pool of siRNAs using Dicer. 3 The main mechanism of siRNA in leading to RNAi is gene regulation, where siRNA binds to perfectly complementary segments of messenger RNA (mRNA) to initiate degradation and prevent transcription from occurring. 18
MiRNA
MiRNAs, unlike siRNAs, exist endogenously. They are transcribed by RNA polymerase II and usually consist of 19 to 25 nucleotides. Contrary to siRNAs, miRNAs induce RNAi by mainly repressing translation, 4 although occasional degradation of mRNA to prevent transcription can occur as well. 19 The modulation of protein at the translational level in animals usually stems from their imperfect base-pairing with target mRNAs.20,21 However in plants, their miRNAs generally bind to mRNAs with near-perfect complementarity, which leads to target mRNA degradation instead,22,23 a mechanism that is similar to that displayed by siRNAs. In a sense, the difference in RNAi regulatory mechanisms between siRNAs and miRNAs is determined by the degree of complementarity during the binding to mRNAs. Yet, due to the slight mismatch of miRNAs towards mRNAs, miRNAs have no true target mRNAs and can, therefore, bind to hundreds or even thousands of them, 24 resulting in massive amounts of translational regulation. Since their discovery, 25 we now know that miRNAs govern physiology at an organism level, especially since they are involved in the entire range of cellular processes such as cell proliferation,26,27 differentiation,10,26,28,29 senescence30–32 and even apoptosis.33,34
Plasmids encoding shRNA
The shRNAs are 19–22 base pairs in length, linked by a short loop of 4–11 nucleotides and drive RNAi similarly as siRNAs. 35 A major difference between shRNAs and siRNAs is that shRNAs are encoded by plasmids, and long-term expression can be achieved if the plasmids are introduced into the nucleus through viral means. By incorporating inducible promotors in the transcription vector, the expression of shRNAs can also be turned on and off, allowing RNAi to be regulated as desired.36,37
ASOs
ASOs are synthetic, single-stranded DNAs that are 8–50 nucleotides in length. They can bind to target mRNAs through complementary base-pairing to induce endonuclease-mediated transcript knockdown and subsequently, protein downregulation.38,39 The concept of ASOs was first described in 1978 by Stephenson and Zamecnik, 40 and ASOs were the first generation of biomolecules that were used for disrupting protein expression, albeit in viruses. Further refinement of the synthesis procedures have since allowed potent modifications to be made on the nucleotides, conferring ASOs enhanced pharmacological properties, such as altering pre-mRNA splicing (gene regulation) and blocking mRNA translation (protein regulation).38,41 Certain modifications, such as the use of a phosphorothioate backbone as well as 2′-O-methyl (2′-OMe) and 2′-O-methoxy-ethyl (2′-MOE) oligonucleotides, can also extend their potency beyond functional effects to increase their circulation duration in serum,42,43 increase their resistance to nuclease degradation, 44 increase their hybridization affinity to their target RNA44–46 and even reduce immunostimulatory activities. 47
RNAi mechanism
The mechanism for eliciting RNAi is generally similar for these four classes of small nucleic acids. One notable difference between them, however, is the degree of modification required to be functional.5,48 In their mature form, these nucleic acids will associate with the RNA-induced silencing complex (RISC), which comprises of Argonaute-2 (Ago-2) and Dicer, before binding to their complementary sequences present in the target mRNAs (Figure 1). 5 Through this binding, the mRNAs will not be able to undergo RNA translation as it will thereafter be either degraded or physically hindered from associating with ribosomes.
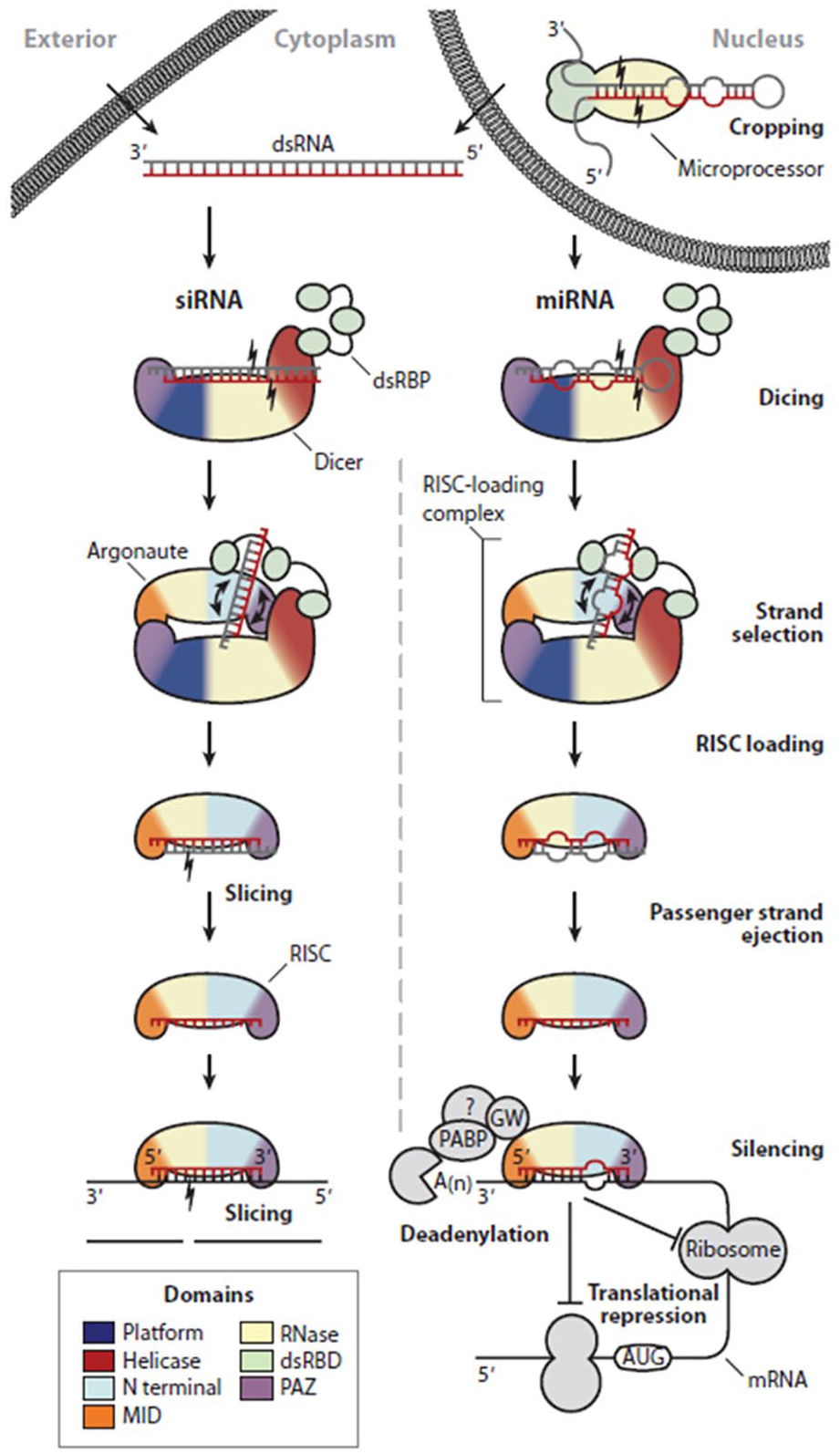
Mechanism of RNAi. RNAi is a general mechanism that siRNAs, miRNAs, shRNAs and ASOs can adopt. The main difference between them lies in the extent of modifications before being functional. Figure depicts a well-established RNAi pathway by siRNAs and miRNAs. Adapted with permission from RC Wilson and Doudna in Annu Rev Biophys, 2013. 5
Delivery methods
While these nucleic acids are known to induce therapeutic effects, the benefits in using these molecules for treating pathologies in the CNS would not be attainable if these biomolecules are unable to reach their intended target environment, that is, the CNS and cytoplasm of cells. Fundamentally, as these nucleic acids possess numerous phosphodiester linkages between each nucleotides, the entire structure tends to be anionic. 49 This indicates that electrostatic attraction is their main form of interaction with other biological materials. In addition, they are also hydrophilic and unable to penetrate effectively into tissue structures within the body. Delivery approaches can, therefore, be designed to either cater to or capitalize on their inherent properties for achieving a therapeutic outcome.
Systemic delivery
There are many strategies for delivering nucleic acids into the body. However, if the delivery region of interest is the CNS and the therapeutics are nucleic acids, then options are limited since the presence of the blood–brain barrier (BBB) and the blood–spinal cord barrier (BSCB) would reduce the efficacy of many conventional (intravenous, intramuscular, oral, transdermal) delivery methods substantially.
Surprisingly, gene/RNAi therapy is currently still in its infant stage of application in the clinics. After conducting a comprehensive search on the list of approved clinical trials related to RNAi within the CNS, it was found that there is currently no approved RNAi therapy for treating CNS-related injuries or diseases. Plasmid deliveries are mostly enabled through viral vectors and are only applicable for neurodegenerative diseases or tumours in the CNS. However, none of them have, to date, reached Phase 3 clinical trial (clinicaltrials.gov, identifier: NCT00876863, NCT00985517, NCT00004080, NCT00004041). To clarify, the physical limitations posed by the BBB and BSCB remain true only under physiologically healthy conditions. In the event of an injury in the CNS, depending on the injury intensity and site, their functional integrity may be compromised. Under these circumstances, there may exist a window for therapeutic interventions and the widely used intravenous delivery route may be the preferred method for delivering nucleic acids before the barriers regenerate. Alternatively, if surgery is unavoidable, then the nucleic acids can also be loaded into a biomaterial, most commonly hydrogels,50–54 and implanted into or placed near the injury site for localized and sustained delivery.
In the event when bypassing the BBB or BSCB is unfavourable, then nucleic acids can also be delivered directly into the CNS through intrathecal infusion. Although not for nerve injuries, this was the strategy employed to deliver ASOs in the first-ever approved treatment of spinal muscular atrophy (SMA). 55 This method of delivery was also used previously in another completed Phase 1 trial for amyotrophic lateral sclerosis (ALS). 56
Besides intrathecal injection, another notable method for the delivery of therapeutics into the CNS is microbubble-assisted focused-ultrasound blood–brain barrier disruption (MB + FUS BBBD). 57 By focusing ultrasonic waves onto acoustically responsive microbubbles injected into the blood vessels, acoustic cavitations are induced. This in turn leads to the transient disruption of the BBB, thereby, allowing intravenously administered drugs to enter the brain parenchyma efficiently. 57 However, it should be noted that optimization of the parameters (microbubble composition, size distribution and concentration as well as ultrasound settings) is required to prevent thermomechanical-induced haemorrhaging 58 and immunoactivation. 59
Tissue-localized delivery
Technically, nucleic acids are usually delivered non-virally in either an encapsulated, complexed or naked form. Encapsulation of these labile nucleic acids protects them from free nucleases and ensures that these drugs are not degraded before they can even be taken up by cells. On the contrary, complexed and naked delivery of nucleic acids 44 will require chemical modifications in order to enhance their resistance to biodegradation. This is highlighted by a previous study which demonstrated that direct administration of both naked siRNAs and ASOs into the brain through intracerebroventricular application led only to the detection of ASOs but not siRNAs. 60
As nucleic acids complexed with a delivery vector are not spared from the harshness of the in vivo milieu, chemical modifications of the former are recommended as well to prevent its degradation. Based on current knowledge, the single most important consideration for the most basic cellular delivery is electrostatic interactions. As both nucleic acids and cellular membrane are similarly charged (anionic), it is not efficient for naked, unmodified nucleic acids to be taken up by cells naturally. 61 Therefore, vectors that are suitable for non-viral delivery are usually cationic (Lipofectamine 2000 reagent,61,62 TransIT-TKO,63,64 polyethylenimine (PEI), 65 etc.). Due to their high net positive charge, these delivery vectors can form complexes with nucleic acids through electrostatic attraction and still remain cationic, which facilitates cellular uptake by enabling the nucleic acid-vehicle complex to attach to the surface of cells for various vesicular transports (endocytosis and/or macropinocytosis) to occur. 66
In addition, the delivery of such complexes can be further augmented through scaffold-mediated approaches. In essence, a scaffolding system can be designed to function as a depot to house the nucleic acids, either naked or complexed with a cationic carrier, thereby ensuring their localization. By concentrating the nucleic acids in a confined region, cells that are present will have an increased chance of encountering them, which promotes their uptake and silencing efficiency. Accordingly, common strategies for retaining the naked and complexed nucleic acids within current conceivable scaffolds include electrostatic interaction,67,68 surface adsorption69–71 and also encapsulation.51,72,73
Besides providing localized delivery, scaffolds can also be tuned to manipulate the extent and even the location of uptake within cells. 74 This can be achieved by altering the physico-chemical properties (elasticity, topology, composition, etc.) of the substrate, which the cells are attached to, thereby regulating cytoskeletal remodelling 75 and subsequently internalization pathways 74 (Figure 2 and Table 1).

Tissue-localized delivery of RNAi. (A) The area around the lateral ventricle of the rat brain 3 h after intracerebroventricular (i.c.v.) injection of fluorescence-labelled siRNA. The i.c.v. injection of 2 μg siRNA and cell detergent i-Fect with 1:4 w/v ratio (d) was only detected but others not (a: 50 μg siRNA dissolved in siRNA suspension buffer without detergent, b: 2 μg siRNA and the other cell detergent DOTAP (1:4 w/v), c: i-Fect only). Adapted with permission from C Senn et al. 60 (B) MiRNAs stimulated extensive neurite ingrowth into fibre-hydrogel scaffolds 2 weeks after spinal cord injury. Neurotrophin-3 (NT-3) and/or miRNAs were treated with PCL fibre-collagen scaffold (a). Neurofilament protein-200 (NF-200) was significantly higher in miRNA-treated groups compared to control group (NT only) (b). Adapted with permission from N Zhang et al. 54
Table comparing the advantages and disadvantages of various nucleic acid delivery strategies. These comparisons provide different perspectives to the choice of delivery method.

CNS and neuroglial cells
The CNS is the part of the nervous system that consists of the brain and spinal cord. The CNS tissue is comprised of two main cell types, namely, the neurons and the neuroglia cells and is enclosed by connective tissue membranes of meninges. 79 The neuroglial cells further consist of astrocytes, microglial and oligodendrocytes (OL). Neurons are the most studied cell type in the nervous system since they have been discovered, in part due to the importance of their role and also probably due to the lack of appreciation of the other neuroglia cells. This notion is rapidly changing recently as more studies are revealing that the neuroglial cells are, in fact, equally as crucial as neurons in maintaining the functionality of the CNS. 80 The highly branched neuroglial cells that are located between neurons have intimate functional relationships with the neurons, providing both mechanical and physiological support.81,82
CNS diseases are a type of neurological disorders caused by various factors including trauma, infections, degeneration, autoimmune diseases and stroke, and can alter and degrade the function or structure of cells and tissues in the CNS.83,84 In particular, the age-dependent neurodegenerative diseases represent a major disease in human because the elderly population has increased in recent years. 85 The major types of neurodegenerative diseases include Alzheimer’s (AD), Parkinson’s (PD), Huntington’s (HD), ALS, multiple sclerosis (MS), Creutzfeldt–Jakob disease (CJD) and others. 86 The pathological characteristics of these neurodegenerative diseases include aggregation and accumulation of specific proteins in the CNS that are associated with neuronal defect in the CNS. In addition, the gliosis that proliferates and activates neuroglial cells is a major hallmark of neurodegenerative diseases.87,88
Among the neuroglial cells, astrocytes are morphologically heterogeneous cells that provide physical support for neurons and mediate the metabolite exchange between the neurons and the blood vessels, including glutamate re-uptake, ionic buffering and water balance.89,90 In addition, astrocytes play an important role in BBB maintenance, neuroinflammatory regulation and the repair of the CNS.91,92 In the case of AD, astrocytes become reactive as a result of generation and deposition of beta-amyloid (Aβ), which in turn leads to decreased glutamate uptake due to reduced expression of uptake transporters, altered energy metabolism and ion homeostasis, increased release of cytokines and inflammatory mediators and disruption of astrocytic networks.91,93 Recently, it was reported that neuroinflammation and ischaemia induced two different types of reactive astrocytes, A1 and A2. A1 astrocytes highly upregulate many classical inflammatory cascade genes to be destructive to synapses, whereas A2 astrocytes upregulate many neurotrophic factors. The A1 astrocyte that is induced by activated inflammatory microglia via the secretory cytokines, including IL-1a, TNF (tumour necrosis factor) and C1q, induces the death of neurons and OLs. 94
Microglia, the resident immune cells of the CNS, have immunological surveillance 95 and defence functions against invading micro-organisms, damaged cells and the debris of apoptotic cells.96–98 Microglia are derived from monocyte progenitor cells and their function is tightly regulated by the CNS microenvironment. 99 Microglia express many pattern-recognition receptors (PRRs) and detect pathogen-associated molecular patterns (PAMPs) or damage–associated molecular patterns (DAMPs), including Toll-like receptors (TLRs) 4 and TLR1/2; their coreceptors; CD14; NOD-like receptors (NLRs); receptors for nucleic acids; C-type lectin receptors (CLRs), such as CLEC7A; and chemokine receptors, such as CX3CR1 and CXCR4. In addition, microglia express immune receptors such as TREM2 (triggering receptor expressed on myeloid cells 2) and multiple receptors for neurotransmitters and neuropeptides released by neurons. 97 Therefore, the microglia are highly responsive to injury or neuroinflammatory disease and are activated under the chronic neurodegenerative diseases of the CNS such as AD, PD, MS and ALS.100,101 For example, proliferation and activation of microglia is a hallmark of AD and the impaired activities and altered microglial responses to Aβ increase the AD pathogenesis and can be harmful to neurons. 102 The over-activated microglia can release cytotoxic factor and inflammatory cytokines. For this reason, inhibiting the activity of microglia appropriately may be an effective way for the treatment of neurodegenerative diseases. 103
OLs are responsible for the formation and maintenance of myelin sheath by neurotrophic support in the CNS.104–106 In order to myelinate properly, OLs have high metabolic activities with oxygen and adenosine triphosphate (ATP) consumption and Reactive oxygen species (ROS) or hydrogen peroxide formation as toxic by-products. Hence, oxidative damage is a common cause to OL disorder or loss under many neuropathological diseases like MS and ischaemia. 107 The damage of OL is observed in neurodegenerative disorders of the CNS, especially chronic demyelinating diseases such as MS, spinal cord injury, AD, PD and ALS. 108 For example, the toxic effects of Aβ accumulation, a major pathological process in AD, induced OL dysfunction and demyelination. 109 In the case of MS, autoimmune system attacks mainly the myelin and OLs. The consequence of such an autoimmune attack is a local demyelination and subsequent axonal loss that is one of the pathological hallmarks of MS. 110
Consequently, the deprivation or dysfunction of these various neuroglial cell populations following traumatic nerve injuries and neurodegenerative diseases hinders recovery given the lack of support, as well as inhibitory effects posed by necrotic cellular debris. 111 As such, the maintenance of spared glial cells followed by their quantitative and steady-state functional restoration is a potential area of focus for facilitating treatment.
Neuroglial cells as potential targets of RNAi for nerve injury treatment
RNAi in astrocytes
Astrocytes constitute the largest proportion of all cells in the mammalian CNS (around 20%–40%). 112 They control neuronal activity and well-being through neurometabolic coupling 113 and help to remove excess neurotransmitters, potassium 114 and glutamate 115 from the extracellular space. Following a traumatic nerve insult, astrocytes become activated or reactive, displaying functional and morphological changes that constitute astrogliosis. 116 Over time, they may also migrate and align to form a glial scar that surrounds the injury site, 117 physically and chemically inhibiting local axonal growth. 118
Studies involving RNAi, conducted in both in vitro and in vivo settings, are thus commonly aimed at circumventing the repercussions caused by this barrier of astrocytes. However, in order to achieve effective gene silencing, it is first imperative to establish a suitable working concentration of RNAi effectors; excessive amounts may be cytotoxic while the opposite may lack potency for an outcome.
Ki et al. conducted a comprehensive testing of the optimal concentration of siRNA for gene silencing in primary cultured rat astrocytes in vitro. Using Lipofectamine 2000 as a transfection reagent, they showed that a siRNA concentration of 20 nM is sufficient to induce efficient uptake (~80%) and RNAi (silencing efficiency ~95%), while preserving the cells’ viability. 119
A similar concentration range was also used in another in vitro study by Li et al., where they investigated the relationship between ephrin-B2 and glial scar formation. Using a custom-made microfluidic platform containing chimeric ventral spinal cord 4.1 (VSC4.1) motoneurons, primary rat astrocytes and rat meningeal fibroblasts (MFb), they demonstrated that the administration of 30 nM of siRNA against ephrin-B2 complexed with SuperFectinTMII reagent (silencing efficiency ~84%) was able to ameliorate the effects of transforming growth factor-β1 (TGF-β1)-induced astroglial-fibrotic scar formation and enhance axonal growth of VSC4.1 motoneurons (Figure 3). 120

RNAi targeting in astrocyte. Silencing of inhibiting factors in astrocytes enhanced axonal growth. (A) Ephrin B2-targeting siRNA restored the motor axon outgrown from VSC4.1 culture towards the axon/scar chamber on the microfluidic platform. (Aa) Diagram depicting the design of the microfluidic platform. (Ab) Photograph of the actual device. (Ac) Schemes depicting motor axon growth towards the axon/scar chamber. In the absence of TGF-β1, astrocytes/MFb coculture allows ingrowth of motor axons into the axon/scar chamber; in the presence of TGF-β1, the coculture forms cell clusters, which resemble astrocyte/fibrotic scar, and inhibits the ingrowth of motor axons. (d, d′) The growth of motoneuron axons slowed down as they approached the axon/scar chamber when CSPGs was present in the axon/scar chamber (e, e′), or when the astrocytes/MFb coculture in the axon/scar chamber was treated with TGF-β1 (h, h′). When the axon/scar chamber was added with astrocytes/MFb coculture (f), or astrocytes/MFb coculture with siRNA (g), VSC4.1 motoneurons extended fine and long axons from the soma chamber and entered the microchannels. (i, i′) Addition of TGF-β1 and ephrin B2-targeting siRNA enhanced length of VSC4.1 axons as compared to TGF-β1 alone. Adapted with permission from Y Li et al. 120 (B) Increased neurite outgrowth on/in conditioned media (CM) from chondroitin polymerizing factor (ChPF)-targeting siRNA-treated Neu7 astrocyte cell line. Cerebellar granule neurons were either grown on a combination of poly-L-lysine (PLL), laminin (LAM) and immobilized CM (a, c) in half DMEM and half neurobasal (NB) + B27 or on a combination of PLL and LAM in half NB + B27 and half CM from the Neu7 cells (b, d). ChPF siRNA attenuated the detrimental effect of CM to neurite outgrowth (e). Adapted with permission from TL Laabs et al. 121
Laabs et al. targeted chondroitin polymerizing factor (ChPF), a key enzyme in the chondroitin sulphate proteoglycan (CSPG) biosynthetic pathway, in Neu7 cells (astrocytic cell line) using plasmids encoding shRNA through nucleofection (a method of transfecting nucleic acids into the cellular nucleus). Although the concentration of plasmids used was not stated, they were able to achieve up to 75% uptake efficiency and ~50% silencing efficiency, resulting in reduced CSPG glycosaminoglycans (GAGs) expression by Neu7 cells. The depletion of these inhibitive polysaccharides subsequently promoted axonal growth (Figure 3). 121
In another recent work, Smith et al. delivered a modified form of siRNA, siRNA three-way junction (siRNA-3WJ), against astrocyte reactivity effectors, namely lipocalin 2 (Lcn2), glial fibrillary acidic protein (GFAP) and vimentin (Vim) into mice astrocytes using Lipofectamine RNAiMAX. Due to the enhanced potency of siRNA-3WJ, they reported ~87%, ~60% and ~65% gene knockdown of Lcn2, GFAP and Vim, respectively, in activated astrocytes using a dosage of only 5 nM. In addition, the uptake efficiency of the astrocytes, both quiescent and activated, was consistently more than 70%. 122 Following this, they proceeded to administer 10 µg of siRNA-3WJ against Lcn2 in mice with a contused spinal cord intralesionally and were able to obtain ~55% reduction in Lcn2 gene expression. Consequently, the contused region showed mitigated levels of Lcn2 protein and was relatively void of cyst formation as compared to the sham control. 122 More examples that are relevant to this review have been compiled in Table 2 and readers are advised to refer to it for completeness. Collectively, all these studies suggest that the modulation of astrogliosis is indeed a potential strategy for enhancing nerve and functional recovery.
A summary of the RNAi-induced gene targets of various glial cells for treating spinal cord injury. Commercially available Lipofectamine is the preferred method for transfecting these cells.
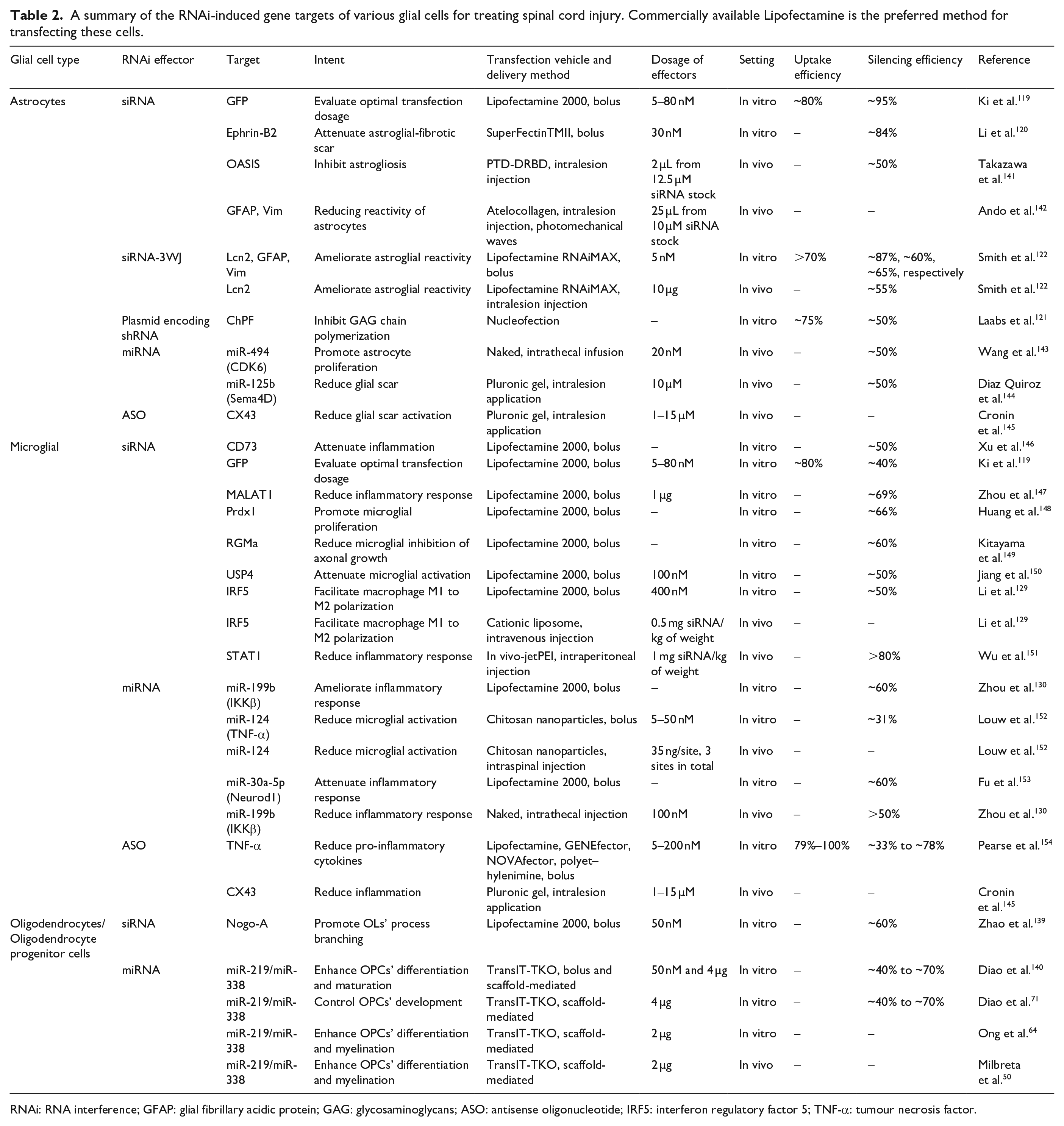
RNAi: RNA interference; GFAP: glial fibrillary acidic protein; GAG: glycosaminoglycans; ASO: antisense oligonucleotide; IRF5: interferon regulatory factor 5; TNF-α: tumour necrosis factor.
RNAi in microglia
Microglia are the resident innate immune cells within the CNS. 123 Following a traumatic injury, microglia become activated and along with monocyte-derived macrophages found in blood, begin releasing ROS and pro-inflammatory cytokines to attract more immune cells to the injured region. 124 Once there, these immune sentinels will mediate the confinement of the lesion, 125 as well as facilitate debris clearance. 126 Studies have also demonstrated that these activities are actually neuroprotective.124,127 While these events appeared to depict rosiness, it is, in reality, very volatile. Specifically, neuroprotective outcomes can easily be replaced with neurotoxic endings if chronic inflammation persists since microglial cells are phenotypically plastic 128 and may react negatively when they are continually exposed to inflammatory signals. Yet, precisely because they are extremely plastic, efforts to encourage nerve repair are possible and usually consist of factors that are capable of skewing them towards a restorative phenotype.
For instance, Li et al. utilized siRNAs to target a transcription factor, interferon regulatory factor 5 (IRF5), known to upregulate genes produced by classically activated macrophages. Using 400 nM of siRNAs against IRF5, they demonstrated that transfected microglial and macrophages isolated from the injured spinal cord had significantly decreased expression of IRF5 (silencing efficiency ~50%) and corresponding amounts of pro-inflammatory cytokines, such as TNF-α, IL-6 and IL-12. Conversely, the anti-inflammatory cytokine IL-10 was enhanced. 129 When these siRNAs against IRF5 were intravenously administered into mice with contusion injuries, similar trends with reduction and enhancement of pro-inflammatory (CD86) and anti-inflammatory (Arg1) microglial, respectively, were observed. Importantly, these observations were associative with limited spinal cord tissue damage and improved locomotor functional recovery (Figure 4). 129

RNAi targeting in microglial. Modulation of macrophage phenotype provided a more conducive environment for tissue regeneration. IRF5 siRNA treatment reduced IRF+ cells (a, b) and CD86 presenting M1 macrophage (a, d). On the other hand, it increased Arginase 1 (Arg1) presenting M2 macrophages (a, c) in the wound of SCI animals. Adapted with permission from J Li et al. 129
In another study, Zhou et al. utilized miR-199b, which they established as a negative regulator of IKKβ, to modulate production of inflammatory cytokines. They demonstrated in vitro in primary rat microglial that overexpression of miR-199b through transfection of miR-199b mimic led to a corresponding suppression of IKKβ gene expression (silencing efficiency ~60%). As IKKβ positively regulates inflammatory cytokines such as TNF-α and IL-1β, the downregulation of IKKβ also resulted in the downregulation of these cytokines. 130 Further administration of 100 nM of miR-199b through intrathecal injection in rats with a compression-induced spinal cord injury similarly attenuated the levels of IKKβ (silencing efficiency > 50%) as well as TNF-α and IL-1β. Subsequent behaviour tests were also performed better by miR-199b-treated rats. 130
While it appears that, in these examples mentioned, along with many more in Table 2, the reduction of pro-inflammatory signals is correlated with enhanced functional outcomes, it is advisable to be most prudent when attempting to manipulate the immune cells’ secretome as a treatment option. Besides the contrasting observations in a very recent work suggesting that prescribed inflammation could promote recovery, 131 it should be recognized that the response of the immune system is far more complex to influence than it seems given its spatiotemporal sensitivity.
RNAi in OLs
OLs are responsible for forming myelin sheath around the axons in the CNS, 132 consequently enabling essential saltatory signal conduction. 133 Following a traumatic nerve injury, OLs become necrotic and disintegrate, leaving behind highly inhibiting myelin components that hinder new myelin formation. Without these sheaths, energy-efficient conduction in axons are impeded, resulting in their eventual degeneration and functional impairment. 134 By targeting OLs and their immature progenitor cells, oligodendrocyte progenitor cells (OPCs), the intended outcome is straightforward: restore the formation of myelin sheaths in the region. Interestingly, while searching the literature, it was observed that viral-based approaches appeared to be the preferred choice135–138 for enacting RNAi in OLs to enhance their myelinating capacity and subsequent extent of recovery from nerve injuries. Despite this trend, it does not imply that non-viral delivery methods are ineffective for transfecting OLs.
Zhao et al. demonstrated that Lipofectamine 2000 could be used to transfect primary OL culture with siRNAs against Nogo-A (50 nM) and achieve a ~60% gene knockdown efficiency. Besides that, they showed that the suppression of Nogo-A in OLs enhanced its process branching, which is a crucial event for initiating myelination. 139
In our lab, we employed a non-viral, scaffold-mediated strategy to transfect OLs. Specifically, we complexed a cocktail of miR-219 and miR-338 (2–4 µg or ~200–400 nM of miRs in total) with a commercial transfection vehicle, TransIT-TKO, and adsorbed these complexes onto polydopamine-coated electrospun fibrous scaffolds before allowing primary rat OPCs to be reverse transfected after they were seeded onto these scaffolds. Through this method of delivery, we were able to obtain ~40% to ~70% of gene silencing efficiency71,140 as well as observe the biological effects of miR-219/miR-338 in enhancing OPC differentiation64,140 and OL myelination.50,64 Furthermore, when we loaded the miR-TKO complex into a three-dimensional (3D) hybrid scaffold containing electrospun fibres and collagen and implanted it into a spinally injured rat, we found that despite the inhibitive microenvironment, miR-219/miR-338 was able to preserve the viability of oligodendroglial lineage cells, promote the extent and rate of their differentiation, and also most importantly, augment their myelinating capacity (Figure 5 and Table 2). 50

RNAi targeting in oligodendrocytes. Promoting oligodendrocyte remyelination will facilitate the functionalization of regenerated axons. (a) Representative images obtained at Week 4 depicting MBP+ tubular structures surrounding NF+ axons. Scale bar represents 5 µm in the normal images and 2 µm in the magnified images. (b) Myelination index obtained at host-implant interface. (c, d) Representative transmission electron microscopy images showing the (c) presence and (d) lack of myelinated axon formation in (c) miR-219/miR-338 and (d) Neg miR groups, respectively. N.S.: not significant (Student’s T-test). Scale bars for the TEM images are labelled with their respective scale bar size. Adapted with permission from U Milbreta et al. 50
Authors’ perspectives and conclusion
In our opinion, the discovery of RNAi and the effectors driving this process is both ground-breaking and exciting. Specifically, the effector nucleic acids are able to provide more utility as compared to normal conventional drugs. For instance, siRNAs can be used to target a single specific gene while miRNAs are often used for targeting multiple genes. On the contrary, shRNAs can be employed to prolong gene regulation with the added benefit of being switchable (on or off). Finally, ASOs can be modified to be used for naked and efficient delivery. However, there are also some downsides pertaining to the use of these nucleic acids for RNAi such as competition with endogenous RNAs, 155 possible activation of innate immune responses 156 and also accidental suppression of off-target genes. 157 In addition, there is always a need to consider the method of intracellular delivery, which usually involves their complexation with a cationic delivery vehicle or encapsulation into nanoparticles.
In the context of spinal cord injury, we feel that miRNAs have the greatest potential for treatment purposes. Recent RNA sequencing of the rodent spinal cord158,159 revealed thousands of dysregulated genes post traumatic injury. Given the ability of miRNAs to regulate numerous genes concurrently, 24 future studies can be conducted to identify the miRNAs required to re-regulate these dysregulated genes back to normalcy and perhaps to map the regulated pathways to the respective cell type.
RNAi holds great potential for treating injuries and diseases, which are frequently associated with dysregulated cellular gene and protein expressions. A major concern, as discussed in this review, is the method to efficiently deliver RNAi drivers into the traumatically injured CNS in order to regulate cellular gene and protein expressions towards an advantageous state, where functional recovery may be attained. Glial cells, being more abundant than neurons, are ideal targets for RNAi therapy. Besides being more amenable than neurons in accepting cationic carrier-complexed small nucleic acids, glial cells are also more proficient in shaping the injured/diseased milieu, through signalling molecules to enhance tissue regrowth. More importantly, besides the previously approved ASO-based therapeutics, with the recent (2018) FDA approval of Onpattro (patisiran), a siRNA-based therapeutic for the treatment of peripheral nerve disease (polyneuropathy) in adult patients, RNAi has become a clinically viable treatment option. Following this landmark acceptance, we can now anticipate that other potent and safe RNAi therapeutics will soon be approved as well. A greater implication of this approval, however, is the motivation for researchers to look towards RNAi as a preferred treatment choice. Specifically, as described, using RNAi to elicit changes in glial cells may provide an avenue to attain a more permissive environment for neuronal regeneration and/or locomotor recovery.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: SY Chew and J Lin would like to acknowledge partial funding support from the Singapore National Research Foundation under its National Medical Research Council Cooperative Basic Research Grant (NMRC-CBRG, NMRC/CBRG/0096/2015) and administered by the Singapore Ministry of Health’s National Medical Research Council; Ministry of Education Tier 1 grant (RG38/19). This research was supported in part by the National Research Foundation (NRF), Republic of Korea (2015K1A1A2032163, 2018K1A4A3A01064257, 2018R1A2B3003446).