
Abstract
Enamel matrix derivative is used to promote periodontal regeneration during the corrective phase of the treatment of periodontal defects. Our main goal was to analyze the bioactivity of different molecular weight fractions of enamel matrix derivative. Enamel matrix derivative, a complex mixture of proteins, was separated into 13 fractions using size-exclusion chromatography and characterized by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and liquid chromatography–electrospray ionization–tandem mass spectrometry. Human periodontal ligament fibroblasts were treated with either enamel matrix derivative or the different fractions. Proliferation and cytokine secretion to the cell culture medium were measured and compared to untreated cells. The liquid chromatography–electrospray ionization–tandem mass spectrometry analyses revealed that the most abundant peptides were amelogenin and leucine-rich amelogenin peptide related. The fractions containing proteins above 20 kDa induced an increase in vascular endothelial growth factor and interleukin-6 secretion, whereas lower molecular weight fractions enhanced proliferation and secretion of interleukin-8 and monocyte chemoattractant protein-1 and reduced interleukin-4 release. The various molecular components in the enamel matrix derivative formulation might contribute to reported effects on tissue regeneration through their influence on vascularization, the immune response, and chemotaxis.
Introduction
Enamel matrix derivative (EMD) was introduced commercially in 1996 as a product (Emdogain; Straumann, Basel, Switzerland) for promoting oral soft and hard tissue regeneration. EMD applied to the root surface during periodontal surgery has been shown to promote periodontal regeneration in experimental1,2 and in clinical studies.3–6 The mechanism of action of EMD is still poorly understood. It is not known whether there are specific components in EMD that promote wound healing or whether the effect is caused by its composition as a whole. EMD contains a complex mixture of proteins generated by the proteolytic processing of a range of alternatively spliced amelogenin gene products. It has also been proposed that EMD also contains growth factors that might be responsible for bioactive effects.7–12 However, Gestrelius et al. 13 reported negative results when they screened EMD using immunoassays for the presence of epidermal growth factor, basic fibroblast growth factor, insulin like growth factor, platelet-derived growth factor, nerve growth factor, and transforming growth factor-β (TGF-β). This issue is further complicated by the fact that EMD stimulates endogenous production of growth factors in different cell lines. 14 Moreover, it has been suggested that specific alternatively spliced gene products or their degradation products in EMD might activate bone morphogenetic protein (BMP) or TGF signaling pathways15–18 and thus mimic the action of certain growth factors.
Some studies have analyzed the effect of different components of EMD on several cell lines. Riksen et al. 19 showed that two isoforms of amelogenin had different effects on messenger RNA (mRNA) expression and protein secretion from osteoblasts. A proteolytic processing product of amelogenin, the tyrosine-rich amelogenin peptide (TRAP), down-regulated osteocalcin while up-regulated osteopontin expression and decreased mineral nodule formation by cementoblasts. 20 Obregon-Whittle et al. 21 found no differences in alkaline phosphatase (ALP) activity in primary human osteoblasts treated with EMD or various fractions of EMD.
The periodontal ligament (PDL) fibroblast has a predominant role in tissue homeostasis, wound healing, and periodontal regeneration.22,23 Cells isolated from the PDL present osteoblast-like properties as evidenced by their capacity to induce mineralization in vitro, express bone sialoprotein, osteocalcin, and ALP.24,25 Fibroblasts contribute to the formation of the extracellular matrix during wound healing through the production of collagen that supports later cell ingrowth. 26 Early in the repair process, cytokines and growth factors secreted by macrophage and fibroblasts initiate a complex signaling network and act as regulators of the inflammatory, proliferative, and remodeling phases of wound healing. 27 Further studies are needed to clarify the complex events underlying periodontal regeneration and especially with regard to the influence of EMD on these events and subsequent PDL fibroblast behavior.
The aim of the present study was to analyze the effect of different molecular weight fractions of EMD on cytokine secretion by periodontal ligament fibroblasts (PDLF).
Materials and methods
EMD fractions
Twenty milligrams of EMD were dissolved in 0.75 mL of 0.125 M formic acid and subjected to size-exclusion chromatography using a 90 × 1.6-cm column of Bio Gel P10 (Bio-Rad, Hemel Hempstead, UK). The column was eluted with 0.125 M formic acid at a flow rate of 0.3 mL min−1. The column eluent was monitored at 280 nm, and 5-mL fractions were collected and lyophilized. A total of 13 fractions (F1–F13) were obtained. F1 was the first fraction to elute and contained the highest molecular weight proteins. Thus, the fractions obtained later had decreasing molecular weights, and F13 was the last fraction to elute and contained the lowest molecular weight proteins or peptides.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis
Fractions were characterized by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) as described by Laemmli. 28 Samples were diluted 1:3 with four times concentrated SDS-PAGE sample loading buffer, heated at 90°C for 2 min and loaded at 10 µL per lane on handcast 15% Tris-HCl gels (Mini-Protean Tetra Cell; Bio-Rad Laboratories, Richmond, CA, USA). Gels were calibrated for molecular weight using 5 µL molecular weight standards diluted 40 times in SDS-PAGE loading buffer (Broad range pre-stained markers; Bio-Rad). After electrophoresis at 200 V, the gels were silver stained and images were taken.
Mass spectrometry
Aliquots of the F3–F12 containing lower molecular weight components identified by SDS-PAGE were subjected to liquid chromatography–electrospray ionization–tandem mass spectrometry (LC-ESI-MS/MS) as previously described. 29 Samples were injected into a nano-ultra high performance liquid chromatography (UHPLC) system (Ultimate 3000 RSLC; Dionex, Sunnyvale, CA, USA) coupled to an ESI-ion trap/Orbitrap (LTQ Orbitrap XL; Thermo Scientific, Bremen, Germany) mass spectrometer. For peptide separation, an Acclaim PepMap 100 column (50 cm × 75 µm) packed with 100 Å C18 3 µm particles (Dionex) was used with a flow rate of 300 nL min−1 and a solvent gradient of 3% B to 35% B in 20 min. Solvent A was 0.1% formic acid, and solvent B was 0.1% formic acid/90% acetonitrile. Survey full scan MS spectra (from m/z 300 to 2000) were acquired in the Orbitrap with the resolution R = 60,000 at m/z 400 after accumulation to a target of 1,000,000 charges in the LTQ. The method used allowed sequential isolation of up to the seven most intense ions for fragmentation on the linear ion trap using collision induced dissociation (CID) at a target value of 10,000 charges. Target ions already selected for MS/MS were dynamically excluded for 60 s. Data were acquired using Xcalibur v2.5.5, and raw files were processed to generate peak list in Mascot generic format (*.mgf) using ProteoWizard release version 3.0.331. Database searches were performed using Mascot in-house version 2.4. to search the following peptides: porcine P173 amelogenin, 30 porcine P56 leucine-rich amelogenin peptide (LRAP), 30 and porcine P73 containing exon 4, 31 assuming the digestion enzyme non-specific, fragment ion mass tolerance of 0.60 Da, parent ion tolerance of 10 ppm and oxidation of methionines, propionamide of cysteines, and acetylation of the protein N-terminus as variable modifications. Scaffold (version Scaffold 4.3.4; Proteome Software Inc., Portland, OR, USA) was used to validate MS/MS-based peptide, and protein identifications were accepted if they could be established at greater than 95.0% probability by the Peptide Prophet algorithm 32 with Scaffold delta-mass correction. Protein identifications were accepted if they could be established at greater than 98.0% probability to achieve a false discovery rate (FDR) less than 1.0%.
Cell cultures
Commercially available human PDL fibroblasts (Catalog number CC-7049, Lonza, Walkersville, MD, USA) 21 were cultured in media (fibroblast growth medium-2 (FGM-2); Lonza, Basel, Switzerland) containing insulin, fibroblast growth factor, gentamicin/amphotericin-B, fetal calf serum and ascorbic acid. Cells were subcultured at 37°C in a humidified atmosphere of 5% CO2 prior to confluence, according to the manufacturer’s instructions. Cells at passage 8 were used for this experiment.
The cultured cells were treated with either EMD (Biora, Malmö, Sweden) (10 µg mL−1) or the various EMD fractions in molar concentrations equivalent to their content in 10 µg mL−1 of whole EMD. Untreated cells were used as controls. Three replicate experiments were conducted for every fraction, EMD, and the controls. Cell culture media were harvested after 1, 3, 7, and 14 days.
Proliferation assay
Cell proliferation was measured using [3H]-thymidine incorporation. Subconfluent cells were treated with either EMD (Biora, Malmö, Sweden) (10 µg mL−1) or the various EMD fractions in molar concentrations equivalent to their content in 10 µg mL−1 of whole EMD. Untreated cells were used as controls. The cells were pulsed with 1 µCi/well of [3H]-thymidine 12 h prior to harvest at 1 and 3 days. The medium was removed and the cells were washed twice with ice-cold phosphate-buffered saline (PBS) and twice with ice-cold 5% trichloroacetic acid to remove unincorporated [3H]-thymidine. The cells were solubilized in 500 µL of 1 M sodium hydroxide, and 400 µL of the solubilized cell solution was transferred to 4 mL of Instagel II Plus liquid scintillation fluid (Perkin Elmer, Applied Biosystems, Foster City, CA, USA) and measured for 3 min in a liquid scintillation counter (Packard 1900 TR).
Protein quantification in cell culture medium
Aliquots of the cell culture medium were concentrated 5-fold using centrifugation filters with 3 kDa nominal cut-off (VWR International, Radnor, PA, USA) according to the manufacturer’s instructions. Multianalyte profiling of the level of cytokines in the concentrated cell culture medium of PDFL was performed on the Luminex-200 system (Luminex Corp., Austin, TX, USA). Acquired fluorescence data were analyzed using the 3.1 xPONENT software (Luminex). The amount of eotaxin, granulocyte colony-stimulating factor (G-CSF), interferon-α2 (IFN-α2), interferon-γ (IFN-γ), interleukin-1β (IL-1β), interleukin-1 receptor antagonist (IL-1ra), interleukin-2 (IL-2), interleukin-4 (IL-4), interleukin-5 (IL-5), interleukin-6 (IL-6), interleukin-7 (IL-7), interleukin-8 (IL-8), interleukin-10 (IL-10), interleukin-12p40 (IL-12p40), interleukin-13 (IL-13), interleukin-15 (IL-15), interleukin-17 (IL-17), interferon-γ induced protein (IP-10), monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-1α (MIP-1α), macrophage inflammatory protein-1β (MIP-1β), regulated upon activation of normal T-cell expressed and secreted (RANTES), soluble interleukin-2 receptor-α (sIL-2Rα), tumor necrosis factor-α (TNF-α), and vascular endothelial growth factor (VEGF) secreted into the culture medium was measured using the human cytokine/chemokine kit (Milliplex human cytokine MPXHCYTO-60k; Millipore Corp., Billerica, MA, USA). All analyses were performed according to the manufacturers’ protocols. The intra-assay and interassay variation for the cytokines analyzed ranged between 3.7 and 15.7%.
Values below the detection limit were set to half of the detection limit. This was done to include these values in further statistical analyses.
Statistical analysis
Statistical comparison between groups and controls was performed using parametric one-way analysis of variance (ANOVA) and post hoc Dunnett’s test with multiple comparisons against the untreated control group. When the normality test was not passed, ANOVA on ranks (Kruskal–Wallis test) and post hoc Dunn’s test were performed (SigmaStat software; Systat, San Jose, CA, USA); p ⩽ 0.05 was considered significant.
Results
Characterization of EMD fractions
The SDS-PAGE showed that the most prominent bands were found between 38 kDa and 5 kDa in the unfractionated EMD (Figure 1). The most intense band found in EMD at 20 kDa corresponded to the well-documented proteolytic processing product comprising residues 1–148 of P173 amelogenin. The fractions F1 and F2 presented a similar protein profile to EMD, although F2 had less stained bands over 38 kDa. F3–F6 had a weaker protein band around 20 kDa, did not contain some of the proteins between 20 and 10 kDa, and contained a band around 5 kDa. F3 had an additional component of 8.8 kDa, which was not observed in F4 and F5. The findings of the mass spectrometric analyses are shown in Figure 2, Table 1, and Supplementary Table S1. Amelogenin P173-related peptides were more predominant in F3–F7 (Figure 2). Fractions F5–F8 presented a higher number of peptides exclusive to amelogenin P173 (Table 1). The quantitative analyses revealed that LRAP related peptides were more abundant in F9–F13 (Figure 2). However, the quantitative analyses of these fractions have to be interpreted cautiously since many sequences attributed to LRAP were also shared with amelogenin P173 (Supplementary Table S1, column labeled “other proteins”). More peptides exclusive to LRAP were found in F5, F6, and F8 (Table 1). LRAP containing exon 4 was not found in any of the fractions analyzed (Table 1 and Figure 2). A shorter TRAP was found in F9 (comprising residues 3–43) and F10 (comprising residues 1–43) (Supplementary Table S1). An interesting finding is that amelogenin is processed into many different peptides, and there is a high variation in cleavage sites (Supplementary Table S1).

SDS-PAGE of EMD and of the different fractions obtained after size-exclusion chromatography. Lane M represents the molecular weight standards with sizes marked on the left (in kDa); Lane EMD represents enamel matrix derivative; Lanes F1–F13 represent EMD fractions, where F1 is the first eluate. Arrows point to different molecular weights.
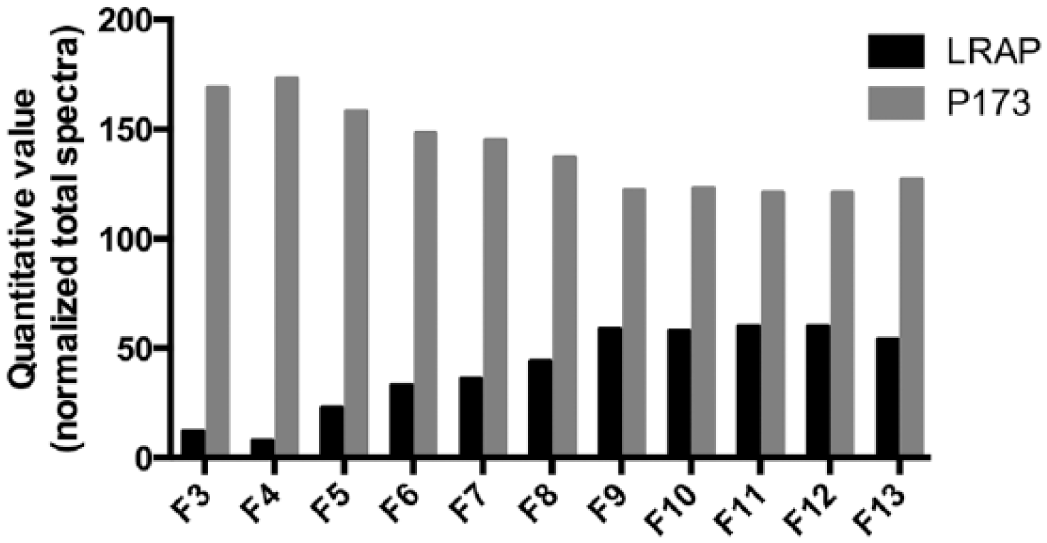
Quantitative analyses by mass spectrometry (LC-ESI-MS/MS)—normalized spectrum count.
Mass spectrometric analysis of fractions 3–7.

F3–F13: fractions 3 to 13. P173: porcine amelogenin sequence of 173 residues; LRAP: porcine leucine-rich amelogenin peptide of 56 residues.
Exclusive unique peptides: number of amino-acid sequences, regardless of any modification that are associated with a protein. Exclusive unique spectra: number of specific spectra associated with a single protein. Sequence coverage (%): percentage of all the amino-acids in the protein sequence that were detected in the sample. Protein identification probability: calculated probability for the protein identification. 33
Cell proliferation
The proliferation rate was increased 2- to 5-fold by lower molecular weight fractions (F8, F9, F10, and F11) at day 1, whereas no changes were found at day 3 (Figure 3).

Effect of EMD and F1–F13 on proliferation of periodontal ligament fibroblasts at day 1 and 3. Cell proliferation was measured as [ 3 H]-thymidine incorporation in counts per minute (CPM). Data presented as median with interquartile ranges (SD) (n = 3).
Effects of EMD and fractions on cytokine secretion
An overview of changes in cytokine levels for the different fractions is given in Table 2. EMD stimulated a 3-fold increase in the concentration of VEGF in cell culture medium at day 1 compared to the control group (p < 0.001) (Figure 4). By day 3, the concentration of VEGF was still elevated (2.5-fold) and by day 7 had increased 660-fold (although not reaching statistical significance) (Figure 4). At day 3, EMD stimulated a 4-fold increase in IL-6 (p < 0.001) secretion and induced a 30% reduction in IL-4 (p = 0.045) secretion compared to the control group (Figures 5 and 6). EMD promoted the secretion of MCP-1 (around 80-fold, p < 0.001) at day 7, although this effect was reversed toward the end of the experiment (Figure 7). EMD also down-regulated IL-8 secretion at day 14 (p = 0.011) (Figure 8).
Cytokine levels in cell culture medium relative to control.

PDL: periodontal ligament; EMD: enamel matrix derivative Emdogain; VEGF: vascular endothelial growth factor; IL-4: interleukin-4; IL-6: interleukin-6; IL-8: interleukin-8; MCP-1: monocyte chemoattractant protein-1.
Cytokines after symbol ↑ mean that they are significantly increased compared to the control group. Cytokines after symbol ↓ mean that they are significantly decreased compared to the control group.
p < 0.05; **p < 0.01; ***p < 0.001.

Mean VEGF concentration (pg mL−1) in cell culture medium from periodontal ligament fibroblasts at day 1, 3, 7, and 14. Data are presented as mean ± standard deviation (SD) (n = 3).

Mean IL-6 concentration (pg mL−1) in cell culture medium from periodontal ligament fibroblasts at day 1, 3, 7, and 14. Data are presented as mean ± standard deviation (SD) (n = 3).

Mean IL-4 concentration (pg mL−1) in cell culture medium from periodontal ligament fibroblasts at day 1, 3, 7, and 14. Data are presented as mean ± standard deviation (SD) (n = 3).
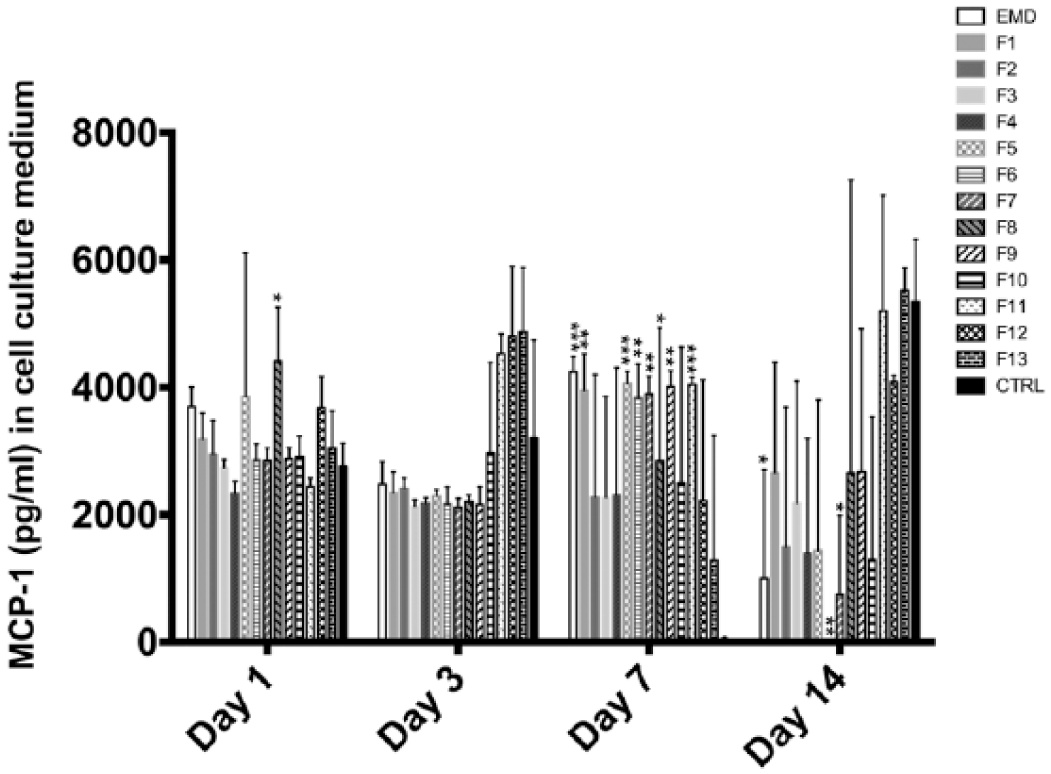
Mean MCP-1 concentration (pg mL−1) in cell culture medium from periodontal ligament fibroblasts at day 1, 3, 7, and 14. Data are presented as mean ± standard deviation (SD) (n = 3).

Mean IL-8 concentration (pg mL−1) in cell culture medium from periodontal ligament fibroblasts at day 1, 3, 7, and 14. Data are presented as mean ± standard deviation (SD) (n = 3).
F1 and F2 down-regulated IL-4 secretion during the first week (p < 0.05), but by the end of the experiment, F2 increased the release of IL-4 compared to the control group (p = 0.032) (Figure 6). F2 also increased the release of VEGF at day 3 (4-fold, p < 0.001) and IL-6 at day 7 (1500-fold, p = 0.033) (Figures 4 and 5).
F3 stimulated the release of VEGF at day 3 (around 7-fold, p < 0.001), and later, it sharply increased VEGF release by 2400-fold at day 7 (p < 0.001) (Figure 4). This effect on VEGF secretion was not seen in lower molecular weight fractions.
At day 7, F5 stimulated the release of MCP-1 80-fold (p = 0.001) and IL-6 1500-fold (p = 0.036) compared to the control group (Figures 5 and 7).
As was the case with EMD and F2, lower molecular weight fractions F7–F13 had an influence on the secretion of chemoattractant cytokines belonging to the chemokine family, such as MCP-1 and IL-8 (Figures 7 and 8). During the first week, lower molecular fractions up-regulated chemokine secretion (F5–F9 and F11) by a similar magnitude as higher molecular fractions: close to an 80-fold increase for MCP-1 and a 500-fold increase for IL-8 (Figures 7 and 8). In contrast, there was a down-regulation of IL-8 and MCP-1 secretion by F5 at day 1 (p = 0.025) and by F4–F7 at day 14 (Figures 7 and 8). The down-regulation of IL-4 seen in higher molecular weight fractions was also observed in lower molecular weights at day 3 (Figure 6).
The secretion of 20 cytokines (eotaxin, G-CSF, IFN-α2, IFN-γ, IL-1β, IL-1ra, IL-2, IL-5, IL-7, IL-10, IL-12p40, IL-13, IL-15, IL-17, IP-10, MIP-1α, MIP-1β, RANTES, sIL-2Ra, TNF-α) out of the 25 cytokines analyzed were consistently below the detection limit for the assay for all treatment groups.
To sum up, the effect on proliferation and the cytokine secretion study cannot be related to any of the specific proteins analyzed in LC-ESI-MS/MS as LRAP- and P173-related peptides were found in all fractions. However, by comparing to the SDS-PAGE gel, it appears that VEGF and IL-6 release was related to EMD components above 20 kDa, since their secretion was only stimulated by fractions containing these molecular weights. On the other hand, lower molecular weight components appear to enhance cell proliferation and secretion of MCP-1 and IL-8 and reduce IL-4 release. Fractions F9 and F10, containing LRAP and truncated forms of TRAP, promoted enhanced proliferation of PDL cells.
Discussion
A systematic analysis of the level of 25 cytokines secreted to the cell culture medium from PDFL after the administration of different fractions of EMD showed that both high and low molecular weight proteins contribute to the bioactive effect of EMD.
Primary human PDL cells were chosen as a model to study the effect of molecular weight fractions of EMD on cytokine secretion as primary cells are more representative of PDL cells in vivo compared to transformed cell lines. 34 The use of cells from one donor allows experimental repeatability since the inclusion of cells from several donors would incur individual variations that might mask any potential effects. Donor variability can explain differential cell behavior in culture conditions.35–37 This issue is further complicated in the study of pleiotropic cytokines with multiple biological activities. Nonetheless, the results of this study should be interpreted with the caveat that they are based on cells provided by one donor. Moreover, in vitro studies lack the complex environment of living tissues.
The SDS-PAGE analyses showed that most proteins contained in the EMD preparation comprised molecular weights between 5 and 25 kDa. This is in agreement with Maycock et al. 38 who also separated EMD using SDS-PAGE. The mass spectrometric analyses revealed the presence of P173- and LRAP-derived peptides in fractions F3–F13. The smaller P56 LRAP and TRAP sequences were, as expected, more frequently found in lower molecular weight fractions. The finding that proteolytic enzymes cleave amelogenin-derived peptides in a wide range of amino-acid sequences (Supplementary Table S1) is in accordance with data presented by Nagano et al. 39 and Ryu et al. 40
Lower molecular fractions (F8, F9, F10, and F11) stimulated cell proliferation, in line with the results from Johnson et al., 7 who found that lower molecular weight EMD components enhanced proliferation of endothelial cells. This effect might be mediated through stimulation of local growth factor secretion 14 or biologically active component(s) in EMD fractions: for example, LRAP whose presence and bioactivity in EMD was demonstrated by Stout et al. 18 EMD did not induce proliferation in the present study. However, the proliferative effect of EMD might manifest at later time points following its administration.41,42
In the present study, higher molecular weight fractions induced a different cytokine secretion profile compared to lower molecular weight proteins. Thus, fractions containing a 20 kDa component enhanced the release of VEGF and IL-6. F5, with relatively few components compared to higher molecular weight fractions, also promoted the secretion of IL-6. Mass spectrometry revealed that this fraction contained peptides related to the P173 amelogenin and LRAP. Therefore, it is not possible to establish which specific component induced IL-6 secretion. On the other hand, this effect was not found with lower molecular weight fractions. Parkar and Tonetti 43 found that EMD administration down-regulated the IL-6 gene, whereas IL-6 production was stimulated in the present study. By contrast, this enhancement of IL-6 secretion was also observed by Lyngstadaas et al. 14 IL-6 is a pleiotropic cytokine that regulates immune response, inflammation, and hematopoiesis. Although IL-6 has been considered a classic inflammatory cytokine, it has also many regenerative and anti-inflammatory activities. 44 IL-6 appears to have a crucial role during wound healing as demonstrated by the fact that IL-6 deficient mice have been shown a delayed skin-wound healing. 45 In addition, F2 and F3 induced VEGF secretion. This is in line with other studies demonstrating an increase in VEGF gene expression 43 or protein release. 46 This observed effect might explain the proliferative activity of amelogenin on microvascular endothelial cells. 7 Classical inflammatory cytokines, for example, IL-1β and IL-6, are released early during the inflammatory response of wound healing and stimulate the production of other cytokines and growth factors, such as VEGF, initiating a cascade of molecular events leading to inflammation, tissue formation, and remodelling. 47
Lower molecular weight fractions (F7–F13) had an effect on IL-4 and chemotactic cytokines (i.e. IL-8 and MCP-1). The chemokines IL-8 and MCP-1 regulate leukocyte migration from blood to tissues and play an important role during wound healing. IL-8 was decreased at day 1 and day 14 by EMD and fractions F4–F8. IL-8 is decreased during wound healing in fetal tissues, and this suggests that the diminished inflammatory response in fetal healing might contribute to a scarless wound healing and regeneration in some tissues.47,48 Nonetheless, IL-8 and MCP-1 secretion were increased at day 7 by several fractions in the present experiment. This increased pattern of MCP-1 and IL-8 expression during the first week has been correlated with leukocyte migration to the wound site in an in situ hybridization study. 49 Early in the healing phase (day 1), IL-8 was correlated with neutrophil migration, but at day 4, IL-8 expression declined. 49 In the same study, the migration of macrophages was observed after strong expression of MCP-1 from day 2 to day 7. Lymphocyte recruitment also correlated in the first 4 days with MCP-1 expression. 49 Therefore, the sequential cytokine expression, leading to activation and inhibition of cytokine release, seems to be an essential aspect of an improved wound healing. IL-4 is a pleiotropic cytokine with an important role in wound healing that induces the production of extracellular matrix components such as type I and type III collagen 50 and stimulates the proliferation of fibroblasts. 51 Surprisingly, IL-4 was down-regulated by almost all fractions. Parkar and Tonetti 43 found that genes related to IL-4 were also down-regulated by the application of EMD to PDL fibroblasts. Further investigations are needed to clarify the relation between EMD and IL-4.
To our knowledge, this is the first report that systematically assesses the secretion of cytokines from primary human PDL fibroblasts after the administration of different fractions of EMD. To try and better understand wound healing mechanisms associated with EMD treatment, it was decided to assess the cytokine expression over a 14-day period since this is the period during which the most important events occur during the early wound healing phase. Furthermore, the retention of Emdogain on the root surface has been demonstrated during a period of 2–4 weeks following its clinical application. This period of time seems to be enough to permit recruitment and recolonization of PDFL over the root surface.2,52,53
Conclusion
Our results indicate that higher molecular components of EMD might be related to an angiogenic effect through the stimulation of VEGF release and modulate wound healing through the expression of IL-6, while lower molecular weights are related to the secretion of IL-4 and chemotactic cytokines (IL-8 and MCP-1). LRAP and TRAP might be the active components of lower molecular weight fractions as shown in the mass spectrometric analysis.
Footnotes
Acknowledgements
We are thankful to Aina Maria Lian (Oral Research Laboratory, Institute of Clinical Dentistry, University of Oslo, Norway) for skillful technical assistance.
Supplementary material
The supplementary material for this article can be found online.
Declaration of conflicting interests
The authors declare no conflicts of interest.
Ethical approval
Not required.
Funding
This work was supported by the University of Oslo and the Norwegian Research Council.