
Abstract
Keywords
Introduction
Chronic lymphocytic leukemia (CLL) is the most common adult leukemia in Western countries. The disease is very heterogeneous with some patients showing extremely slow progression while others proceed rapidly into advanced disease stages and require immediate treatment [Cramer and Hallek, 2011]. Despite considerable improvement of therapeutic strategies in the last decade, CLL remains incurable by conventional chemoimmunotherapies. The development of new treatment options remains an important goal.
Nonsteroidal anti-inflammatory drugs (NSAIDs) have been demonstrated to not only be useful in the treatment of pain, inflammation and fever, but also to possess a considerable antineoplastic effect [Thun et al. 2002; Shiff and Rigas, 1999]. As for most of the classical NSAIDs, use as an anticancer agent is limited by mainly gastrointestinal and cardiovascular side effects at required concentrations (for a review see Ng and Chan [2010]), so chemical modifications have been conducted. These modifications focused on the association of traditional NSAIDs with phospholipids, cyclodextrins, or chemical moieties that release gastroprotective mediators such as nitric oxide (NO) via an aliphatic, aromatic or heterocyclic spacer (for reviews see Abdel-Tawab et al. [2009] and Burgaud et al. [2002]). The pharmacokinetic and pharmacological properties of the final substance are largely dependent on the chemical structure of the spacer. NO-donating acetylsalicylic acid (NO-ASA) can be considered the classical NO-NSAID. Here, an aromatic spacer links the classical acetylsalicylic acid molecule to a NO-releasing moiety (–ONO2) [Baron, 2003]. Upon oral administration esterases rapidly cleave NO-ASA into ASA and the NO-releasing moiety linked to the spacer. Actual release of NO takes place in the subsequent metabolism of the spacer/NO-releasing complex [Wallace et al. 2002]. The general structure of NO-ASA enables the generation of several variants depending on the position of the –ONO2 group. Despite identical atomic composition these isomers may differ significantly in their pharmacological profiles, hence featuring considerably distinct drug activities. NO-ASA has three positional isomers depending on the position of the –ONO2 group in the benzene ring (para-, ortho-, or meta-position). The effects of these isomers have been studied mainly in colon cancer. While the para- and the ortho-isomers have been described to be largely equipotent in reducing colon cancer cell growth and survival in vitro and tumor formation in vivo (APCMin/+ mice), the meta-isomer was in all respects significantly less effective [Kashfi et al. 2005]. Also in the human T-cell leukemia cell line Jurkat the para-isomer showed a superior effect with regard to the reduction of cell growth compared with meta-NO-ASA (IC50 of 20 µM versus 200 µM, respectively) [Nath et al. 2005]. The same effect was seen in the breast cancer cell line MCF7 [Nath et al. 2009]. Up to now the precise mechanism of action of NO-ASA in general and the mechanistical differences of the isomers in particular are not completely understood. While a Cox-dependent effect has been excluded [Kashfi et al. 2005], it was noticeable that all cancers in which the para-isomer showed potent antineoplastic effects possessed aberrant activity of the Wnt/β-catenin/TCF/Lef-1 signaling pathway. Indeed para-NO-ASA has been demonstrated to disrupt the transcriptionally active β-catenin/TCF/Lef-1 complex in colon cancer [Gao et al. 2005; Nath et al. 2003] and breast cancer [Nath et al. 2009], thereby contributing to apoptosis induction. Also in CLL Lef-1 was described to be one of the most overexpressed genes [Jelinek et al. 2003]. Furthermore, the Wnt/β-catenin/TCF/Lef-1 signaling pathway is aberrantly active [Lu et al. 2004] and its therapeutic inhibition has been demonstrated to induce apoptosis in primary CLL cells in vitro and reduce tumor growth in a xenograft mouse model [Gandhirajan et al. 2010]. We have recently demonstrated, confirming data obtained in solid tumors, that para-NO-ASA potently induced apoptosis in vitro in primary CLL cells and prevented tumor growth in a xenograft CLL-like mouse model [Razavi et al. 2011]. Also here β-catenin cleavage was involved in apoptotic cell death. Nevertheless, the exact mechanism of action of para-NO-ASA in CLL is not clear up to date. Further, no previous research has described whether the positional isomerism of NO-ASA is of significance for its antineoplastic effects in CLL.
In the present study, we investigate the impact of positional isomerism on survival of primary CLL cells in vitro and on tumor growth in a xenograft mouse model in vivo using para- and meta-NO-ASA. Further, the capacity of the positional isomers to interfere with the aberrantly active β-catenin/Lef-1 signaling pathway is studied.
Results
Para-NO-ASA but not meta-NO-ASA reduces CLL cell survival within a short treatment period of 12 hours
NO-ASA has been demonstrated to possess strong antineoplastic efficacy in several solid tumors, mainly colon and pancreatic cancers. In most studies NO-ASA inhibited cell growth or induced apoptosis rapidly, usually within the first 4 hours of treatment. We have previously shown that in primary CLL cells para-NO-ASA induced apoptosis as early as 6 hours of drug exposure at a low concentration of 10 µM. As positional isomerism has been described to be highly relevant for drug efficacy we intended to compare the two positional isomers para- and meta-NO-ASA (Figure 1) for their impact on CLL cell survival. As assessed by annexin V-FITC/PI staining, the para-isomer rapidly induced apoptosis within the first 6 hours of treatment with no significant increase of apoptosis thereafter. At equimolar doses the meta-isomer did not reduce CLL cell survival within the studied time frame of up to 12 hours (96.4% ± 2.7% surviving annexin V-FITC/PI double negative cells) (Figure 2A).
Chemical structure of para- and meta-isomers of nitric-oxide-donating acetylsalicylic acid (NO-ASA). Survival reduction by para- and meta-isomers of nitric-oxide-donating acetylsalicylic acid (NO-ASA). (A) Primary chronic lymphocytic leukemia (CLL) cells (n = 3) were treated with 10 µM of either isomer for 3, 6, 9, or 12 hours. Survival was assessed by annexin V-FITC/PI staining measured on FACS Canto. Para-NO-ASA reduced survival (annexin V-FITC/PI double-negative cells) within the first 3 hours to approximately 50%. At 6, 9 and 12 hours survival was significantly reduced by para-NO-ASA when compared with meta-NO-ASA (p = 0.0461, p = 0.0252, and p = 0.0212, respectively). (B) Primary CLL cells (n = 7) were treated with meta- and para-NO-ASA at concentrations ranging from 0.1 µM to 100 µM for an extended treatment period of 24 hours. ATP content was analyzed as an indirect measure of viability by CellTiter-Glo luminescence cell viability assay. For para-NO-ASA treatment an LC50 of 4.34 µM could be determined. Meta-NO-ASA only marginally reduced survival at high concentrations of 100 µM (83.0 ± 3.1% surviving cells). (C) Experimental set up was as described for (B). Survival was assessed by annexin V-FITC/PI negativity on FACS Canto (n = 4). Calculated LC50 for para-NO-ASA was 8.93 µM. Meta-NO-ASA reduced survival marginally to 86.0 ± 4.3% at 100 µM.

Para-NO-ASA but not meta-NO-ASA reduces CLL cell survival at low micromolar concentrations within a prolonged treatment period of 24 hours
Meta-NO-ASA has been described to generally act less rapidly than para-NO-ASA. That is why we extended the treatment period to 24 hours. Furthermore, several concentrations were tested ranging from 0.1 µM to 100 µM. While para-NO-ASA treatment resulted in a concentration-dependent decrease of both ATP levels (as an indirect measure of apoptosis; Figure 2B) as well as annexin V-FITC/PI double negativity (Figure 2C), the meta-isomer neither affected CLL cell ATP content nor annexin V-FITC/PI status after 24 hours of treatment. Further, caspase activation and PARP cleavage were monitored upon treatment with both isomers as indicators for apoptosis induction. As we have shown previously, para-NO-ASA considerably induced cleavage of caspases and PARP after 24 hours in a concentration-dependent manner. Here, we demonstrate an activation of the initiator caspase 9, effector caspases 3 and PARP cleavage upon treatment with the para-isomer, while at equimolar dose the meta-isomer did not have any impact on caspase or PARP cleavage in primary CLL cells (Figure 3).
Caspase activation by para- and meta-isomers of nitric-oxide-donating acetylsalicylic acid (NO-ASA). Primary chronic lymphocytic leukemia (CLL) cells were incubated with meta (m)- or para (p)-NO-ASA at 20 µM for 24 hours. Cells were lysed and subject to an immunoblotting procedure using antibodies targeting caspase 9, 3, PARP, cleaved (cl.) caspase 9, 3 and cleaved (cl.) PARP. β-actin functioned as a control. Para-NO-ASA reduced protein levels of intact caspases 9 (A) and 3 (B) and PARP (C). At the same time cleaved, hence activated caspases 9 (A), 3 (B) and PARP (C) levels were significantly increased (p = 0.0152, p = 0.0471, and p = 0.0256, respectively). Meta-NO-ASA did not alter caspase 9 (A), 3 (B) or PARP (C) protein levels. Bar diagrams represent corresponding densitometric band intensities for three independent experiments. p-values were calculated using densitometric measures by two-tailed Student’s t-test with Welch correction.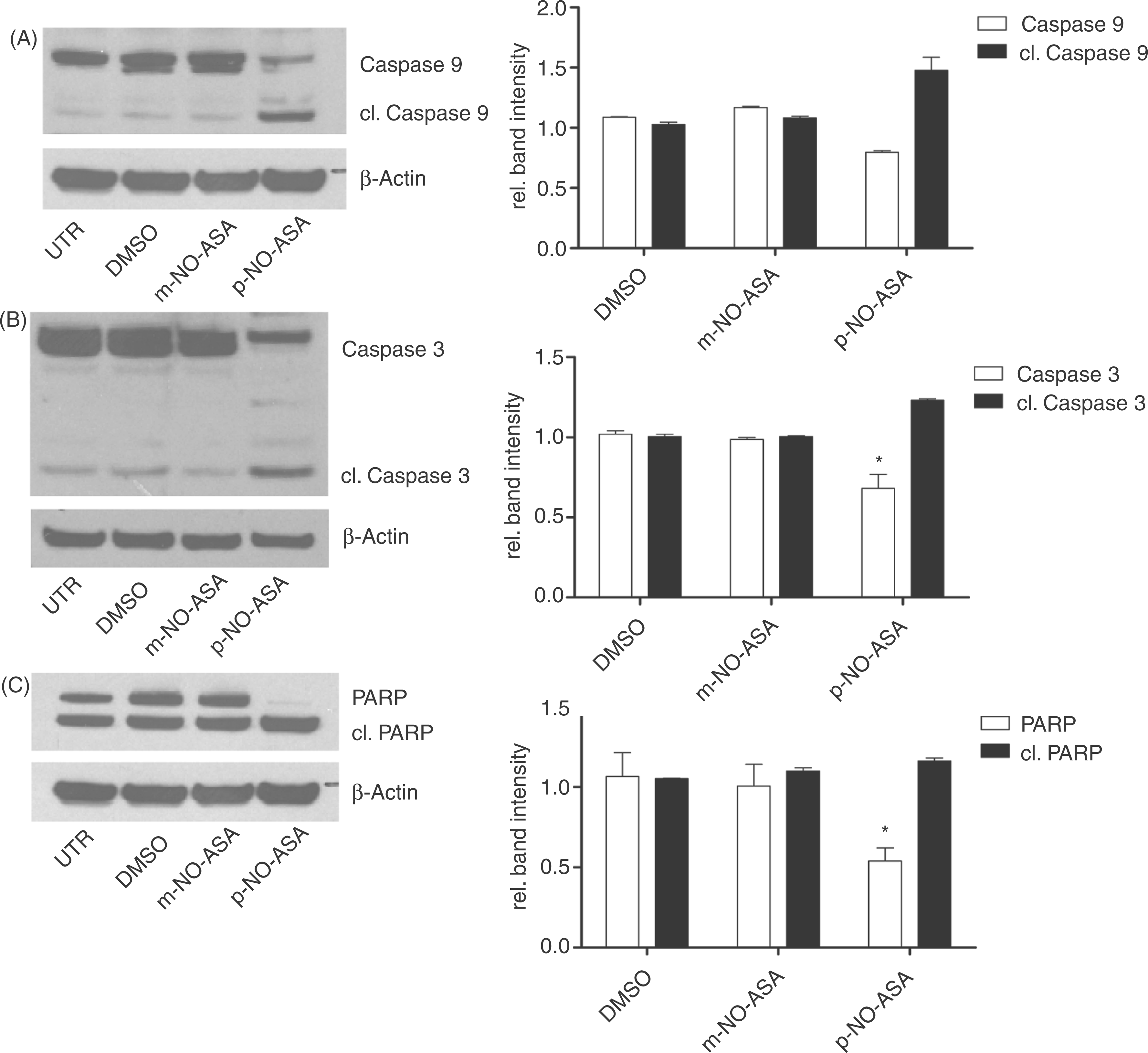
Lef-1 protein levels are significantly reduced by para-NO-ASA, but remain completely unchanged by meta-NO-ASA
NO-ASA action has been described to involve blockage of the β-catenin/TCF/Lef-1 transcriptional complex in colon cancer cells. As this signaling cascade was also described to be aberrantly active in CLL [Lu et al. 2004] and its inhibition efficiently leads to apoptosis in CLL cells [Gandhirajan et al. 2010], we were interested in studying the impact of different NO-ASA isomers on this signaling pathway in CLL cells. Neither protein levels of β-catenin and Lef-1, nor the known target gene of the transcriptionally active β-catenin/TCF/Lef-1 complex CyclinD1 were altered upon treatment with the meta-isomer. At the same time, para-NO-ASA potently induced β-catenin cleavage, thereby entailing an almost complete reduction of its uncleaved intact form. Going along with β-catenin downregulation, Lef-1, which is activated in a feedback loop manner, as well Cyclin D1 were downregulated upon treatment with the para-isomer (Figure 4).
Involvement of β-catenin and known Lef-1 targets in nitric-oxide-donating acetylsalicylic acid (NO-ASA) action. Primary chronic lymphocytic leukemia (CLL) cells were incubated with meta (m)- or para (p)-NO-ASA at 20 µM for 24 hours. Cells were lysed and subject to an immunoblotting procedure using antibodies targeting β-catenin, cleaved (cl.) β-catenin, Lef-1 and Cyclin D1. β-actin functioned as a control. (A) Para-NO-ASA induced β-catenin cleavage (p = 0.0319). Meta-NO-ASA did not alter levels of intact β-catenin or result in β-catenin cleavage. (B) Lef-1 protein was significantly reduced upon para-NO-ASA treatment (p = 0.0367). Also Cyclin D1 was inconsistently downregulated (p = 0.0681). Neither Lef-1 nor Cyclin D1 protein levels were altered by meta-NO-ASA treatment. Bar diagrams represent corresponding densitometric band intensities for three independent experiments. p-values were calculated using densitometric measures by two-tailed Student’s t-test with Welch correction.
Caspase inhibition does not have an influence on the impact of meta-NO-ASA on CLL cell survival
We have previously shown that para-NO-ASA requires caspase activity in order to reduce CLL cell survival. Also β-catenin cleavage and downregulation of β-catenin/Lef-1 target genes was caspase-dependent. To further investigate the differences of the para- and meta-isomers of NO-ASA on CLL cell survival, we blocked caspase activity using a pan-caspase inhibitor prior to NO-ASA treatment. While para-NO-ASA activity was largely diminished, caspase inhibition did not alter the meta-NO-ASA-mediated effect on CLL cell survival (Figure 5A). Going along with survival reduction the highly upregulated anti-apoptotic proteins Bcl-2 and XIAP were significantly downregulated upon treatment with para-NO-ASA, while equimolar doses of meta-NO-ASA did neither reduce Bcl-2 nor XIAP protein levels (Figure 5B). Also here para-NO-ASA-induced downregulation was highly dependent on caspase activity.
Impact of caspases on nitric-oxide-donating acetylsalicylic acid (NO-ASA)-mediated effect on apoptosis induction. Primary chronic lymphocytic leukemia (CLL) cells were incubated with 50 µM pan-caspase inhibitor prior to treatment with meta- or para-NO-ASA at 20 µM. (A) Survival was assessed by annexin V-FITC/PI staining measured on FACS Canto. Caspase inhibition completely prevented para-NO-ASA-induced survival reduction (p = 0.0032 and p = 0.1715 for survival after treatment relative to dimethylsulfoxide (DMSO) treatment without and with caspase inhibition, respectively). Meta-NO-ASA was ineffective independent on status of caspase activity. (B) Upon treatment cells were lysed and subject to an immunoblotting procedure using antibodies targeting Bcl-2 and XIAP. β-actin functioned as a control. Bcl-2 (p = 0.0235) and XIAP (p = 0.0070) were downregulated by para-NO-ASA treatment. Caspase blockage prevented Bcl-2 (p = 0.2635) downregulation. XIAP downregulation was still significant when caspases were blocked (p = 0.0103). Both anti-apoptotic proteins remained unaltered upon meta-NO-ASA treatment independent on the caspase activity status. Bar diagrams represent corresponding densitometric band intensities for three independent experiments. p-values were calculated using densitometric measures by two-tailed Student’s t-test with Welch correction.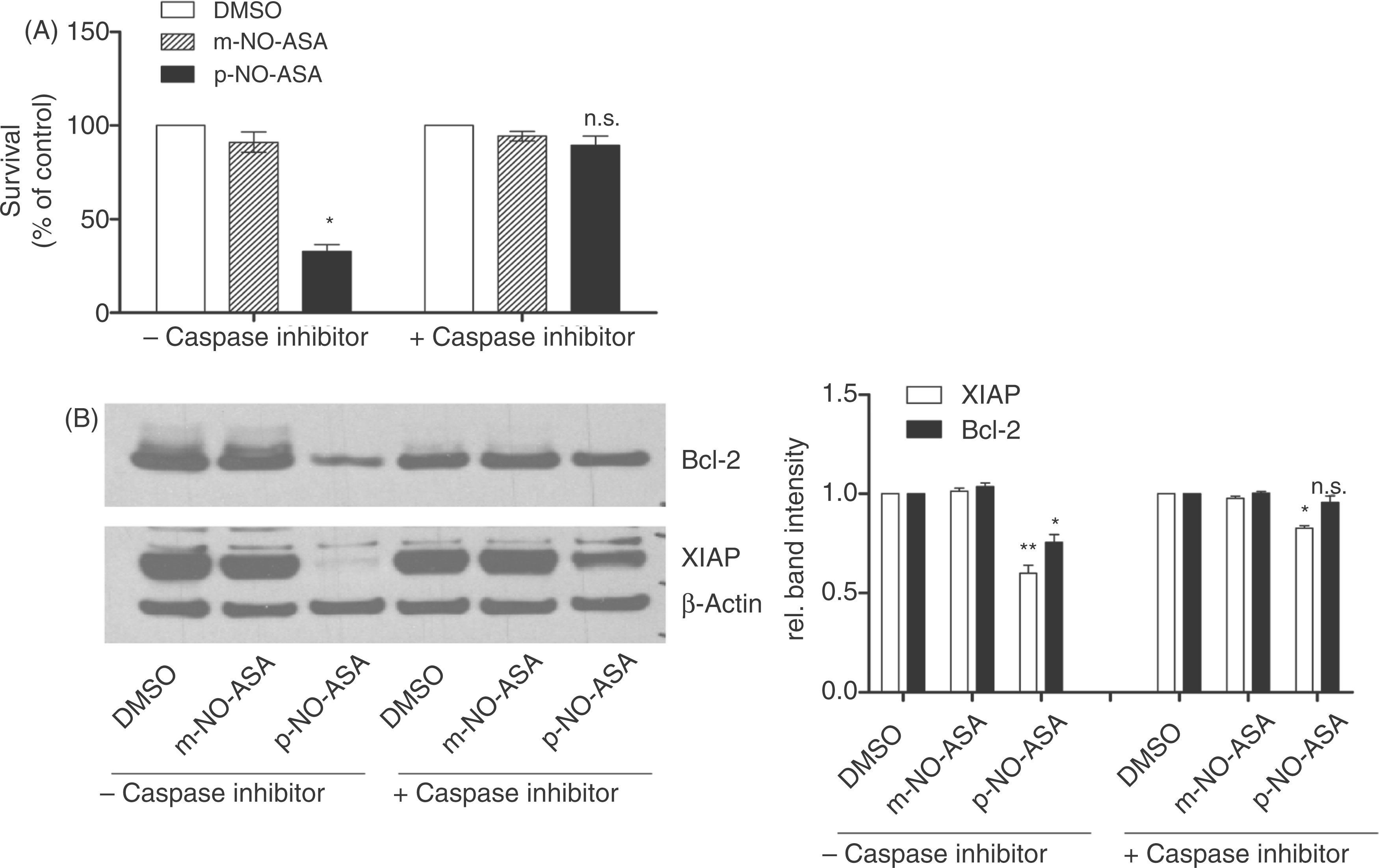
Meta- and para-NO-ASA possess significantly differential effects on tumor growth in a JVM3 xenograft mouse model
To investigate whether positional isomerism also impacts the in vivo efficacy of NO-ASA, the JVM3 cell line, which is a B-lymphoid cell line with some CLL-like features, was used for engraftment of a subcutaneous tumor in nude mice. Mice either received meta-NO-ASA, para-NO-ASA or vehicle control per os (p.o.) via gavage daily for a total of 21 days. Also here a drastic difference between the two isomers on their impact on tumor growth was obvious. While para-NO-ASA induced a significant growth inhibition as early as 9 days of treatment and reached a final tumor inhibitory rate of 84.3% (tumor volume 775.4 ± 219.6 mm3 versus 128.7 ± 27.6 mm3 for vehicle- and para-NO-ASA-treated group, respectively; p = 0.0091), meta-NO-ASA did not significantly affect tumor growth compared with vehicle-treated control at any given time point. As illustrated in Figure 6, we observed a limited, not statistically significant inhibitory effect of meta-NO-ASA on tumor growth with an IRmax of 47.9%.
Efficacy of meta- and para-isomers of nitric-oxide-donating acetylsalicylic acid (NO-ASA) on tumor growth in a chronic lymphocytic leukemia (CLL)-like xenograft mouse model. Xenografted nude mice were treated daily for 22 days with 100 mg/kg body weight of NO-ASA isomers or vehicle control (n = 10 per group). Tumor volume was determined using a standard caliper. Para-NO-ASA significantly inhibited tumor growth as early as 9 days into treatment. Tumor volume remained constant until the end of treatment (128.7 ± 26.2 mm3). Tumor growth in meta-NO-ASA-treated animals was not reduced compared with vehicle-treated control animals until the end of treatment (406.9 ± 86.1 mm3 versus 775.4 ± 208.4 mm3 for meta-NO-ASA and vehicle control, respectively). p-values were calculated by two-tailed Student’s t-test with Welch correction.
Discussion
Our findings underline the relevance of positional isomerism for the antineoplastic effect of NO-ASA in CLL. In this study, the meta-isomer was much less effective than the para-isomer. A significantly higher potency of para-NO-ASA over meta-NO-ASA has been described before, for example in colon cancer [Kashfi et al. 2005; Williams et al. 2004], pancreatic cancer cell lines [Rosetti et al. 2006], or leukemic Jurkat cells [Nath et al. 2005]. Subsequently, a crucial role for aromatic resonance interaction can be suggested. In general it has been speculated that a major requirement for the cytotoxic effect is the presence of an aromatic ring substituted with both a masked phenolic group and a para-methylene group allowing the generation of a quinone methide [Dunlap et al. 2009, 2007; Hulsman et al. 2007]. Owing to the structural position of the −ONO2 group, the meta-isomer is not capable of generating such a toxic methide. Quinones are known to exhibit strong cytotoxic properties in cancers [Dunlap et al. 2007; Bolton et al. 2000]. There are various known mechanisms for how quinones cause this effect. They are capable of alkylating both proteins and DNA, which can severely disturb physiological processes of the cell. Furthermore, quinones can provoke the generation of reactive oxygen species (ROS), resulting in oxidative stress [Bolton et al. 2000]. Gao and colleagues showed that oxidative stress induced by binding of the NO-ASA spacer to glutathione lead to activation of the intrinsic apoptotic pathway in a colon adenocarcinoma cell line [Gao et al. 2005]. Also Sun and colleagues demonstrated generation of oxidative stress going along with induction of cell death upon NO-ASA treatment (para- and ortho-isomer). Pretreatment with the antioxidant N-acetyl-cysteine (NAC) reduced ROS levels going along with improved survival upon NO-ASA treatment [Sun et al. 2009], thereby strongly suggesting ROS generation to be implicated in the mechanism of NO-ASA action.
Recently, Hua and colleagues investigated the differences between the two NO-ASA isomers para and meta for their ability to manipulate major intracellular signaling pathways in colon cancer [Hua et al. 2009]. They described para-NO-ASA to have a more rapid effect and to predominantly alter JNK1/2, whereas the meta-isomer exerts its major effect via ERK1/2 and Akt signaling. Since ROS can activate JNK1/2 signaling [Lo et al. 1996], a potential mechanism of apoptosis induction by para-NO-ASA in CLL could be the initiation of oxidative stress by quinone formation, followed by activation of JNK1/2 signaling and a subsequent activation of caspase-mediated apoptosis. Cleavage, hence inactivation of β-catenin through active caspase 3, reduces the expression of prosurvival target genes, preventing the cell from antagonizing the apoptotic process. Nevertheless, this suggestion has to be taken with care due to several reasons. In the study of Hua and colleagues, the effects on signaling pathways were highly dependent on concentration and treatment duration. Further, the study of Hua and colleagues was conducted in colon cancer cells and cannot be simply extrapolated to CLL cells, where all of the mentioned signaling pathways participate in survival of the malignant cells.
As CLL cells are highly dependent on their microenvironment and their actual in vivo situation is only partially resembled by in vitro cell culture, we tested both substances in an in vivo setting utilizing xenografted nude mice. Despite the ineffectiveness of meta-NO-ASA in vitro, a potential effect of meta-NO-ASA in vivo could not be excluded completely, as some substances require microenvironmental factors in order to properly carry out their effects. Although a slight tumor growth inhibition was observed after 21 days of treatment with meta-NO-ASA, statistical significance was not reached. This reduced average tumor volume was mainly due to the exclusion of one animal at day 19 which featured a heavy tumor burden with a tumor size exceeding the ethical limit of 1000 mm3. Without exclusion of this animal, the ineffectiveness of meta-NO-ASA would be even more pronounced. In contrast, para-NO-ASA effectively prevented tumor growth with an IR of almost 90%, which was also shown earlier [Razavi et al. 2011]. Hence, it can be clearly stated that the localization of the NO-releasing moiety at the aromatic spacer part has an essential impact on the antineoplastic activity of NO-ASA in this setting, suggesting para-NO-ASA, but not meta-NO-ASA as potential therapeutic option in CLL. However, with regards to a feasible clinical application the toxicity profile of para-NO-ASA remains to be determined.
In relation to translation of this experiment to the human organism, it has to be kept in mind that the xenograft model used (solid tumor, predominantly subcutaneous, immune-suppressed mice) features limitations with respect to the actual in vivo situation (blood, bone marrow, lymphatic tissue).
Concluding from this study, we could not only emphasize the critical difference between the para- and the meta-isomers for their antineoplastic effect, but also contributed to the determination of the mechanistical background of NO-ASA action in CLL.
Material and methods
Patients, sample preparation and cells
Clinical characteristics of CLL patients.

The JVM-3 cell line was purchased from the German Collection of Microorganisms and Cell Cultures (DSMZ) and cultured in RPMI1640, 20% FBS, 1% penicillin/streptomycin under above-mentioned standard conditions.
Drugs
Para-NO-ASA (2-(acetyloxy)-4-[(nitrooxy)methyl] phenyl ester, benzoic acid, NCX-4040) and meta-NO-ASA (2-(acetyloxy)-3-[(nitrooxy)methyl]phenyl ester, benzoic acid, NCX-4016) were purchased from Cayman Chemical Company (Figure 1). Drugs were stored at –20°C as stocks of 10 mM in dimethylsulfoxide (DMSO). DMSO was used as vehicle control for all in vitro experiments. For in vivo experiments the compounds were suspended homogenously in 1% carboxymethyl cellulose (CMC; Sigma-Aldrich). For inhibition of caspases, BD™ ApoBlock was used (BD Pharmingen).
Survival assessment
The Annexin V-FITC/PI Apoptosis Detection Kit (BD Pharmingen) was used to determine the percentage of nonapoptotic cells following the manufacturer’s instructions. Annexin V/PI double-negative cells were considered nonapoptotic cells. Survival was displayed relative to DMSO control. Lethal concentration 50 (LC50) was determined with Graph Pad Prism Software.
Further, the CellTiter-Glo luminescence cell viability assay (Promega GmbH) was used following the manufacturer’s instructions. This assay is used as an indirect measure of viability by means of the cells’ ATP content.
Immunoblot analysis
Following treatment for 24 hours primary CLL cells were lysed using mammalian protein extraction reagent (M-PER; Pierce). Protein separation and transfer was done using NuPage™ Novex 4–12% bis-tris gels and nitrocellulose membrane (Invitrogen GmbH). Primary antibodies targeting Bcl-2 (catalog number 610539), XIAP (catalog number 610762), Cyclin D1 (catalog number 554181), β-catenin (catalog number 610154) and β-actin (catalog number 612656) were from BD Bioscience. Primary antibodies targeting Lef-1 (catalog number 2230), caspase 9 (catalog number 9501), cleaved caspase 9 (catalog number 9501), caspase 3 (catalog number 9662), cleaved caspase 3 (catalog number 9662), poly(ADP-ribose) polymerase (PARP) (catalog number 9542) and cleaved PARP (catalog number 9542) were from Cell Signaling Technology. Secondary horseradish peroxidase (HRP)-labeled antibodies were obtained from DAKO GmbH. Antibody binding was detected using Amersham ECL™ Western blotting detection reagents (GE Healthcare UK). Band intensities were analyzed using ImageJ software.
Xenografted nude mice
Xenograft nude mouse model was established as described by Loisel and colleagues [Loisel et al. 2005]. Briefly, JVM-3 cells were injected subcutaneously into the right flank of 4-week-old CD-1 female nu/nu mice (Charles River Laboratories). Mice were divided into three groups comprising 10 animals each with an average tumor volume of 60 mm3. Substances or vehicle control were administered daily via oral gavage at 100 mg/kg bodyweight for a total period of 22 days. Tumor volume was evaluated every third day, using a standard vernier caliper, starting with the first day of treatment (day 0).
Statistical analysis
Statistical analysis was performed using GraphPad Prism™ (GraphPad Software) for all experiments. Data are given as mean + standard error of the mean (SEM). Unpaired Student’s t-test was used with two-tailed comparison of the differences between test values and controls. p-values <0.05 were considered significant.
Footnotes
Acknowledgement
We thank Samir Tawadros for his assistance with animal experiments.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of interest statement
The authors declare that there are no potential conflicts of interest.