
Abstract
Objective:
Chondroitin sulfate is the major sulfated polysaccharide attached to the core protein, aggrecan, in the hyaline cartilage matrix. Sulfation of the cartilage matrix polysaccharide is vital for normal matrix integrity and compressive stiffness of the tissue and is therefore crucial to normal cartilage formation and consequently to endochondral ossification. Several forms of chondrodysplasia, a condition resulting in clear macroscopic deficiencies in the mechanical properties of the cartilage and characterized by reduced levels of sulfate, have been identified in both human beings and animals.
Design:
In this study, the authors used capillary electrophoresis to investigate the sulfation state of extracted chondroitin sulfate polymers.
Results:
Significantly, cartilage from affected sheep had a lower ratio of the chondroitin-derived enzymatically liberated disaccharides Δdi-mono4S to Δdi-mono6S, demonstrating reduced levels of chondroitin 4-sulfate, but not chondroitin 6-sulfate, in chondrodysplastic sheep compared to age-matched controls at all ages measured.
Conclusion:
This supports the hypothesis that a difference in chondroitin sulfate disaccharides is detectable in affected newborn lambs prior to the development of lesions.
Animals Used and Samples Collected
Several forms of chondrodysplasia, a condition resulting in clear macroscopic deficiencies in the mechanical properties of the cartilage and characterized by reduced levels of sulfate, have been identified in both human beings and animals.1-7 In this work an entire humerus was collected from sheep that had undergone postmortem examination and was stored at −20°C for 6 to 24 months. Near-full-thickness articular cartilage shavings (10-15 mg) were harvested from the central humeral heads of 8 animals of various ages. Due to the age-related differences in articular cartilage thickness and composition, sulfation profiles are expected to vary with age of the animal: an issue that we attempted to ameliorate by using age-matched controls. Samples 1 and 2 were from obligate affected newborn lambs (the form of chondrodysplasia investigated in this study is a heritable disorder with a simple autosomal recessive mode of inheritance, so although there are no macroscopic or microscopic lesions identifiable in newborn lambs, lambs with both parents affected by the disorder were considered to be obligately affected), samples 3 and 4 were from control newborn lambs, sample 5 was from a 1-month-old lamb with severe chondrodysplasia, sample 6 was from a 1-month-old control lamb, sample 7 was from a mature chondrodysplastic ram, and sample 8 was from a mature control sheep. The animals used as controls were not taken from the same back-cross breeding trial that produced the affected animals for the study. This was because although lambs with both parents affected could be considered obligately affected, there was no way to confidently determine nonaffected newborn lambs. The control animals were genetically unrelated but age matched and from the same farm. Although unfortunately the breed was not recorded in this instance, the results reported herein are highly suggestive and should stimulate further work.
Chondroitin Disaccharide Extraction
The method for extraction of chondroitin disaccharides from articular cartilage was adapted from similar previous studies.8,9 Briefly, the glycosidic bond linking sulfated glycosaminoglycans to aggrecan was cleaved by incubation with 100 µL of 0.5M NaOH per mg of sample at 4°C, with agitation overnight. The NaOH was then removed by minidialysis against distilled water in 3.5-kDa molecular weight cutoff dialysis cassettes (Slide-A-Lyzer, Pierce Biotechnology, Rockford, IL) for 48 hours at 4°C. Samples were then centrifuged at 13,000 g for 3 minutes and the supernatant freeze-dried and then rehydrated in 25 µL deionized water. In order to digest the extracted polysaccharides into disaccharides, 10 µL of resuspended supernatant was incubated with 10 µL chondroitinase ABC (from Proteus vulgaris, Sigma-Aldrich Corp., St. Louis, MO; 0.05 mU/µL in 200 mM Tris base at pH 7.3) for 1 hour at 37°C. Digested samples were filtered through 3-kDa nominal molecular weight limit centrifugal filter units (Microcon ultracel YM-3, Amicon Bioseparations, Millipore, Billerica, MA) at 13,000 g for 90 minutes, and the filtrate containing the disaccharides was further diluted with 20 µL distilled water.
Capillary Electrophoresis
Experiments were performed using an automated capillary electrophoresis (CE) system (HP 3D), equipped with a diode array detector, based on a previously reported method.10,11 Briefly, electrophoresis was carried out at reverse polarity and 25°C in a fused silica capillary of internal diameter of 50 µm and a total length of 46.5 cm. Phosphate buffer (15 mM) at pH 3.0 was used as a CE background electrolyte, with UV detection of the unsaturated bond resulting from enzymatic digestion absorbing at 232 nm. Samples were loaded hydrodynamically at 5,000 Pa for 10 seconds and typically electrophoresed across a potential difference of 20 kV. Standard samples of 2-acetamido-2-deoxy-3-O-(4-deoxy-α-L-threo-hex-4-enopyranosyluronic acid)-4-O-sulfo-D-galactose (known as chondroitin 4-sulfate disaccharide, chondroitin sulfate A disaccharide, or Δdi-mono4S) and 2-acetamido-2-deoxy-3-O-(4-deoxy-α-L-threo-hex-4-enopyranosyluronic acid)-6-O-sulfo-D-galactose (known as chondroitin 6-sulfate disaccharide, chondroitin sulfate C or Δdi-mono6S) (Sigma-Aldrich Corp.) were used to identify ionic species and to construct standard curves for quantification. All samples were run at least 4 times and the peaks integrated using HP Chemstation software (Agilent Technologies, Santa Clara, CA). Where a particular disaccharide exhibited a double peak, both anomeric forms were pooled. Normalized 12 peak areas were subsequently used, via a calibration of absorptivity using the standard samples, to produce concentrations in mg/L.
Analysis
In order to compensate for slight differences in injection volume, the comparison of the results from the different samples was carried out by taking the concentration of Δdi-mono4S and dividing by the concentration of Δdi-mono6S, giving a ratio for each sample. Statistical analyses were performed using S-Plus 8.0 student edition (Insightful Corp., Seattle, WA, 2007). The analysis of variance (ANOVA) model was as follows: ratio of Δdi-mono4S to Δdi-mono6S ~ Age + Status, where Age was the age of the animal from which cartilage was taken, and Status was the presence or absence of chondrodysplasia. The Student t test was used to compare individual animals.
A typical electropherogram is shown in Figure 1 , demonstrating peaks caused by Δdi-mono4S, Δdi-mono6S, and a peak at the expected location for unsulfated chondroitin disaccharide, as demonstrated by Karamanos et al. 11 Double peaks were routinely observed, owing to the different anomeric forms of the hexosamine present at the reducing terminal of the disaccharide. Ten milligrams of each sample was run and mean concentrations of disaccharide for each of the tested animals are listed in Table 1 , expressed as ng/10-mg sample. Concentrations of Δdi-mono4S were markedly lower in cartilage from chondrodysplastic sheep of all ages compared with controls, but there was no such difference in Δdi-mono6S concentrations. The ratios of Δdi-mono4S to Δdi-mono6S were also lower in chondrodysplastic sheep than in unrelated control animals in all age groups (P ≤ 0.0001; Fig. 2 ). In addition, the ratio of Δdi-mono4S to Δdi-mono6S was markedly lower in the mature control than in the younger controls and in the mature affected ram than in the younger affected lambs (P ≤ 0.0001). The concentration of unsulfated chondroitin disaccharide did not vary significantly with disease status.
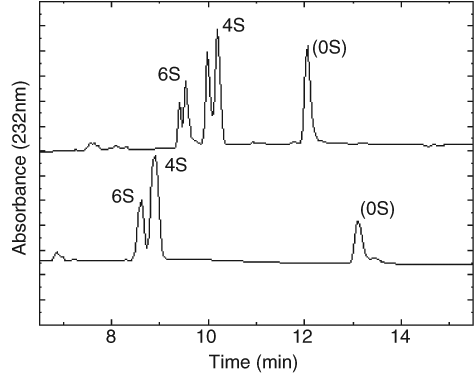
Typical electropherogram of chondroitin disaccharides from ovine articular cartilage. (
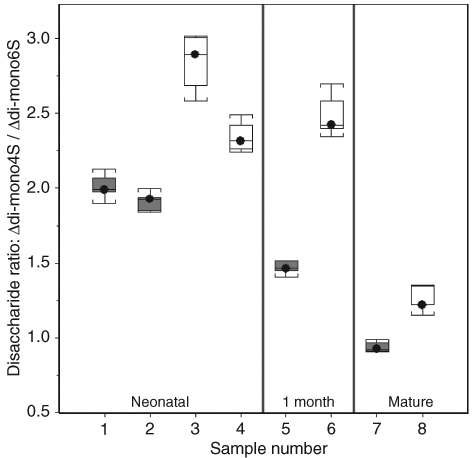
Ratios of Δdi-mono4S to Δdi-mono6S in chondrodysplastic and control Texel sheep of different ages. Gray plots are chondrodysplastic animals; white plots are control animals. The ratios are lower in affected animals of all ages (P ≤ 0.0001).
Concentrations of Chondroitin Sulfate Disaccharides from Chondrodysplastic and Control Texel Sheep of Varying Ages
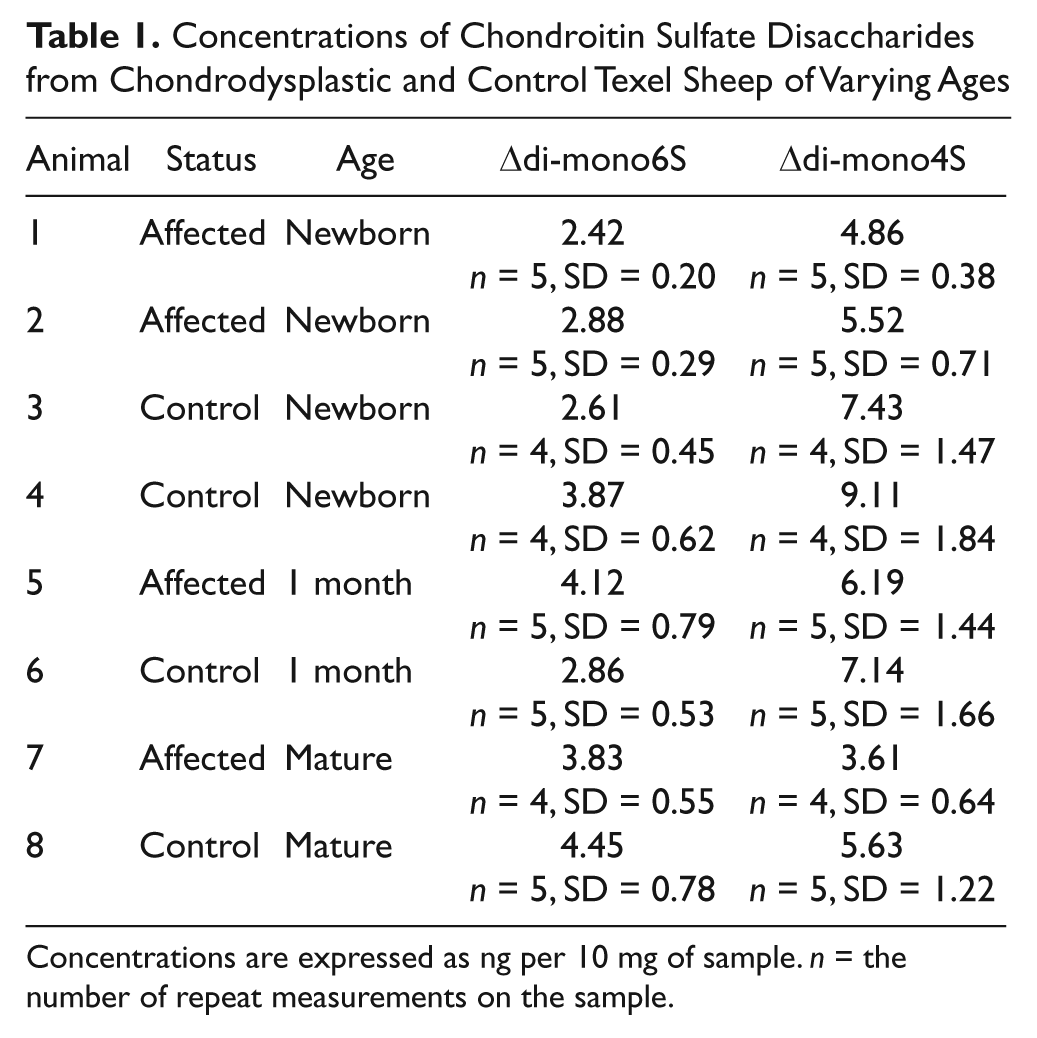
Concentrations are expressed as ng per 10 mg of sample. n = the number of repeat measurements on the sample.
Although the ratio of C4S to C6S decreases with age, alterations due to degenerative joint disease are variable, with some previous reports of no change and other reports of an increased ratio of C4S to C6S. The older affected animals in this study demonstrated angular limb deformities with resultant misloading of joints, and therefore it is possible that the change in chondroitin sulfation was a reflection of secondary degenerative joint disease. The change in sulfation, however, was a decrease rather than an increase in the ratio of C4S to C6S, and the presence of an altered sulfation profile in the newborn lambs that showed no gross or microscopic joint lesions supports a primary alteration in chondroitin sulfation.
A decreased ratio of Δdi-mono4S to Δdi-mono6S has been recorded in human chondrodysplasias resulting from sulfate transport disorders due to reduced activity of the diastrophic dysplasia sulfate transporter (DTDST) 13 and in mutations in the gene encoding chondroitin 4-sulfotransferase-1, CHST11. 14 This finding indicates that CHST11, which maps to ovine chromosome 3 at approximately 221.4cM, is potentially a candidate gene for involvement in the condition. A naturally occurring decrease in the ratio of Δdi-mono4S to Δdi-mono6S has previously been described in aging articular cartilage in humans,9,15 and indeed a similar change was evident in the articular cartilage of mature sheep in this study.
The decreased ratio of Δdi-mono4S to Δdi-mono6S in newborn chondrodysplastic lambs indicates that although affected lambs cannot be distinguished phenotypically from normal or heterozygous lambs at birth, the underlying cartilage abnormality is present. Capillary electrophoresis techniques may therefore be useful in identifying the phenotype of newborn animals that are yet to demonstrate gross or microscopic lesions of chondrodysplasia. Although more samples would be required to confirm the true diagnostic power of the ratio of cartilage Δdi-mono4S to Δdi-mono6S, this study indicates that the information would at least be useful in future genetic studies as quantitative trait data associated with the chondrodysplastic phenotype.
The newborn affected lambs, which had no gross or microscopic lesions, and the mature affected ram, which had mild lesions of chondrodysplasia, all had ratios of Δdi-mono4S to Δdi-mono6S closer to the values for control animals than did the 1-month-old affected lamb, which demonstrated severe lesions of the disease. This suggested that the magnitude of the difference in the ratio of Δdi-mono4S to Δdi-mono6S may have been related to the severity of lesions. A study including a greater number of animals of different ages and representing the full spectrum of severity in this disease would be required to investigate this further. Such a study could be based on capillary electrophoresis of serial cartilage biopsies because of the small sample size (10 mg of cartilage) required for this technique.
Although the number of animals studied was relatively small, there are statistically significant, even striking, differences between affected sheep and control animals. Whatever the precise origins of the disease, it is clear that the manifest changes in sulfation at one particular position on the sugar ring produce large changes in the macroscopic mechanical properties of the resultant cartilage. Understanding in detail how such molecular change is mediated, presumably through modifications of biopolymeric assembly, would also be an interesting topic of further work.
Footnotes
Declaration of Conflicting Interests
The authors received no financial support for the research or authorship of this article.
Funding
The authors declare no potential conflicts of interests with respect to the authorship and publication of this article.