
Abstract
Auto-immune diseases are a form of chronic disorders in which the immune system destroys the body’s cells due to a loss of tolerance to self-antigens. Systemic lupus erythematosus (SLE), identified by the production of autoantibodies in different body parts, is one of the most well-known examples of these diseases. Although the etiology of SLE is unclear, the disease’s progression may be affected by genetic and environmental factors. As studies in twins provide adequate evidence for genetic involvement in the SLE, other phenomena such as metallization, histone modifications, and alterations in the expression of noncoding RNAs (ncRNAs) also indicate the involvement of epigenetic factors in this disease. Among all the epigenetic alterations, ncRNAs appear to have the most crucial contribution to the pathogenesis of SLE. The ncRNAs’ length and size are divided into three main classes: micro RNAs, long noncoding RNAs (LncRNA), and circular RNAs (circRNAs). Accumulating evidence suggests that dysregulations in these ncRNAs contributed to the pathogenesis of SLE. Hence, clarifying the function of these groups of ncRNAs in the pathophysiology of SLE provides a deeper understanding of the disease. It also opens up new opportunities to develop targeted therapies for this disease.
Keywords
Introduction
Auto-immune diseases are a group of chronic disorders in which the immune system attacks the body’s own cells. Loss of immune tolerance to self-antigens causes this phenomenon. Although the cause of these diseases is not well known, they affect around 3% of the European and American populations. 1 Environmental and genetic variables, hormones, and infectious diseases have all been identified as major contributors to the development of auto-immune disorders in studies. 2 Some of the well-recognized auto-immune disorders include Addison’s disease, Hashimoto’s thyroiditis, multiple sclerosis (MS), Sjögren’s syndrome, systemic lupus erythematosus (SLE), and type I diabetes (T1D). 3 Although the symptoms of auto-immune diseases vary and depend on the type and location of the disease, the general indications of these disorders include fatigue, fever, lethargy, joint pain, and rash. Despite advances in studies related to auto-immune diseases, there is still no definitive treatment for this disorder. However, antimalarial drugs, steroidal and nonsteroidal anti-inflammatory drugs, immunosuppressive agents, and target therapies such as cytokine inhibitors [e.g. tumor necrosis factor-alpha (TNF-α) blockers] can all prevent the progression of auto-immunity.4,5 SLE is a histologically heterogeneous auto-immune disease defined by the development of autoantibodies directed against nuclear antigens. 5 This disease, which most frequently affects young to middle-aged women, has an incidence of 1 to 10 per 100,000 people annually. 5 Due to the uncertainty of SLE’s cause, several factors like genetic and environmental variables (epigenetic) have been studied in the incidence of SLE. 6 On the other hand, epigenetic factors, including methylation, histone modification, and especially ncRNAs, may be involved in the pathogenesis of SLE. ncRNAs are synthesized from a bigger portion of the genome that does not encode proteins and are implicated in gene expression and protein activity control. 7 MicroRNAs, long noncoding RNAs (LncRNA), and circular RNAs (circRNAs) are three of the most significant types of ncRNAs. MicroRNAs are small, endogenous, 19- to 25-nucleotide (nt) RNAs that inhibit the translation of specific mRNAs.8–11 Another type of ncRNA is LncRNAs which have more than 200 nt and do not convert to functional proteins. 12 CircRNAs are endogenous RNA molecules that can range in size from remarkably short (100 nt) to more than 4 kb and include exonic or intronic sequences of their parental genes. 13 According to studies, ncRNAs have a key role in B-cell activation and the development of SLE by the B-cell activation and the nuclear factor kappa light chain enhancer of activated B cells (NF-κB), type I interferons (IFN-I), and tumor growth factor (TGF) signaling pathways. 14 In this review, we will discuss the factors involved in the pathogenesis of SLE, especially microRNAs and LncRNAs, and their possible therapeutic use in the course of the disease.
Auto-immune disease
Auto-immune disorders are a group of long-lasting diseases that begin with the body’s inability to distinguish its cells from foreign cells, and the result is an attack and the development of an immune response against self-antigens. 1 Contrary to the low prevalence of auto-immune diseases (about 3%), their substantial influence on mortality cannot be ignored. 15 Normal immune responses often result in the removal of foreign antigens, e.g. pathogen-associated molecular patterns (PAMPs) and cancerous antigens from the body. Contrary to the physiological state, in auto-immune diseases, the immune system is unable to distinguish between self-antigens and foreign antigens. The result is the generation of excessive immune response to self-antigens and subsequent organ damage.3,16 Although a variety of factors may contribute to the generation of auto-immunity, immune tolerance breach is a crucial mechanism in the pathogenesis of this disease. 17 Central tolerance in the thymus and bone marrow is critical for immune system homeostasis. Before maturing and entering the bloodstream, developing lymphocytes in the thymus undergo positive selection in the cerebral cortex. It is noteworthy that in a healthy host, the thymic medulla negatively selects and eliminates lymphocytes with potential self-reactivity. 18 Peripheral tolerance or secondary selection of mature T cells occurs after exiting the thymus. In this phenomenon, most of the self-reactive T cells are eliminated or become anergic. 18 In addition, immature B cells are eliminated through a process named clonal deletion or clonal anergy if they express surface IgM that identifies common self-cell-surface antigens. 18 The ‘receptor editing’ approach permits autoreactive B cells to evade elimination. Peripheral tolerance can affect mature B cells as well. 18 The breakdown of these immune tolerance mechanisms and the presence of these potentially self-reactive T and B lymphocytes or their capacity to generate autoantibodies cause the onset of classical or pathological auto-immunity.2,19 However, even in healthy individuals, although peripheral and central tolerance is carefully maintained, some potentially autoreactive lymphocytes are present in the periphery, causing no symptoms of auto-immune disease.2,19 This state is named physiological auto-immunity, which is without evidence of clinical disease. The body’s natural antibodies, which aid in the removal of degraded self and foreign antigens to preserve homeostasis, are evidence of this condition. 20 Furthermore, despite the unknown factors influencing the immune response to this disease, the impact of environmental and genetic factors has been discussed in several studies.2,21,22 Numerous genetic polymorphisms, the majority of which are found in the regulatory regions of genes, have been implicated in the development of a variety of auto-immune diseases.23,24 For instance, among all the genes linked to auto-immune disorders, those with specific human leukocyte antigen (HLA) alleles had the highest and longest-standing correlations. 24 For instance, T1D (HLA-II: DQ2 and DQ8; HLA-I: HLA-A and DQB1*0602), rheumatoid arthritis (RA) (HLAII: DR4; HLA-III: TNF), auto-immune thyroid disease (HLA-II: DR3 and DR4), and psoriasis (HLA-I:Cw*0602, Cw1203, and HCP5) are highly correlated with certain HLA alleles. 25 Moreover, several auto-immune illnesses have been related to genetic variations in cytokines and cytokine receptors. Ankylosing spondylitis, Behcet’s disease, Crohn’s disease, psoriasis, and ulcerative colitis are all disorders where the IL23R gene has been found to have genetic variations. 26 In addition, rare cases of fulminant auto-immunity caused by genetic changes in a single gene have been reported. In light of this, auto-immune polyendocrine syndrome (APS) and immune dysregulation polyendocrinopathy enteropathy X-linked (IPEX) are instances of monogenetic auto-immune disorders. Auto-immune regulator (AIRE) and forkhead box P3 (FOXP3) mutations, respectively, cause these disorders directly.27,28 Growing evidence indicates that the development of auto-immune disease depends on environmental variables in addition to genetic predisposition. Nutrition, the microbiota, infectious processes, and xenobiotics such as cigarette smoke, pharmaceuticals, hormones, ultraviolet (UV) radiation, silica solvents, heavy metals, vaccinations, and collagen/silicone implants are among these environmental influences.29–31 It has long been believed that infections can cause auto-immune disorders.32,33 For instance, MS has been linked to postmortem brain tissue showing indications of Epstein–Barr virus (EBV) infection, but not other inflammatory diseases. 34 Furthermore, it was demonstrated that patients with relapsing–remitting MS have been known to experience relapses due to systemic infections that increase myelin-specific T-cell responses. 32 RA and periodontal infections provide another evidence of the correlation between infections and auto-immune. 35 Moreover, the potential effect of the microbiome on local and systemic immune responses has also been reported. Immune responses to intestinal commensal microorganisms that are dysregulated and too aggressive lead to inflammatory bowel disease (IBD). 36 In addition, commensal microorganisms have been linked to auto-immune diseases including T1D. 36 UV radiation is a nonmicrobial environmental factor that has been linked to auto-immune disorders like cutaneous lupus. This association may be addressed by the concept that UV radiation causes the apoptotic death of many different cell types and enhances the burden of nuclear antigens, particularly if the dead cells cannot be effectively removed. 37 It has been proposed that groups of tolerance-promoting dendritic cells (DCs) preserve peripheral tolerance to tissue antigens via low-level spontaneous cell death in tissues. This system may become easily overwhelmed in lupus patients because of a genetic predisposition that renders them unable to maintain tolerance in the presence of constant UV exposure. 38 Several causes can contribute to the development of auto-immune diseases. For instance, it was noted that there is presumably an imbalance between effector T cells and functional regulatory T cells (Tregs) in T-cell-dependent inflammatory auto-immune disorders. 2 This pathogenic imbalance was supported by mouse models of auto-immunity. 39 Moreover, human auto-immune diseases may also be initiated by decreased numbers of functional Tregs, or by the resistance of effector T cells to regulation. 2 Antigen-presenting cells (APCs) can present denatured or extracellular peptides atypically, resulting in peptide/major histocompatibility complexes (MHCs), that are distinct from those typically generated inside APCs, and are therefore capable of activating T cells that may be potentially pathogenic. 40 The identification of conformational isomers of the peptide/MHC or a difference in the binding register of a peptide within the groove of the MHC molecule may be the cause of this unusual activation of potentially self-reactive T cells.41,42 Besides altered peptide/MHC recognition, auto-immunity can be triggered by an early innate immune response. 43 In light of this, the lack of ubiquitin-modifying enzyme A20 leads to lethal auto-immunity resulting from unregulated Toll-like receptor (TLR) signals mediated by MyD88. 44 Also, due to enhanced B-cell identification of nucleolar antigens in Yaa mice with a genetic duplication of TLRs-7, these mice exhibit a spontaneous SLE-like syndrome. 45 SLE, one of the most prevalent auto-immune diseases, is a multisystem auto-immune disorder that targets the body’s tissues and results in significant organ damage and inflammation. Despite much investigation, the etiology of SLE is still unclear. Both genetic and environmental variables have been related to the pathogenesis of SLE. In the following sections, we will discuss this disease in detail.
SLE
SLE is one of the most prevalent auto-immune diseases that can affect any organ and part of the body and cause irreversible complications. The literal meaning of lupus in ancient Latin goes back to the wolf, and the name of this disease is because in the 13th century, for the first time, the physician Rogerius likened the facial lesions of a person with this disease to imitating the bite of a wolf. 46 Since then, many researchers have worked on it and have used many descriptions of its skin symptoms, including erythema centrifugum or lupus discoid and disease with butterfly lesions on the face.46–48 Other pathological symptoms and features of this chronic disease were later identified, including central nervous system (CNS) involvement, inflammation, and vascular abnormalities such as vasculitis and immune complex deposition.49,50 Essentially, the clinical manifestations of lupus imitate a viral syndrome with signs of weight loss, fatigue, and low-grade fever with other skin, kidney, respiratory, cardiovascular, CNS, and gastrointestinal indications. 50 Epidemiologically, the disease is more common in adult women during the reproductive period, about 10:1 compared with men. Despite the lack of a comprehensive global method for identifying the true prevalence and incidence of the disease, the yearly incidence of SLE is estimated at 1–10 per 100,000, and the prevalence is 20–70 per 100,000 per year.51–54
SLE has a complex etiology that involves environmental factors and genetic predisposition (Figure 1). As with other auto-immune diseases, loss of immune tolerance to self-antigens along with sustained autoantibody generation are basic pathologic processes in the development of SLE. 50 The tolerance mechanisms in SLE patients have been found to exhibit several obstacles, including defects in the regulation of mature B cells in peripheral tissues, receptor editing processes, and deletion of immature B cells in the bone marrow. 55 Also, a failure in the early B-cell tolerance checkpoints may be a factor in the establishment of SLE since mature naïve B cells in patients with this condition can generate autoantibodies even before the encounters with antigens. 55 Besides, another fundamental concept in SLE pathogenesis is an imbalance between apoptotic cell generation and apoptotic material removal. During apoptosis, the cell membrane creates blebs that pinch off from the cell and include fragmented cellular material, particularly nuclear antigens, which are normally inaccessible to the immune system. 56 Daily, neutrophils in humans undergo apoptosis; exposure to UV radiation, infections, and toxins can increase the apoptosis of these cells, which are all established to be linked to SLE. 57 Furthermore, studies of peripheral blood samples from lupus patients reveal pyroptosis, a form of cell death, as a new pathway for the disease. Accordingly, dying and dead cells can produce a large amount of immune complex components in the blood. Consequently, pyroptosis can significantly affect the auto-immune response in these diseases. Gasdermin D (GSDMD) has recently been identified as the ultimate pyroptosis tormentor in the activation of inflammatory diseases and hence can play an important role in the pathogenesis of SLE (NCT03984227). All of these apoptotic fragments are often quickly removed and are inaccessible to the immune system. However, abnormalities in the removal of apoptotic cells can lead to the development of SLE by immune dysregulation. In this regard, these auto-antigens are recognized as nonself by the innate immune system cells in SLE due to increased cell death and reduced removal of cellular debris. 58 For instance, nucleic acid recognition receptors, like TLRs, can be triggered by persistent apoptotic debris containing nucleic acids in order to initiate an inflammatory response. 59 In addition to the identification of viral infections and protection against intracellular bacteria, nucleic acid recognition receptors are linked to the generation of IFN-I (IFNα/β). 57 The pathogenesis of SLE is currently thought to be highly influenced by abnormalities in these pathways, which both increase disease susceptibility and directly cause monogenic types of SLE. 57 The differentiation of B cells and loss of tolerance are stimulated by IFN-I. 57 Nucleic acid recognition receptor–ligand interactions not only cause the production of IFN-I but also promote the generation of additional innate immune mediators like IL-1, IL-6, and TNF-α. These mediators stimulate APCs, resulting in enhanced antigen presentation to T cells. 60 Both surface IgM receptors for proteins complexed with nucleic acids and direct antigen detection by B lymphocytes allow them to respond to nucleic acids. 57 Also, in SLE patients, due to the defects in the removal of auto-antigens, nuclear material, and modified auto-antigens accumulate on germinal center follicular dendritic cells (FDCs), and autoreactive B cells are exposed to these modified auto-antigens by these FDCs. 61 After being triggered, these B cells mature, proliferate, and start secreting more antibodies, which strengthens the immune system’s adaptive response. 57 Besides, TNF-α, IL-4, IL-6, and IL-10 promote auto-antibody secretion and inflammation. 57 As most of the auto-antibodies found in SLE are high-affinity, somatically mutated IgG, this implies that the germinal centers, where T cells aid in class switching, are the likely site of their origination.49,62 T cells are therefore crucial to the development of the pathogenesis of SLE. In this regard, antigen uptake by APCs is degraded and presented to T cells as peptides–MHC complexes to initiate their responses. The stimulation and differentiation of T lymphocytes are also influenced by the presence of soluble mediators. 58 Pathogenesis of SLE has been linked to changes in the number of cytokines secreted by T cells, including elevated cytokine secretion of type 1 T helper (Th1) (IFN-γ, IL-2), type 2 T helper (Th2) (IL-5, IL-13), and type 17 T helper (Th17) (IL-6, IL-17A, IL-21) and decreased Treg responses, including diminished TGF-β and IL-10 production.63–65 Increased T-follicular helper (Tfh) cell responses promote antibody-producing B cells with a high affinity for clonal expansion in germinal centers. 66 For example, in SLE patients, the production of pathogenic IgG antibodies with a high affinity for double-stranded DNA (anti-dsDNA) results from the interaction of B and T lymphocytes.49,62 Taken together, lymphocyte signaling, IFNα/β generation pathways, nucleic acid sensing, inefficient removal of biological debris like apoptotic cells, and neutrophil extracellular traps are all key contributors to loss of self-tolerance and tissue damage in SLE patients. Genome-wide association studies (GWAS) also support the involvement of this pathway in the pathogenesis of SLE.67,68 As stated, environmental factors affect the pathogenesis of SLE. In this regard, SLE has long been acknowledged to be influenced by hormones and UV radiation.69,70 It has been estimated that 90% of SLE cohorts consist of women, and estrogen and prolactin are known to enhance immune responses through a variety of mechanisms.71,72 It is believed that UV light stimulates the immune system by promoting apoptosis. 73 The role of EBV and cytomegalovirus infections has also been shown in the pathogenesis of SLE. 74 Moreover, there are many investigations on the correlation between SLE and genetics, and the higher incidence in monozygotic twins (24–58%) than dizygotic twins (2–5%) is proof of the accuracy of this hypothesis. 6 Also, SLE is highly correlated with certain HLA alleles (HLA-II: DR3, DR2, and DR8; HLA-III: SCIVaL, CFB, RDBP, DOM3Z, STK19C4A, and C4B). 25 In addition, this disease is influenced by epigenetic mechanisms such as noncoding RNAs (ncRNAs) and DNA methylation (the following sections will provide more details). 62 Due to the complexity of SLE and its treatment methods, these patients must have a comprehensive care plan. These treatments vary depending on the clinical manifestations and may include antimalarial drugs, glucocorticoids, immunosuppressants, and biological drugs. Also, nonsteroidal anti-inflammatory drugs (NSAIDs) can be used to limit inflammation or ache. 75

Factors that contribute to SLE pathogenesis. SLE pathogenesis is influenced by a variety of variables, including genetics, environment, hormones, and epigenetics. Each of these factors, in turn, triggers its own antibodies against the immune system. By presenting these antibodies on lymphocytes and producing various immunoglobulins, inflammatory pathways are activated and destroy various tissues, including the kidneys, skin, lungs, brain, and heart.
Different mechanisms involved in the development of SLE
Lupus is an auto-immune and heterogeneous disease that has been labeled a mysterious and cruel disease in the United States due to its complexity and ambiguity. 76 Although the definite cause of this complex multifactorial disease has not yet been identified, several variables such as genetic, epigenetic, ethnic, immune, hormonal, and environmental factors are involved in its progression. As a result, each of these factors causes abnormal activation of T lymphocytes and other immune cells, and eventually, the accumulation of these cells causes damage to the target organs.77,78 The higher incidence of SLE in first-degree relatives (5.87%) and high heritability (43.9%) is also a reason for the genetic influence on this disease. According to the available evidence and research, a defect in one gene alone cannot cause disease, and the cooperation of several genes and alleles is required for its occurrence. However, it has been shown that defects in complement component genes such as 1q (C1q), subcomponent A (C1QA), C1QB, C1QC, and DNA repair genes like exonuclease 1 (TREX1), or deoxyribonuclease 1-like 3 (DNASE1 L3) can also trigger the monogenic forms of the lupus-like disease.78,79 According to relevant studies in GWAS, HLA genes are found on chromosome 6 and in MHC regions, which are high polymorphic areas. Because of the diversity in its genes, especially MHC II, they can have a greater odds ratio (OR) of SLE involvement. The presentation of specific peptides to CD4+ T cells to remove self-reactive lymphocytes (negative selection) and stimulate the production of T regs by MHC II occurs during the maturation of T cells in the thymus. As a result, if mutations occur in these areas, this process will be disturbed.80–84 Considering the effect of gender on the SLE, it can be noted that because of the effect of estrogen on humoral responses and maintaining the self-response of B cells, the ratio of women to men is 10:1. Compared with estrogen, testosterone appears to suppress anti-DNA antibodies and somehow prevent the development of SLE.85–87 Furthermore, environmental factors encompass a wide range of elements that influence the progression of SLE. Epstein–Barr virus, light exposure, and smoking are examples of biological, physical, and chemical stimuli that increase the development of anti-dsDNA antibodies and cell necrosis, respectively. In addition, certain medications can also cause iatrogenic lupus. 76 Although these factors are significant in the development of SLE, hereditary background and regulation of susceptibility genes are also required. Environmental factors affect the growth of self-reactive lymphocytes and interfere with the processing and presentation of antigens. They also expose B and T lymphocytes to microbial agents and trigger the polyclonal activation of these cells. With the cooperation of these factors, the incidence of SLE and its consequent damages will increase. 87 Epigenetic changes also have a significant influence on genetic diversity in many diseases, including SLE. Alterations such as DNA methylation, histone modifications, and ncRNAs are all included and determine how regulatory disturbance happens in SLE. 77 Global DNA hypomethylation is present in human T lymphocytes of active SLE patients, particularly in those with lupus nephritis.88,89 Since DNA methylation is normally restrictive, the result of this hypomethylation is frequently the upregulation of genes. For example, promoter hypomethylation in this disorder causes the overexpression of genes like CD11a and CD70 in T cells. 88 Furthermore, IFN-stimulated genes (ISGs) were notably hypomethylated when evaluated on a genome-wide basis in individuals with SLE. 89 The extracellular signal–regulated kinase (ERK)1/2 is essential for the induction of T-cell responses. Both independent and dependent responses to the T-cell receptor (TCR) trigger the ERK pathway in T cells. 90 ERK signaling pathway also regulates the expression of T-cell DNA methyltransferase (Dnmt). 90 As the ERK signaling pathway regulates the expression of T-cell Dnmt, 90 defects in this signaling pathway can lead to epigenetic abnormalities in T cells, such as a decrease in DNA methylation at the genome level. 90 In line with this, it has been shown that defective T-cell ERK signaling is associated with the pathogenesis of SLE in the mouse model of the disease. 88 The generation of anti-dsDNA antibodies is induced by defective ERK signaling in T cells. 88 Similar to T cells in human lupus, reduced ERK pathway activation in T cells causes upregulation of the methylation-sensitive genes CD11a and CD70 and reduced expression of Dnmt-1. 88 The degree of T-cell ERK signaling impairment as well as the anti-dsDNA antibody titer is positively correlated with disease severity in lupus patients.91,92 Furthermore, autoreactivity and altered cytokine release may both be caused, directly or indirectly, by aberrant lupus T-cell DNA methylation.92,93 Another crucial signaling pathway in the immunopathogenesis of lupus is phosphoinositide 3-kinase (PI3 K)/Akt (Ak strain transforming)/mammalian target of rapamycin (mTOR) pathway. Cellular proliferation and expression of inflammatory cytokines are greatly influenced by this signaling pathway. 94 SLE patients’ CD4+ T cells had higher Akt expression and phosphorylation levels than healthy donors’ CD4+ T cells, which suggests that PI3 K and mTOR activity have been elevated. 95 There has also been evidence of increased p70S6k (one of the mTOR substrates) activity in SLE CD4+ T cells. 95 Also, when compared with healthy donors’ peripheral blood mononuclear cells (PBMCs), SLE patients’ PBMCs had significantly greater levels of Akt phosphorylation and lower levels of p27kip1 expression. 96 Increased Akt activity and decreased p27kip1 expression appear to be factored in the passage of SLE lymphocytes through the G1/S cell cycle checkpoint. As a result, SLE lymphocytes gather in the G2/M and S-cell-cycle stages as they prepare to undergo proliferation. 96 The association between SLE pathogenesis and ncRNAs is now well established. Contrary to previous notions of these RNAs as evolutionarily junk, they have a substantial role in the molecular processes and pathogenesis of several disorders by forming a large part of the whole transcriptome.33,34 One of the most important ncRNAs associated with the occurrence of SLE and its pathogenesis is microRNAs. These small 22-nt ncRNAs are responsible for the negative regulation of gene expression that binds to target mRNA. In other words, it degrades or inhibits RNA translation and silences it after attaching to the target.76,97 So, ncRNAs affect pivotal signaling pathways involved in SLE pathogenesis, such as NF-κB, IFN-І, and TGF pathways. 14 In addition, the different roles of microRNAs in signal transducer and activator of transcription (STAT) pathway regulation are central. Due to the incorrect regulation of this pathway in SLE patients and its importance in Th17 differentiation, an increase in IL-17 and Th17 rates is observed. 98 Furthermore, ncRNA influenced the activation and differentiation of CD4+ T cells. Thus, dysregulation of ncRNAs such as miR-21, miR-155, miR-31, miR-29a, miR-126, miR-142-3p/5p, and miR-183 C, as well as LncRNAs in CD4+ T cells or PBMCs, has been observed to increase the incidence of SLE through direct and indirect regulation of CD4+ T cells. This is followed by a high proliferation of Th1/Th17/ Tfh cells, which reduces the number of Treg cells and, in this way, contributes to the development of SLE. 99 Also, in a clinical study of pregnant women with SLE, it was found that there was a 40% increased risk of flare-up symptoms with the onset of pregnancy, even in stable and asymptomatic patients. Epigenetic modification and changes in the expression of microRNAs during pregnancy are supposed to be the cause of increased signs and symptoms as well as the risk of flares in SLE (NCT02350491). Besides, recent studies have attempted to elucidate the involvement of circRNAs in SLE via microarrays and high-throughput sequencing. Their results have revealed the involvement of these molecules in the pathogenesis of SLE.100,101 The ncRNAs help identify and differentiate SLE patients besides performing well in diagnosing disease complications. Despite a thorough investigation of coding regions and their association with single-nucleotide polymorphism (SNP) in the pathogenesis of SLE, studies related to noncoding regions as substantial risk factors for SLE need further investigations to be more clarified.14,102 Figure 1 illustrates the pathogenesis-related factors for SLE. In the following sections, we will quite describe ncRNAs. However, there is a need for more studies to clearly understand the importance of these RNAs in SLE’s pathogenesis.
The role of microRNAs in the development of SLE
MicroRNAs are short, endogenous, and ncRNAs 19- to 25-nt long, which perform various functions including cell cycle regulation, differentiation, proliferation, apoptosis, stress tolerance, energy metabolism, and immune response.8–11 Targeting specific mRNAs and destroying or suppressing their translation is another regulatory role of these ncRNAs.9,11 The first short RNA, known as lin-4, was identified in 1993 while screening nematodes. Coincidentally, in the same year, the regulation of lin-14 was also determined by lin-4 and showed the regulatory function of these RNAs for the first time.103,104 To this point, 1917 pre-microRNAs and 2654 mature microRNAs have been identified in Homo sapiens, whose target sites have been predicted by 60% of human coding genes. Mature microRNA is transcribed from the primary microRNA (pri-microRNA) during several stages of its biogenesis. They often originate from introns or LncRNAs and are copied by RNA polymerase II. 105 Numerous studies have shown an association between microRNA expression in peripheral blood cells, body fluids, and damaged tissues of SLE patients with severe damage to various organs in these individuals. For instance, in a study on 100 patients with SLE aged 18 and above who were cured with cyclophosphamide, hydroxychloroquine, and hydrocortisone, the expression levels of microRNAs, e.g. miR181a, miR196a, and miR21, were evaluated for 3 months to record changes in their expression (NCT02756546). According to these findings, microRNAs are significant biomarkers in diagnosing auto-immune diseases like SLE. Improper regulation of microRNAs in T cells can cause immunodeficiency and ultimately lead to auto-immune disease. 106 As a result, investigating the association between microRNAs and auto-immune diseases has a lot of importance nowadays. Table 1 provides a list of microRNAs related to SLE disease. Besides, the role of microRNAs in the pathogenesis of SLE is depicted in Figure 2.
MicroRNAs related to SLE disease.

ANA, anti-nuclear antibodies; anti-dsDNA, anti-double-stranded DNA; BALB/C, Bagg Albino; C, complement; C-C motif, the chemokine; CCL2, ligand 2; circRNAs, circular RNAs; CRKL, Crk-like protein; CRP, C-reactive protein; CXCL10, C-X-C motif chemokine ligand 10; DMEM, Dulbecco’s Modified Eagle Medium; ESR, erythrocyte sedimentation rate; FBS, fetal bovine serum; HIF-1, hypoxia-inducible factor; IFNA1, interferon-alpha receptor; IL, interleukins; LC3, autophagy-related 7, microtubule-associated protein 1A/1B-light chain 3; MRL/MPJ-Fas lpr/J, Murphy Roths large/homozygous for the lymphoproliferation spontaneous mutation; MSCs, mesenchymal stem cells; MTMRT3, myotubularin-related protein 3; NPA, nonbilayer phospholipid arrangements; PBMC, peripheral blood mononuclear cell; SLE, systemic lupus erythematosus; SLEDAI, Systemic Lupus Erythematosus Disease Activity Index; TLRs, Toll-like receptors; TNF-α, tumor necrosis factor-alpha; UC-MSC, umbilical cord mesenchymal stem cells.

Role of microRNAs in the SLE. Many microRNAs, including miR-181, miR-146, miR-155, etc., are involved in the pathogenesis of SLE. High or low expression of any of these ncRNAs regulates multiple signaling pathways and is somehow involved in the pathogenesis of this auto-immune disorder.
MiR-181
MiR-181 is a protected family consisting of miR-181a, miR-181b, miR-181c, and miR-181d, in which expression of miR-181a and miR-181b is generally observed in the brain, bone marrow, spleen, and thymus. These microRNAs are located on chromosomes 1, 9, and 19 in clusters. They have a main part in the regulation of B and T cells in auto-immune diseases. 133 MiR-181a, being expressed in several tissues, inhibits the proliferation of macrophages and induces apoptosis in them. Besides, by inhibiting the differentiation of Th1 cells, miR-181a and miR-181b differentiate regulatory T lymphocytes, which are responsible for hindering inflammation. 108 As miR-181a has a central function in T lymphocyte maturation and development, its aberrant expression may have an association with auto-immune disorders such as SLE. 108 A study was conducted by Lee et al. to evaluate miR-181a levels in active SLE, inactive SLE, and healthy groups. They discovered that the SLE group had greater serum levels of miR-181a than the normal group. Also, the levels of miR-181a in inactive SLE patients were upper than in both other groups. 108 On the other hand, overexpression of miR-181a in the peripheral blood of SLE patients has also been observed. 134 Yong-Ling and colleagues showed that miR-181b facilitates SLE development by directly targeting molecular IFN alpha 1 (IFNA1). For this purpose, they also investigated two SNPs (rs1332190 and rs10811543) located in the IFNA1 promoter region to analyze gene interactions. Interestingly, they found an association between miR-181b rs322931 C and SLE and its uptrend. Although IFNA1 rs1332190 and rs10811543 alone were not significantly related to SLE, a combined analysis of these three factors simultaneously augmented the risk of disease progression. Therefore, He et al., 109 finding a correlation between miR-181b expression and rs322931 and mentioning a downregulation in miR-181b levels in SLE patients due to rs322931 CT /TT genotype compared with normal individuals once again noted the importance of ncRNAs in SLE. Due to the effects of SNPs and their correlation with microRNAs, the association of autophagy-related gene, MTMR3 SNP rs12537, which is the main target gene of miR-181-a, has been investigated with SLE progression. Based on the results reported by Senousy et al., MTMR3 rs12537 TT is considered a risk factor in SLE with a higher risk of autophagy and has an unfavorable prognosis with it. Notably, miR-181a plays a crucial role in the development of SLE. 110 Remarkably, a case–control study also investigated three miR-181 cluster polymorphisms and their association with SLE in the Chinese population. According to this study, Wang et al. reported a higher association of the rs8108402C>T polymorphism on the miR-181c/d gene promoter compared with other genotypes in SLE. 135 Contrary to these findings, in another study, it was found that miR-181a downregulation occurs in T lymphocytes of SLE patients and indicated that using umbilical cord mesenchymal stem cells (UC-MSCs) can upregulate miR-181a, which may help improve the condition of SLE patients. 107
MiR-98
MiR-98, with a size of 119 bases, originates from the let-7 family and is involved in Niemann–Pick disease, type C1, and head and neck squamous cell carcinoma. Moreover, the role of let7/miR-98 in cell proliferation, differentiation, apoptosis, and angiogenesis has been discussed in several studies.136–141 We can highlight the influence of mir-98 in SLE because of the increased apoptosis of T cells in SLE and the association of mir-98 with this phenomenon. It is noteworthy that the amount of mir-98 in SLE patients’ CD4+ T lymphocytes was lower than in normal people. 115 Therefore, this deficiency can be attributed to a defect in the regulation of the apoptosis pathway in SLE patients. As a result, mir-98 can be utilized to improve the condition of these patients by modulating apoptotic pathways while preserving lymphocyte homeostasis. 115 Several studies found a negative correlation between mir-98 and IL-6.142,143 By binding to 3′UTR of IL-6 mRNA, mir-98 negatively regulates this cytokine. The result is increased PBMC proliferation and suppression of the inflammatory factors production such as TNF-α, IL-8, IL-1β, and IL-10 in PBMC. Also, due to the vital role of STAT3 in hindering TNF-α, the ability of STAT inhibitor, S31-201, to revoke the function of both mir-98 inhibitors and high IL-6 expression was realized. According to these findings, mir-98 could be a key target in the treatment of SLE and other IL-6-related diseases. 6
MiR-155
MiR-155, encoded by MIRHG155, or originally the B-cell integration cluster (BIC) gene, matures in the third exon of this cluster, located in the nonencoding part of chromosome 21.144,145 Due to the pivotal role of this ncRNA in regulating crucial signaling pathways, its inappropriate expression can lead to a variety of cancers and auto-immune disorders such as RA, MS, and SLE. 146 The serum cell–free miR-155 expression profile was considered in another study in mixed connective tissue disease (MCTD), SLE, scleroderma and systemic sclerosis (SSc), and RA disease. Its significant expression in SLE in comparison with other disorders can indicate the substantial role of this miR in SLE. 121 MiR-155 has also been documented to have a higher expression in the peripheral blood of SLE patients than normal individuals, which may show renal injuries in these patients. 119 Another study by Shumnalieva et al. investigated the correlation between miR-155 and its changes in whole blood. According to this study, the expression of miR-155 in SLE patients compared with the control group was reported to be 50%, indicating that more studies are needed in this regard. 117 MiR-155 deficiency has also been linked to lesions and complications reduction in Fas LPR/LPR mice with SLE. 147 In contrast, Wang et al. 120 reported a decrease in the expression of miR-155 in serum and urine samples of patients with SLE compared with controls, which was consistent with the level of kidney function of these people. Due to the positive correlation between serum miR-155 levels and lymphocyte count, they have proposed that lymphocytes are the major source of serum miR-155 in SLE patients. Hence, miR-155 expression was likely reduced as a result of the decrease in the serum’s lymphocyte count. In addition, a nonsignificant decrease in the expression of miR-155 in SLE patients compared with the control group was also reported in the study of Latini et al. 148 They have suggested that this may be associated with miR-155’s regulatory roles in immune cells such as T and B cells.
MiR-146
MiR-146a is another type of ncRNAs located on human chromosome 5 in the second exon of the LOC285628 gene. The association of miR-146a with the pathogenesis of several diseases, including auto-immune disorders such as SLE, RA, and Sjögren’s syndrome, has been reported in various studies. 149 In this line, a research was conducted on 42 female SLE patients and 39 matched healthy individuals to evaluate the association between miR-146 and SLE’s ocular symptoms. Based on their results, patients with symptoms including dry eye, cataract, keratitis, and drusen showed less expression of miR-146 than asymptomatic patients. However, high expression of miR-146 was also shown in people with retinopathy symptoms. Consequently, a remarkable association was found between miR-146 and light sensitivity in SLE disease. 126 Of note, an association with secondary antiphospholipid syndrome (APS), which is positive in 30–40% of SLE patients, has been linked to miR-146. Even though there was no significant difference in miR-146 expression between patients with and without secondary APS, SLE patients had decreased expression of this miR-146 when compared with the control group. 123 Stimulation of SLE-associated renal injury in medical research laboratory lymphoproliferation (MRL/lpr) mice can be alleviated by miR-146a. 150 It was shown that miR-146a inhibited classical and nonclassical NF-κB pathways and moderated kidney damage by reducing auto-antibody production through B cells such as anti-dsDNA, anti-single-stranded DNA (anti-ssDNA), and anti-nuclear antibody (ANA). Furthermore, miR-146a can regulate the amount of protein and blood urea nitrogen (BUN) in the urine. Moreover, it can diminish inflammatory factors in the kidney tissues of MRL/lpr mice and moderate their effects on this organ. 150 Considering the obvious and non-negligible role of senescence of MSCs in SLE progression and decline of miR-146a expression in these patients, Dong and colleagues investigated the function of miR-146a in the control of MSC aging using the SA-gal assay. By imitating the functions of miR-146, they attempted to reverse the aging process in these patients, which shows the regulatory role of exosomal miR-146a in this pathway. 125 Also, according to Dominguez-Gutierrez et al., 124 the loss of the ability of miR-146 in SLE patients may reduce its efficiency in regulating the STAT1a isoform.
MiR-210
MiR-210 is an ncRNA that is often induced by hypoxia and is therefore called the ‘hypoxamir master’. 151 MiR-210 has a stem-loop on the intron of chromosome 11p15.5 regulated by both the hypoxia-inducible factor (HIF)-1α and HIF2α genes and is involved in tumor progression, myocardial infarction, and ischemic skin lesions with irregular expression.152–154 According to several studies, miR-210 and HIF1α may be related to a variety of auto-immune diseases including SLE. By purifying the CD4+ T lymphocytes of lupus susceptible mice, high expression of miR-210 and HIF1α was shown on these cells. However, neither of these two factors was significantly altered in CD8+ T cells or CD19+ cells. Nevertheless, the miR-210/HIF1α pathway and its irregular expression can be a new regulatory pathway in SLE. 155 Similarly, in a similar study, miR-210 and HIF1, ERK1/2, and PI3 K/AKT were investigated, and it was indicated that the recognition of these biomolecules can play a part in the diagnosis of SLE. 113
MiR-21
MiR-21, an oncomiR, is known as a tumor suppressor due to its considerable role in apoptosis and necrosis and its importance in a wide variety of malignancies like breast, colon, and gastric cancer. 156 Remarkably, miR-21 has become significant in auto-immune diseases. Despite preserving its low expression in inactivated T lymphocytes, its expression rate is significantly increased after T-cell activation.157–159 According to studies, miR-21 causes auto-immune disorders such as T1D, psoriasis, MS, RA, and SLE via stimulating inflammatory pathways. 156 MicroRNA profile identified miR-21 as an upregulated ncRNA in SLE CD4+ T lymphocytes. Moreover, further studies demonstrate that miR-21 expression is significantly enhanced in SLE patients. In addition, this upregulation was more pronounced in people with lower C3 complement levels than in others. 114 As hypoxia is one of the causes of miR-21 induction, it can affect the pathogenesis of SLE. The study of miR-21 in the serum, urine, and T cells of SLE patients showed more production of miR-21 in these patients than in healthy people. As a result, miR-21 might be utilized as a biomarker to diagnose and treat SLE. 113
MiR29a
This 64-base microRNA is an additional ncRNA located on the 7q32.3 chromosome. 160 MiR-29a is not only involved in functions such as cell proliferation, differentiation, apoptosis, angiogenesis, tumorigenesis, and metastasis but also by binding to the genes that stimulate or inhibit the progression of cancer has carcinogenicity in several neoplasms. 161 It seems that miR-29a, like other microRNAs, can be effective in auto-immune diseases. By comparing miR-29a in the groups of SLE patients with controls, a considerable decline in the amount of this microRNA was noticed in the B lymphocytes of the first group. Downregulation of miR-29a in SLE patients has been shown to enhance Crk-like protein expression, which in turn increases IgG secretion. Therefore, it is believed that adjusting the expression of miR-29a can improve the condition of SLE patients. 130
MiR-153-3p
Other noncoding microRNAs include miR-153-3p identified in human laryngeal squamous cell carcinoma and breast cancer. 162 Similarly, this ncRNA appears to be implicated in auto-immune diseases such as SLE. According to an investigation by Li et al., 163 miR-153-3p was shown to be elevated in SLE patients and was closely associated with the activity of the disease. Based on their results, miR-153-3p overexpression reduced UC-MSCs’ migration and proliferation as well as their ability to reduce Tfhs and enhance Tregs by suppressing PELI1. In addition, the upregulation of miR-153-3p diminished the favorable efficacy of UC-MSCs in MRL/lpr mice in vivo. 163 Hence, miR-153-3p can be considered a therapeutic target for treating SLE.
MiR-125b
MiR-125b-1 and miR-125b-2 are components of miR-125b involved in controlling various signaling pathways such as NF-κB, p53, PI3 K/Akt/mTOR, ErbB2 (erythroblastic oncogene B), and Wnt (Wingless-related integration site). In addition, they may have an essential role in cell proliferation, differentiation, apoptosis, drug resistance, and tumor survival. Increased expression of miR-125b has been revealed in auto-immune diseases such as psoriasis. 164 Given this issue, it is thought that alterations in its expression may also play a role in the pathogenesis of SLE. By studying the effect of UVB on the expression of miR-125-b, reduced values of miR-125-b were observed in both control and patient groups. However, its expression was significantly lower in SLE patients exposed to UVB than in normal individuals. Finally, downregulation in miR-125-b increased UV radiation resistance–associated gene (UVRAG) expression, which also increased autophagy in the PBMC of these patients. 128
MiR-199-3p
One of the microRNAs that appears to be implicated in cancer is miR-199a-3p. Decreased expression of this biomarker in malignancies such as kidney and bladder cancer has shown its role as a tumor suppressor. In addition, it can act as an oncogene with upregulation in gastric and colorectal cancers. 165 MiR-199-3p may have a role in the development of auto-immune disorders like SLE. According to studies with upregulated amounts of IL-10 and decreased amounts of PARP-1 expression in SLE patients, Xiaoping et al. examined the association between miR-199a-3p and these factors. It was demonstrated that the expression of miR-199-3p was increased in SLE patients. In this regard, activation of the ERK/2 pathway inhibited PARP-1 expression and increased IL-10 production in SLE patients. Considering the alterations related to miR-199-3p expression and its positive and negative correlation with IL-10 and PARP-1, respectively, restoring physiological miR-199-3p levels may be a remarkable breakthrough in the treatment of SLE. 131
Other microRNAs
Based on previous data on enhanced differentiation of Th17 cells in SLE by myeloid-derived suppressor cell (MDSC)–arginase (Arg)-1, Pang and colleagues used RNA chips to identify the microRNA regulatory network between MDSC and Th17 cells. They have indicated that increasing miR-322-5p expression not only increases the differentiation of Th17 and Treg but also activates the TGF-β pathway. Since miR-322-5p expression decreased with the Arg-1 inhibitor, the Arg-1/miR-322-5p axis can be a potential therapeutic target for SLE. 166 Furthermore, since Resolvin D1 (RvD1) is a suppressor of inflammatory responses, it has been linked to a Treg/Th17 imbalance in the pathogenesis of SLE. miR-30e-5p is involved in the Treg/Th17 differentiation as an RvD1 downstream microRNA. Thus, RvD1 may enhance SLE progression by upregulating Treg and reducing Th17 cell expression via miR-30e-5p. 167 Given the importance of IL-2 in immune tolerance, its inadequate production by epigenetic modifications such as miR-200a-3p could have an impact on SLE pathogenesis. Katsuyama et al. 127 have reported that decreased expression of miR-200a-3p subsequently reduced IL-2 levels by zinc finger E-box binding homeobox (ZEB)1 and C-terminal binding protein 2 (CtBP2), which highlights the crucial role of microRNA in this disease. As previously stated, pyroptosis and GSDMD are crucial in the pathophysiology of SLE. GSDMD can act as the ultimate pyroptosis tormentor in the activation of inflammatory responses and it may be a promising therapeutic target for a variety of disorders (NCT03984227). MiR-379-5p is located in the DLK1-DIO3 region, associated with organ growth and various diseases such as cancer. While GSDMD is the primary target of miR-379-5p, high levels of miR-379-5p expression have been shown to inhibit the increased effect of arsenite-induced GSDMD levels (NCT03984227).
The role of LncRNAs in the progression of SLE
LncRNAs are RNA molecules of at least 200 nt in length that are incapable of encoding proteins. Since the early 1990s, about 172,216 copies of the human species have been registered in the NONCODE database. 168 These RNAs are transcribed mostly by Pol II/Pol I RNA polymerase (Pol) and sometimes by Pol III RNA. 169 Despite the minor part of LncRNAs in protein coding, they have imperative functions in a variety of biological and physiopathological contexts. In line with this LncRNAs have the potential to engage in cellular organization and control, including DNA replication, RNA transcription, protein translation, cell development, and cell differentiation.170,171 LncRNAs can modify chromatin function, control the assembly and function of membraneless nuclear bodies, modify the stability and translation of cytoplasmic mRNAs, and interfere with signaling pathways depending on their localization and the specific interactions they have with DNA, RNA, and proteins. In diverse biological and physiopathological contexts, these functions ultimately affect gene expression. 12 Due to their exceptional function in regulating cellular processes, they also play a role in the advancement of a variety of disorders such as cancer, inflammation, and auto-immune disease.172,173 Due to alterations in LncRNA expression and its association with SLE in blood, PBMCs, T cells, monocyte-derived dendritic cells (moDCs), and plasma of patients compared with the control group, a significant correlation of LncRNA with SLE pathogenesis was found.174–176 For instance, a clinical study evaluated the expression status of these LncRNAs in the blood samples of SLE patients and compared them with healthy individuals with real-time polymerase chain reaction (PCR) for 1 year (NCT03185767). There is evidence to suggest a link between LncRNA and how they interfere with SLE development, as outlined below. Figure 3 and Table 2 illustrate the association between LncRNAs and SLE.

LncRNAs associated with SLE pathogenesis. Like microRNAs, a substantial number of LncRNAs have been acknowledged that can influence the pathogenesis of SLE. Also, the effect of different SNPs of these LncRNAs on the onset of SLE symptoms can be less effective. The impact of the LncRNAs network and their association with the MAPK pathway has also identified them as diagnostic biomarkers in SLE.
LncRNAs related to SLE disease.
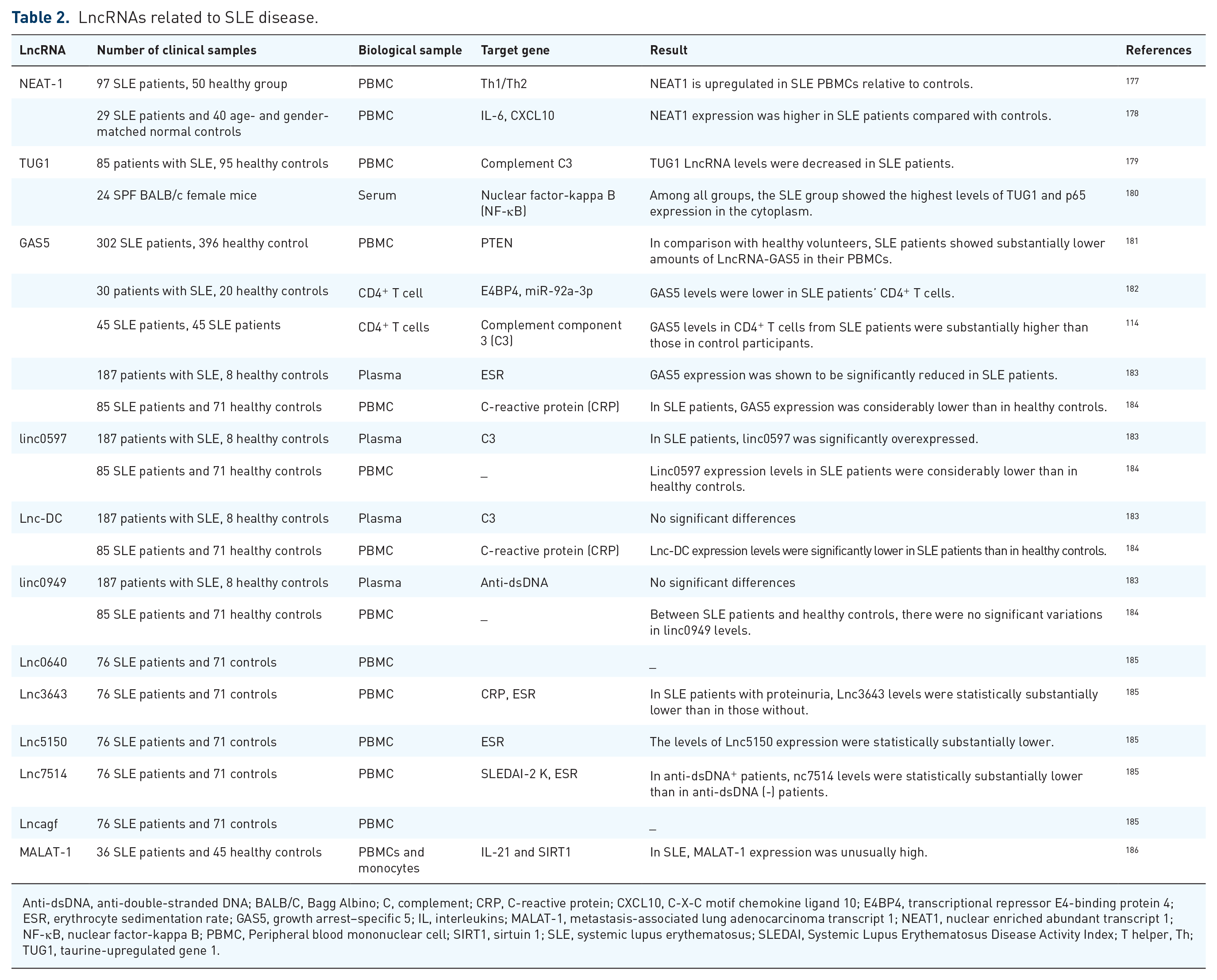
Anti-dsDNA, anti-double-stranded DNA; BALB/C, Bagg Albino; C, complement; CRP, C-reactive protein; CXCL10, C-X-C motif chemokine ligand 10; E4BP4, transcriptional repressor E4-binding protein 4; ESR, erythrocyte sedimentation rate; GAS5, growth arrest–specific 5; IL, interleukins; MALAT-1, metastasis-associated lung adenocarcinoma transcript 1; NEAT1, nuclear enriched abundant transcript 1; NF-κB, nuclear factor-kappa B; PBMC, Peripheral blood mononuclear cell; SIRT1, sirtuin 1; SLE, systemic lupus erythematosus; SLEDAI, Systemic Lupus Erythematosus Disease Activity Index; T helper, Th; TUG1, taurine-upregulated gene 1.
Nuclear paraspeckle assembly text 1
Nuclear paraspeckle assembly text 1 (NEAT1) is 3.2 kb ncRNA located on the 11q13.1 chromosome in the LncRNA subtypes. This LncRNA is produced by transcription from the familial tumor syndrome multiple endocrine neoplasia (MEN) type 1. Two isoforms, i.e. NEAT1-1 and NEAT1-2, have been discovered from this ncRNA that alternative 3′-end processing is responsible for regulating them. 187 Due to its high expression in non-neural cells and cell lines, it participates in several disorders, including cancers like non–small cell lung cancer, ovarian cancer, cervical cancer, hepatocellular carcinoma, etc. 188 Numerous studies have shown an association between NEAT-1 and SLE. 189 Jiang et al. evaluated the expression of NEAT1 in PBMC from patients with SLE and healthy individuals. NEAT1 was shown to be highly upregulated in the PBMC of SLE patients in this research. Consistent with this, a negative connection between its expression and Th1/Th2 balance was reported. Because the Th1/Th2 balance is crucial for the immune system to work correctly, altering it causes abnormal immune responses associated with SLE development. 177 NEAT-1 may have a role in lupus pathogenesis by modulating inflammatory pathways via MAPK. In this regard, individuals with SLE exhibited high levels of NEAT-1, mostly generated by monocytes, compared with controls. They can enhance SLE activity by increasing the number of monocytes in PBMC and the excessive production of inflammatory mediators such as cytokines and chemokines as a result of NEAT-1 overexpression. 178 Also, based on the findings of Dong et al. regarding the correlation between increased NEAT1 expression and granulocytic MDSCs (G-MDSC) in MRL/lpr mice in SLE, they have indicated that NEAT-1 upregulation activates the IFN-I signaling pathway of B cells through BAFF and promotes G-MDSCs, which cause SLE progression. 190
Taurine-upregulated gene 1
Taurine-upregulated gene 1 (TUG1) generates LncRNAs that interact with the polycomb repressor complex to regulate transcription epigenetically. This gene, located on chromosome 22q12.2, promotes cell proliferation and has been linked to cancers such as liver, osteosarcoma, glioma, and bladder. 191 TUG1 has a protective effect being linked to the NF-κB pathway because it can inhibit apoptosis and the production of inflammatory elements.192,193 Cao et al. performed pyrrolidine dithiocarbamate (PTDC) therapy to demonstrate the influence of NF-κB inhibition on renal damage in the SLE mouse model and also its association with TUG1. According to the results, treatment with PTDC increased TUG1 expression and downregulated P65 NF-κB. In brief, this highlights the negative association between TUG1 and NF-B p65 as the effect of these LncRNAs on SLE. 180 They also noted the downregulation of TUG1 expression in SLE patients’ PBMC compared with healthy individuals. 179
Growth arrest–specific 5
The gene encoding growth arrest–specific 5 (GAS5) is found on the 1q25.1 chromosome, which also contains numerous C/D box snoRNA (small nucleolar RNAs) genes in its introns. By interacting with the DNA-binding domain of the glucocorticoid receptor (GCR), this gene can imitate the glucocorticoid response. As a result, it plays a critical function in the androgen receptor, progesterone, and mineralocorticoid regulation. GAS5 may act as a tumor suppressor in diagnosis and management due to its role in suppressing proliferation and cell death. 194 GAS5 is implicated in malignancies and inflammatory illnesses, but its action on GCRs has also been linked to auto-immune disorders. Several GAS5 SNPs have been discovered, including rs145204276, rs2235095, rs6790, rs2067079, and rs1951625, which may be linked to SLE. The effect of the rs145204276 polymorphism on GAS5 expression in SLE patients is a notable example. Individuals with the rs145204276 ID+ genotype had more GAS5 expression than those without it, and they had fewer SLE symptoms, indicating that this polymorphism may be protective. 181 Qian and coworkers discovered a correlation between GAS5 expression changes and SLE based on previous investigations of GAS5 gene localization information. GAS5 was also shown to be downregulated in SLE patients’ T cells and plasma. GAS5 overexpression also inhibits miR-92a-3p, which controls adenovirus E4-binding protein 4 (E4BP4). As a result, it is critical in suppressing T lymphocyte activation in SLE patients. 182 However, during experiments on GAS5 alteration levels in SLE patients, Wu et al. 183 demonstrated a remarkable reduction in its expression in these patients compared with controls. Suo and colleagues surveyed the association of GAS5 with CD4+ T lymphocytes. Due to the increasing trend of GAS5 in patients with pleurisy, rash, anti-dsDNA, and low complement C3, it can be considered for the identification of SLE. In contrast to previous studies, this investigation emphasizes the high level of GAS in CD4+ T cells in SLE patients compared with healthy subjects. 114 Wu et al. 195 examined and analyzed ncRNAs, including GAS5, lnc0640, lnc3643, lnc6655, and lnc7074, as a competing endogenous RNA network (ceRNA). They also analyzed the association of GAS5, lnc0640, lnc3643, lnc6655, and lnc7074 with the pathogenesis of SLE through the inhibition of microRNAs. According to their results, these LncRNAs, which act through the MAPK pathway, might be utilized as novel and diagnostic biomarkers in SLE. 195
Lnc-DC
Lnc-DC is one of the most discussed ncRNA types expressed in DCs. Lnc-DC is capable of maturing DCs via STAT3 phosphorylation. In addition, the impact of Lnc-DC on monocytes causing them to differentiate into DCs can induce T lymphocyte activation. 196 Currently, Lnc-DC has become a central issue in the field of auto-immune disorders such as Sjögren, MS, and SLE. 197 In SLE, Lnc-DC has been identified as a significant biomarker. By examining this factor in SLE patients and normal individuals, Wu et al. 183 achieved a decrease in its expression in lupus patients. Li and colleagues investigated the expression levels of linc0597 Lnc-DC and GAS5 in SLE patients and found a reduction in their expression in SLE. It is worth mentioning that a change in the number of polymorphisms in the mentioned LncRNAs is not related to an enhanced risk of this disorder. 184
Myocardial infarction–associated transcript
Myocardial infarction–associated transcript (MIAT) is a common part of LncRNAs first identified in 2006 as the myocardial infarction gene. MIAT has a wide range of applications in cancer and another disease such as diabetic retinopathy, paranoid schizophrenia, and vascular dysfunction. 198 Even though MIAT is poorly known, it appears to play a critical role in auto-immune disorders. 199 Xu et al. explored the vast network between LncRNA, microRNA, and mRNA called ceRNA. They aimed to find an association between differentially expressed LncRNAs (DE-LncRNAs) in SLE pathogenesis. Interestingly, the influence of particular DE-LncRNAs like MIAT and NEAT1 by binding to mRNA and inhibiting the expression of the corresponding miRNA was observed in SLE. Consequently, LncRNAs, in addition to having a direct effect on SLE disease, also play a pivotal part in the disease by the ceRNA network. 200 Furthermore, investigations on MRL/lpr mice showed that MIAT exacerbates the progression of SLE. Hematoxylin and eosin (H&E) staining of the kidney tissues of MRL/lpr mice showed that MIAT, as a competitive inhibitor of miR-222, by increasing CFHR5 expression, degrades miR-222 and enhances SLE activity. 201
Metastasis-associated lung adenocarcinoma transcript-1
Metastasis-associated lung adenocarcinoma transcript-1 (MALAT-1), with a size of 12,820 bp, is an RNA gene affiliated with the LncRNA class. Principal physiological fundamental characteristics of MALAT-1 are alternative splicing and epigenetic changes. 202 However, it is pathologically involved in various diseases such as hyperglycemia and vulva squamous cell carcinoma. 203 MALAT-1 has recently been discovered to be involved in SLE as a novel inflammatory regulator. Increasing the unusual expression of this biomarker in SLE, especially in monocytes, and the destructive effects of MALAT-1 by the Sirt1 pathway, display its central role in the pathogenesis of SLE. MALAT-1 greatly increased IL-2 secretion in SLE patients compared with healthy individuals. 186
Other LncRNAs
There are additional ncRNAs that have not been investigated sufficiently but may have an impact on SLE. Wang et al. studied the expression pattern of five LncRNAs in the PBMC of SLE patients compared with the controls in the research. The abnormal expression of Lnc3643 and Lnc7514 was related to C-reactive protein (CRP) and positive anti-dsDNA, respectively. Accordingly, Lnc7514 expression in SLE patients was decreased with positive anti-dsDNA patients compared with negative anti-dsDNA. The analysis also revealed a correlation between CRP and the level of expression for Lnc3643. 185
The role of circRNAs in the progression of SLE
CircRNAs are a vast family of ncRNAs generated by reverse splicing of pre-mRNAs; meanwhile, their expression is also influenced by pre-mRNA transcription levels, which are determined by transcription factor activity and epigenetic modifications such as methylation and acetylation. 204 They have a unique structure consisting of a continuous covalently closed loop that lacks a 5′-cap and a 3′-poly A tail. The specific characteristics of circRNAs have made them resistant to degradation by exonuclease ribonucleases. 204 Viroids are one of the first circRNA molecules found in the last 40 years, with the exception that they are not produced by the reverse splicing mechanism. 205 Moreover, circRNAs are often synthesized in the nucleus, although they can also be detected in the cytoplasm and in the nucleus.206,207 Despite electron microscope observations of circRNAs in the cytoplasm of eukaryotic cells, they were regarded as ‘junk’ generated by improper splicing processes. 204 Considering the abundance of circRNAs in body fluids, recent studies have pointed to their enrichment and transport by exosomes. Therefore, the secretion of exosomes from pathological cells can cause an abnormal increase of certain circRNAs, which shows their importance as valuable biomarkers.208–210 A considerable number of studies have revealed that circRNAs have a role in a variety of disorders such as auto-immune diseases, indicating their potential as clinical biomarkers and therapeutic targets. 211 So far, using microarray and high-throughput sequencing techniques, several aberrantly expressed circRNAs have been identified in SLE. 212 Luo et al. 213 investigated the level of peripheral blood circRNAs in SLE patients with renal involvement (SLE + RI) and indicated that the level of hsa-circ-0082688 and hsa-circ-0008675 in SLE + RI patients was enhanced. Furthermore, new research in circRNA’s context has revealed that these RNAs absorb miRNAs like sponges and influence downstream target genes. The elevated expression of hsa-circ-0002003 in SLE may be involved in controlling immunological responses, according to the investigation of Lin et al. 214 On the other hand, Zhang et al. attempted to examine the regulation mechanism of circRNAs in T cells from SLE patients and revealed a correlation between the circRNA–microRNA–mRNA regulatory network. They established a circRNA–microRNA–miRNA network of 8 differentially expressed circRNAs (DECs), 4 miRNAs, and 13 mRNAs using several bioinformatics. Furthermore, they noted that their study may improve the diagnosis, treatment, and prognosis of SLE patients. 215 Zheng et al. stated that they extracted circRNAs for detection from SLE patients’ PBMCs. They discovered the aberrant expression of certain circRNAs in PBMCs. Thus, circRNAs may be beneficial as unique biomarkers for the diagnosis of SLE. 216 Considering the importance of circRNAs in the diagnosis of SLE, Miao et al. investigated 128 circRNAs in PBMCs of patients. According to the study’s findings, a reduction in circPTPN22 in patients with greater SLEDAI revealed a negative connection between these two variables. 217 Furthermore, another study demonstrated the function of hsa_circ_0045272 in the negative regulation of apoptosis and IL-2 secretion in SLE. 218 In addition, Zhang et al. 219 reported reduced expression of the hsa_circRNA_407176 and hsa_circRNA_001308 in the plasma and PBMCs of SLE patients. Generally, circRNAs can behave as novel biomarkers and essential regulators of SLE pathogenesis. However, information on the intricacies of the circRNA mechanism in SLE is inadequate.
Conclusion
The development of an immune response in the absence of risk factors might result in diseases such as asthma, allergies, or auto-immune diseases. In auto-immune disorders, the body mistakenly attacks the body’s own tissues. It has been shown that factors such as microorganisms and drugs may cause changes in the immune system in genetically predisposed individuals. Furthermore, these disorders can be influenced by a variety of factors such as inheritance, environment, hormones, and infectious agents; yet, despite the availability of many medications, no definite cure for this ailment has been established. SLE is a chronic disease with a wide variety of medical symptoms caused by complicated mechanisms such as incorrect apoptosis, dysregulation of the innate and adaptive immune systems, and complement activation. The pattern of clinical presentations, on the other hand, differs from person to person according to heterogeneity, emphasizing the relevance of genetic and environmental variables in this disease. Methylation, histone modification, and, in particular, ncRNAs are all factors in the pathogenesis of SLE. Despite their lack of participation in protein synthesis, ncRNAs play a crucial function in controlling gene expression due to their formation of a major portion of the human genome. Due to the significant role of ncRNAs in B-cell activation pathways and NF-κB pathways, as well as STAT-related pathways in SLE pathogenesis, new therapeutic goals have emerged as a result of research. In addition, greater attention should be paid to the interaction between microRNAs, LncRNAs, and circRNAs in this disorder, and more targeted therapies should be explored in this context. ncRNAs, particularly microRNAs, have been suggested to play a role in the identification of SLE complications, in addition to detecting healthy people from patients.