
Abstract
Activation of the transcription factor STAT5 is essential for the pathogenesis of acute myelogenous leukemia (AML) containing the FLT3 internal tandem duplication (ITD) mutation. FLT3 ITD is a constitutively active tyrosine kinase that drives the activation of STAT5, leading to the growth and survival of AML cells. Although there has been some success in identifying tyrosine kinase inhibitors that block the function of FLT3 ITD, there remains a continued need for effective treatment of this disease. We have identified the psychotropic drug pimozide as an effective inhibitor of STAT5 function. Pimozide inhibits the tyrosine phosphorylation of STAT5, leading to the death of AML cells through the induction of apoptosis. Pimozide shows a combinatorial effect with the tyrosine kinase inhibitors midostaurin (PKC412) and sunitinib in the inhibition of STAT5 tyrosine phosphorylation and the induction of apoptosis. Significantly, pimozide reduces the tumor burden in a mouse model of FLT3-driven AML. Therefore, identifying STAT5 inhibitors may provide a new avenue for the treatment of AML, and these may be effective alone or in combination with tyrosine kinase inhibitors.
Introduction
The initiation and maintenance of hematopoietic cancers are due to the inappropriate function of otherwise normal signaling pathways. In particular, members of the signal transducer and activator of transcription (STAT) family of transcription factors are often constitutively activated in cancer. 1 STATs are activated by receptor and nonreceptor tyrosine kinases by phosphorylation on a critical tyrosine residue, which leads to the activation (and sometimes repression) of transcription of their target genes. These genes code for proteins that are required for cellular survival, proliferation, and self-renewal. Therefore, STATs are critical for transducing signals from tyrosine kinases to the nucleus, where genes critical for the oncogenic phenotype of the cancer are regulated.
Many cancers are caused, at least in part, by the constitutive activation of tyrosine kinases, and the loss of this kinase activity can lead to the death of the cancer cell. 2 Although there have been significant strides in developing effective tyrosine kinase inhibitors, cancer cells frequently develop resistance to these drugs, most often by mutations that allow for the continued activation of STATs.3-6 This can be due to mutations in the targeted tyrosine kinase or the activation of an alternative kinase. Since STATs represent critical mediators of malignant cellular behavior, and they sit at the convergence point of many kinase pathways, the direct targeting of STATs may be an effective means of overcoming this resistance to tyrosine kinase inhibitors.
One of the most lethal hematological cancers is acute myelogenous leukemia (AML), a disease with a 5-year survival rate in the range of 20% to 40%. 7 The treatment for AML consists of conventional cytotoxic therapy that has been in use for decades and that has only limited effectiveness. There are several tyrosine kinase inhibitors in clinical trials geared toward the treatment of AML.8-10 Approximately 30% of AML tumors contain a mutant, constitutively activated form of the FLT3 tyrosine kinase, with the mutation most often being an internal tandem duplication (ITD) of the juxtamembrane domain.11,12 This mutant form of FLT3 results in the constitutive activation of STAT5, which likely contributes to the survival of these cells. Thus, a major focus is on the identification of specific FLT3 inhibitors. One FLT3 inhibitor currently in clinical trials is midostaurin, also known as PKC412. 13 Although this drug shows effectiveness in vitro and in mouse models of AML, it has thus far shown incomplete effectiveness in the clinic. Therefore, identifying new drugs to treat AML is of paramount importance.
Using a cell-based screen, 14 we have identified pimozide as a STAT5 inhibitor, which decreases STAT5 activated by BCR/ABL as well as a constitutively activated form of STAT5. 15 Pimozide is a neuroleptic drug used to treat Tourette syndrome but was not previously known to inhibit STAT function. In this study, we demonstrate that pimozide reduces the activation status of STAT5 in cell models of AML, which leads to the induction of apoptosis. Pimozide shows synergy when combined with PKC412 and the clinically relevant tyrosine kinase inhibitor sunitinib. Pimozide also shows efficacy in a mouse model of AML. These studies demonstrate that pimozide may be useful clinically for the treatment of AML either alone or in combination with other drugs.
Results
Pimozide reduces the activation of STAT5
Given the importance of STAT5 in the pathophysiology of AML, we examined the effect of pimozide on STAT5 activity in cells in which this protein is activated through mutation of FLT3. Ba/f3 cells reconstituted with the FLT3 ITD mutation (Ba/f3 FLT3 ITD) were treated with pimozide or with the tyrosine kinase inhibitor PKC412 (midostaurin), which directly inhibits FLT3. Pimozide caused a reduction of STAT5 phosphorylation at 3 hours (Fig. 1). Similar results were seen with the human AML cell line MV411, which endogenously expresses a FLT3 ITD mutation (Suppl. Fig. S1). PKC412 also inhibited STAT5 tyrosine phosphorylation (Fig. 1). Therefore, pimozide effectively reduces the activating phosphorylation of STAT5.
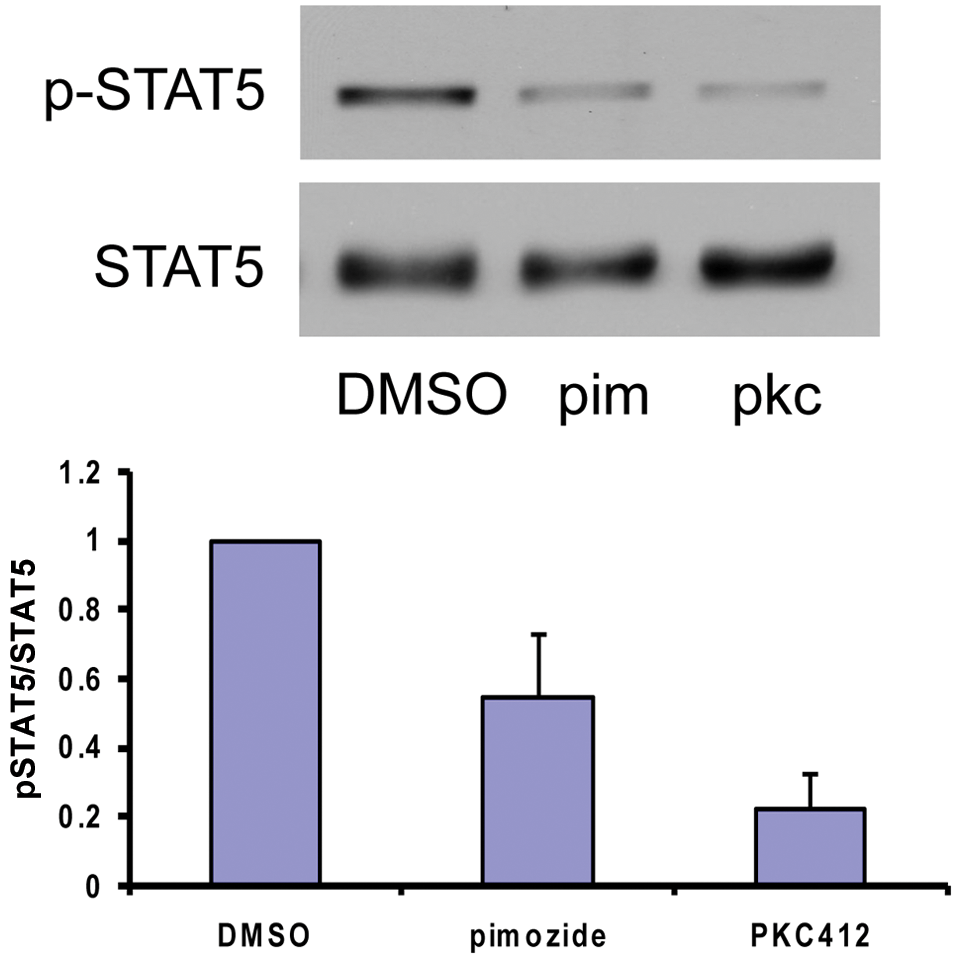
Pimozide reduces the tyrosine phosphorylation of STAT5. Ba/f3 FLT3 ITD cells were left untreated or treated for 3 hours with 5 µM pimozide or 5 nM PKC412, after which whole cell lysates were prepared and analyzed by immunoblotting for phosphorylated and total STAT5. The lower panel shows quantitation of 3 separate experiments.
Pimozide has little effect on FLT3 kinase activity
We considered the possibility that pimozide decreased STAT5 phosphorylation by inhibiting FLT3 tyrosine kinase activity. FLT3 activity is associated with autophosphorylation on numerous tyrosine residues, including tyrosine 591, which we measured by immunoblot analysis. In contrast to PKC412, which leads to near-complete inhibition of FLT3 phosphorylation, pimozide had only a minimal effect on FLT3 (Fig. 2). However, phosphorylation of FLT3 Tyr591 may not always be altered upon kinase inhibition. 16 Therefore, we performed in vitro kinase assays to further analyze kinase activity in the presence of pimozide. Whereas PKC412 significantly inhibited FLT3 activity in in vitro kinase assays, pimozide had no effect on FLT3 kinase activity (Table 1). These data indicate that, as opposed to PKC412, pimozide does not directly inhibit FLT3 function.
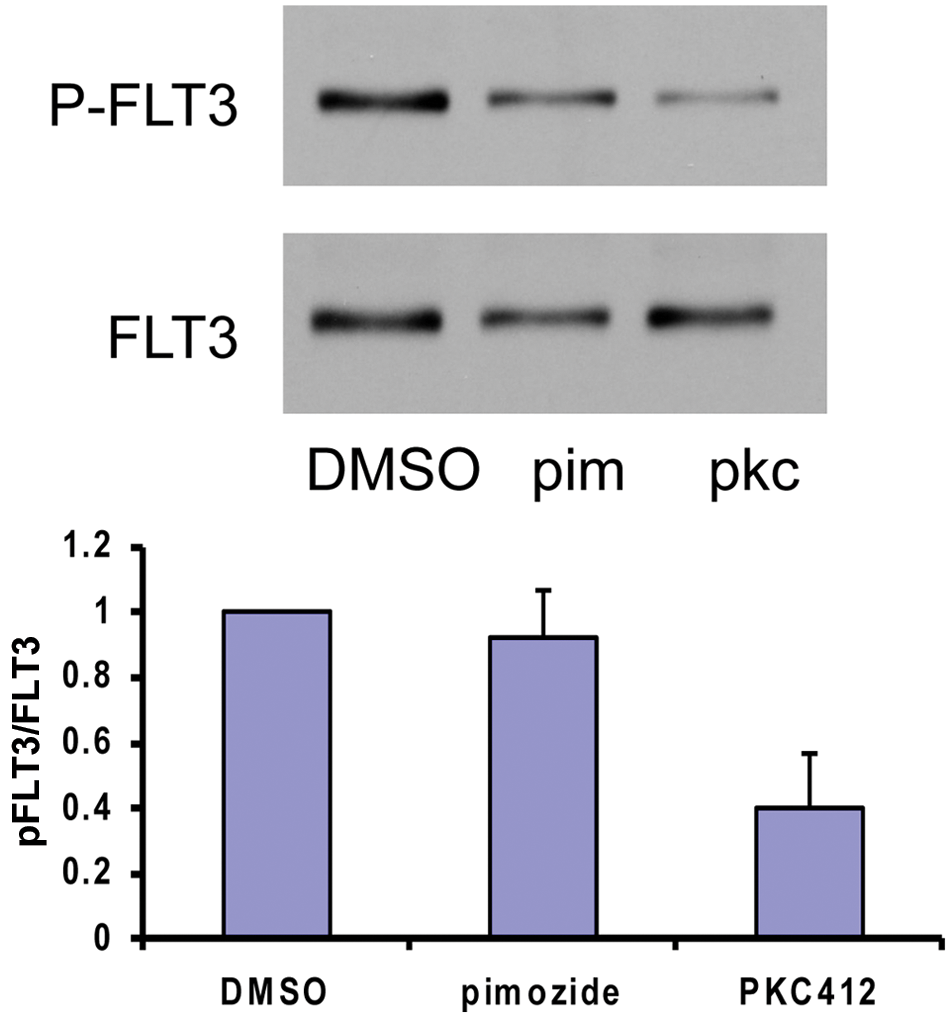
Pimozide has little effect on FLT3 phosphorylation. Ba/f3 FLT3 ITD cells were treated with 5 µM pimozide, or 5 nM PKC412, or were left untreated for 3 hours. Immunoblot analysis was performed using antibodies against phosphorylated and total FLT3, and the intensity of the bands was quantitated for 3 separate experiments.
Pimozide does not inhibit FLT3 tyrosine kinase activity

Note: The effect of pimozide (5 µM) was tested on the in vitro kinase activity of FLT3 and the FLT3 D835Y constitutively active mutant. PKC412 (5 nM) was used as a positive control. Values represent the percentage of inhibition of kinase activity, with values above 20 representing significant activity.
Pimozide reduces the viability of cells containing the FLT3 ITD mutation
Since pimozide reduces STAT5 phosphorylation, we reasoned that this drug would likely have an effect on cell viability. Therefore, we treated Ba/f3 FLT3 ITD cells or the human AML cell line MV411 (which endogenously contains the FLT3 ITD mutation) with increasing doses of pimozide for 48 hours. Pimozide treatment resulted in a distinct reduction in the number of viable cells, with an IC50 of between 3 and 5 µM for both cell types (Fig. 3A and 3B).
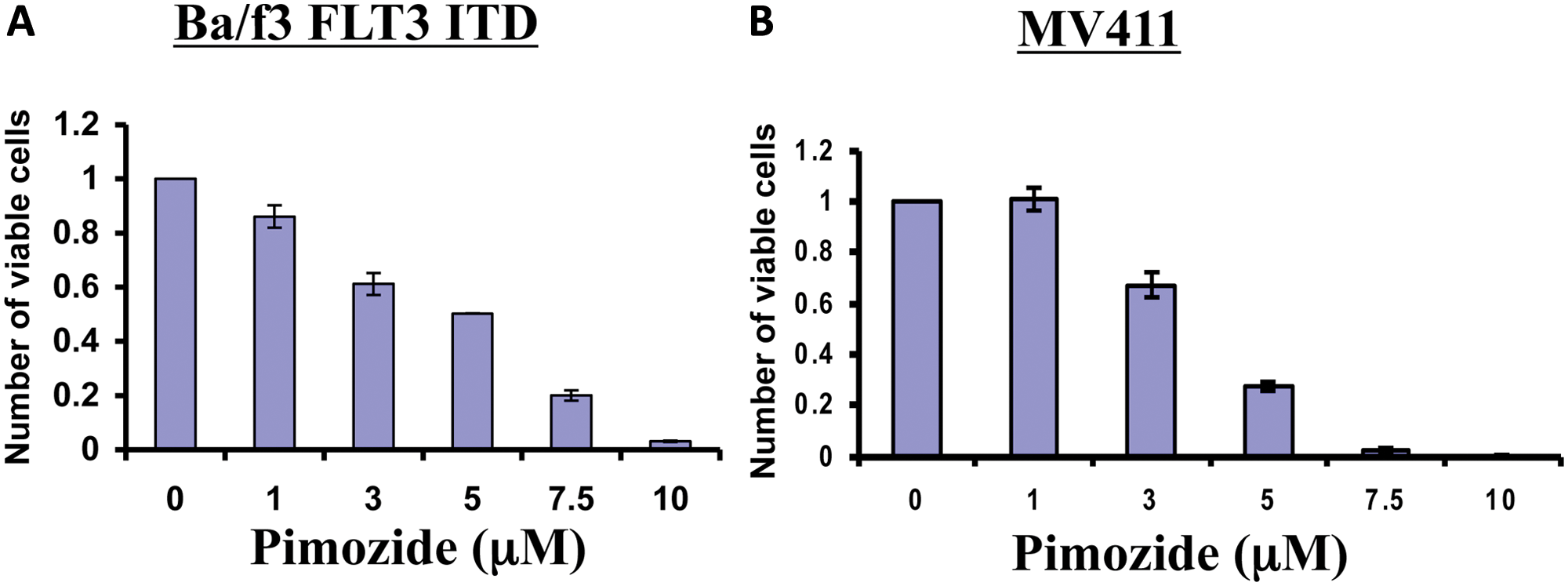
Pimozide reduces the viability of cells containing the FLT3 ITD mutation. Ba/f3 FLT3 ITD cells (
The combination of pimozide and PKC412 results in enhanced reduction of STAT5 phosphorylation
Since both pimozide and PKC412 inhibit STAT5 tyrosine phosphorylation through distinct mechanisms, we asked whether these drugs could be more effective in the inhibition of STAT5 phosphorylation when used in combination. Ba/f3 FLT3 ITD cells were treated with each drug at concentrations that give rise to approximately 50% reduction in viability. These doses result in a submaximal reduction in STAT5 phosphorylation. However, when used in combination, these doses of pimozide and PKC412 result in a near-complete inhibition of tyrosine phosphorylation of STAT5 (Fig. 4, left panel). Similar results were seen in MV411 cells (Fig. 4, right panel). Therefore, the effect of pimozide on tyrosine-phosphorylated STAT5 is significantly enhanced when combined with PKC412, suggesting that the combination of these drugs may result in a maximal effect.

The combination of pimozide and PKC412 results in enhanced reduction of STAT5 tyrosine phosphorylation. Ba/f3 FLT3 ITD cells (
Cells treated with pimozide and PKC412 show a synergistic loss of viability
Having shown that PKC412 enhances the effect of pimozide in inhibiting STAT5 phosphorylation, we analyzed the ability of this drug combination to affect cell viability. Ba/f3 FLT3 ITD cells were treated with a range of dose combinations of pimozide and PKC412, and the number of viable cells was evaluated. Nearly all combinations of these drugs show a synergistic loss of viability as measured by combination index quantitation (combinatorial index [CI] values of 0.37-1.0) and isobologram analysis (Fig. 5A). Similar results were seen with MV411 cells (Suppl. Fig. S2).
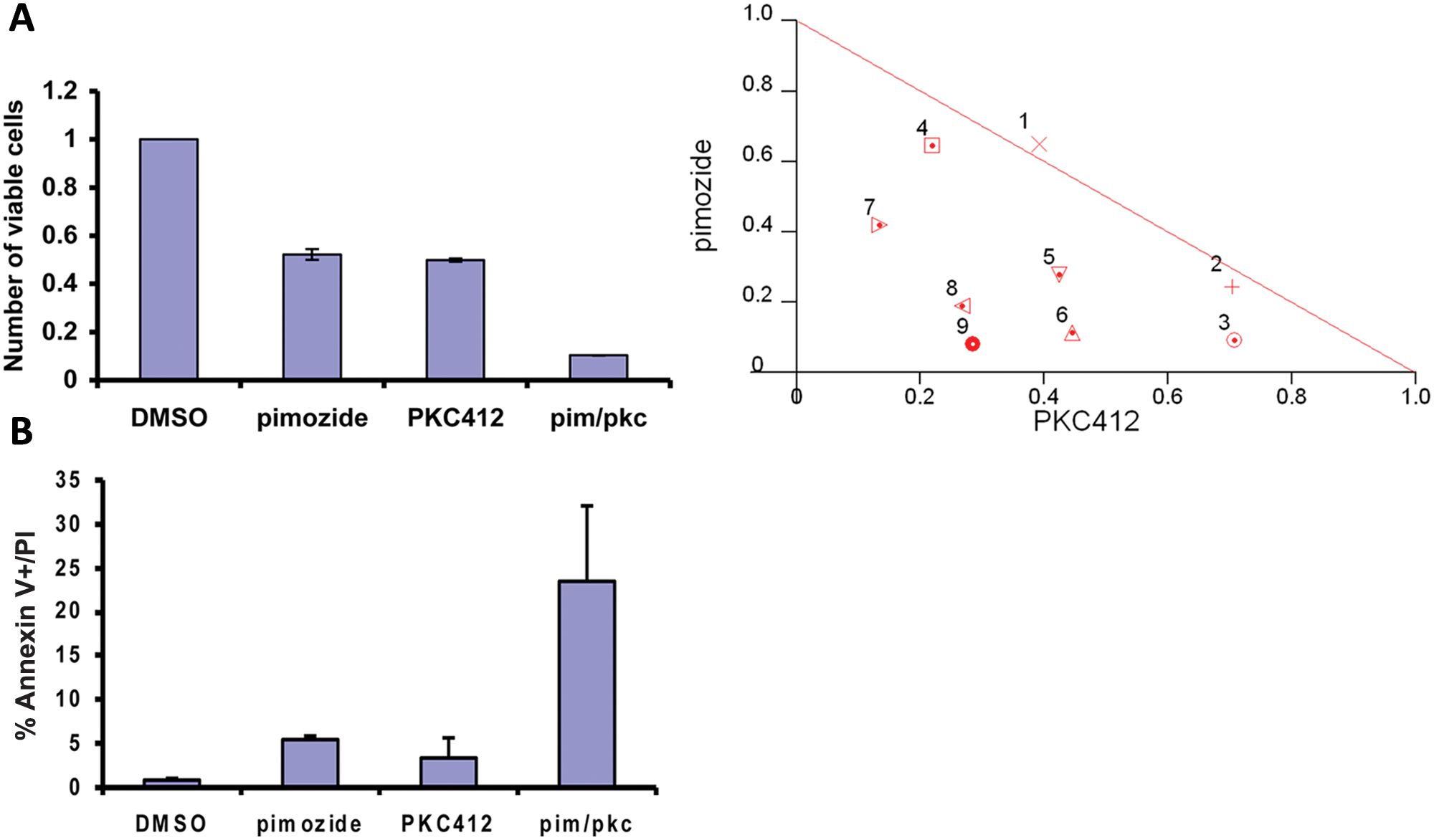
Pimozide and PKC412 synergize to reduce cell viability and induce apoptosis. (
To determine the mechanism for this combinatorial effect, Ba/f3 FLT3 ITD cells were treated with pimozide and PKC412 alone and in combination, stained with propidium iodide (PI) and annexin V–FITC, and analyzed by flow cytometry. The combination of pimozide and PKC412 showed an approximate 6-fold increase in annexin V–positive/PI-negative cells (Fig. 5B), indicating that this combination results in significantly enhanced apoptosis.
The effect of pimozide on STAT5 phosphorylation is enhanced by the multikinase inhibitor sunitinib
Since the FLT3 inhibitor PKC412 enhanced the effect of pimozide, we asked whether other tyrosine kinase inhibitors already in clinical use could exert a similar effect. To examine this, we utilized the multi–tyrosine kinase inhibitor sunitinib, which has been found to inhibit FLT3 activity. 17 Using doses of both pimozide and sunitinib that give rise to a 50% reduction in viability of Ba/f3 FLT3 ITD cells, we analyzed STAT5 phosphorylation by immunoblot. Whereas both pimozide and sunitinib alone showed partial reduction of STAT5 phosphorylation, the combination of these drugs maximally reduced STAT5 phosphorylation (Fig. 6A). Thus, the combination of pimozide with the multikinase inhibitor sunitinib shows an enhanced effect on STAT5 phosphorylation.
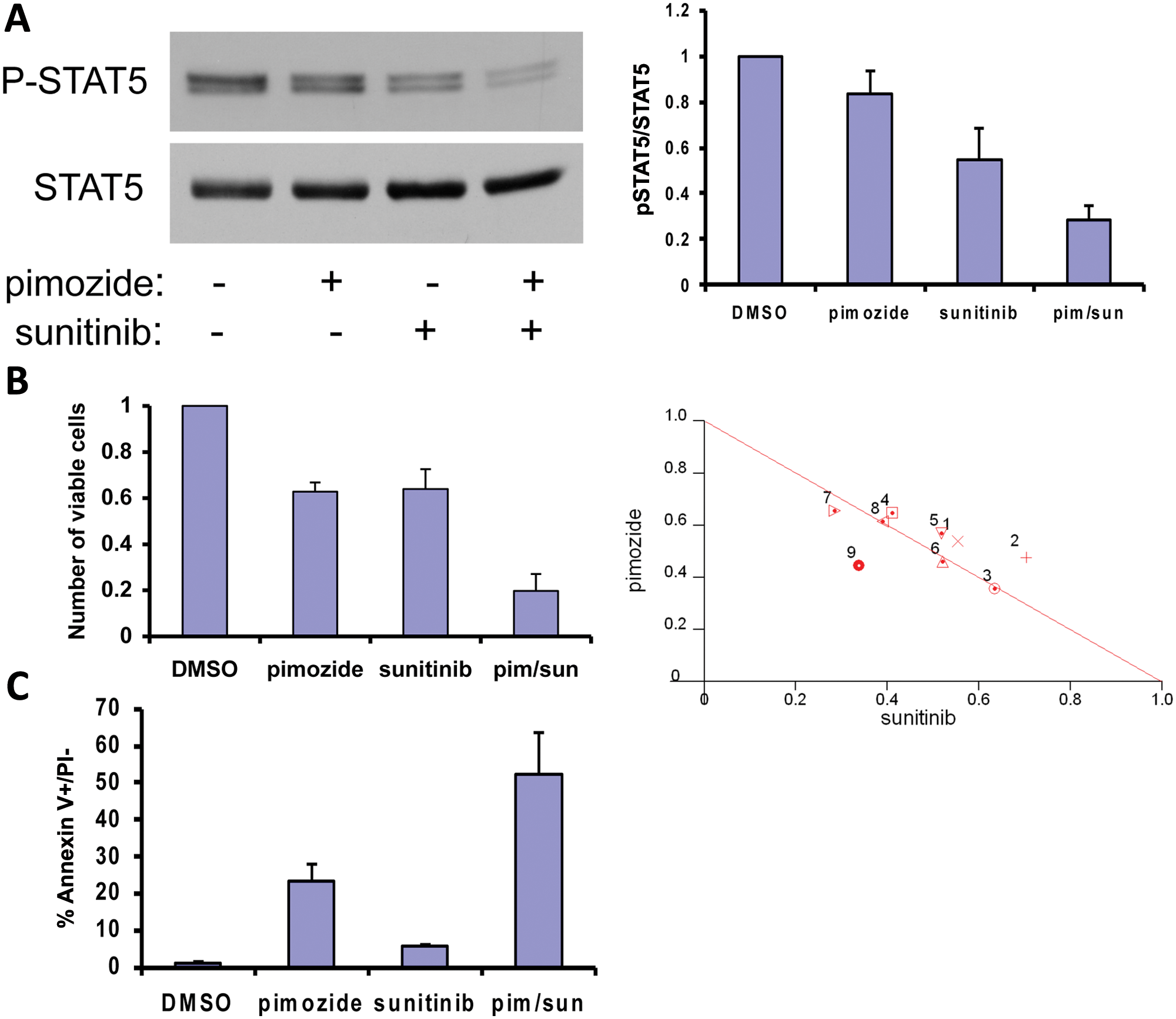
Pimozide and sunitinib synergize in a cellular model of AML. (
Cells treated with pimozide and sunitinib show enhanced loss of viability
Given the notable combinatorial effect of pimozide and sunitinib on STAT5 tyrosine phosphorylation, we wanted to determine if the combination of these drugs affected cellular viability. Ba/f3 FLT3 ITD cells were treated with 9 different dose combinations of pimozide and sunitinib, and the number of viable cells was determined (Fig. 6B). Most dose combinations show a moderately synergistic to additive effect when analyzed using the combination index calculation (CI values of 0.78-1.1). In addition, 9 dose combinations of pimozide and sunitinib in MV411 cells are also in the moderately synergistic to additive range, with CI values between 0.67 and 1.1 (Suppl. Fig. S3).
To determine if the enhanced loss of viability can be seen at the level of apoptosis, we treated cells with pimozide and sunitinib and measured apoptosis by quantitating annexin V/PI–positive cells. The combination of pimozide and sunitinib caused increased apoptosis in Ba/f3 FLT3 ITD cells (Fig. 6C). These data demonstrate that the synergistic to additive effects seen with this combination in viability assays correlate with increased apoptosis.
Pimozide reduces the tumor burden in a mouse model of AML
Having shown that pimozide reduces cell viability through an induction of apoptosis in cellular models, we next examined the effect of pimozide in a mouse model of AML. FLT3 ITD–transformed Ba/f3 cells (engineered to express firefly luciferase) were inoculated by intravenous injection into immunodeficient nude mice. As previously described, animals developed disseminated leukemic disease, which could be visualized and quantified by bioluminescence imaging. 18 Mice were treated by oral administration with 25 mg/kg pimozide or 25 mg/kg PKC412 as a positive control. Doses of PKC412 as high as 100 mg/kg have been used in murine studies, although we wished to use the minimal dose predicted to fully inhibit FLT3. Treatment with pimozide reduced the tumor burden assessed by bioluminescence imaging, comparable to treatment with PKC412 (Fig. 7A). Control mice treated with the vehicle became moribund, with most animals requiring sacrifice within 19 days of starting drug treatment (Fig. 7B), thereby confounding group-wise comparisons using bioluminescence imaging. Therefore, we continued to treat animals and determined the effects of treatment on animal survival. Treatment with pimozide prolonged the survival of mice to an extent comparable to treatment with PKC412 (Fig. 7B). Both treatments were well tolerated with no significant effects on body weight (Fig. 7C). The combination of pimozide and PKC412 at these doses was not well tolerated by the mice, and thus, efficacy studies of the combination could not be performed. Nonetheless, these data indicate that pimozide has antileukemia efficacy in vivo without overt toxicity.
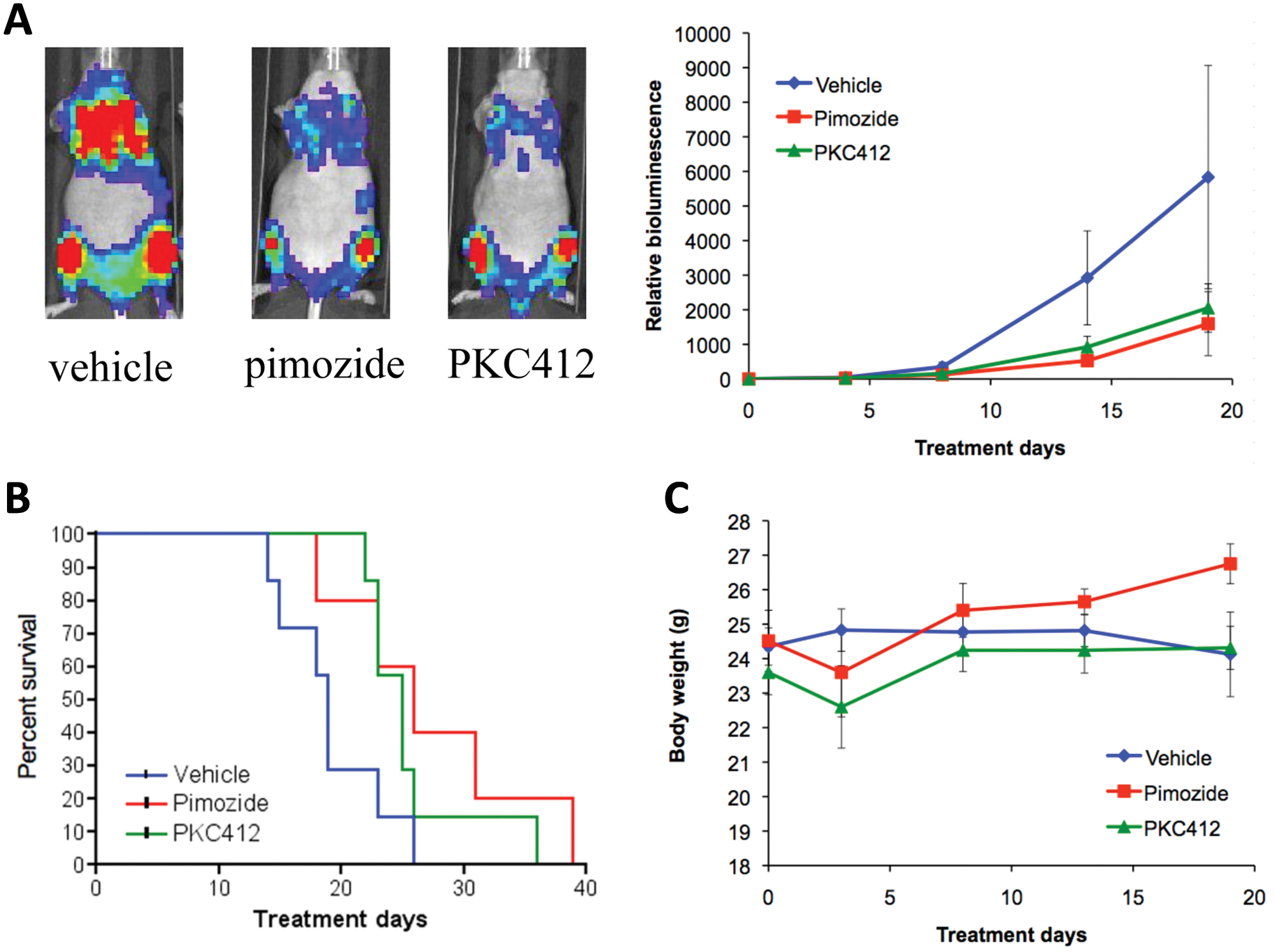
Pimozide reduces the tumor burden in a mouse model of AML. Pimozide or PKC412 was administered to mice previously injected with Ba/f3 FLT3 ITD luc+ cells. (
Discussion
Since tyrosine kinases are essential for the survival of many tumors, a great deal of research has gone into the development of tyrosine kinase inhibitors. There are a number of such inhibitors currently in clinical trials or clinical use, and these have shown significant efficacy. These include imatinib, nilotinib, sunitinib, and dasatinib, among others.19-22 Although tyrosine kinase inhibitors have shown effectiveness, they rarely eradicate a tumor, and the emergence of resistance remains a significant concern.
The many years of clinical use of imatinib provides an excellent resource to study the emergence of resistance to tyrosine kinase inhibitors in a clinical setting. 23 Although less is known about resistance to FLT3 inhibitors, resistance to PKC412 has been documented in cell culture models, and these studies have demonstrated a number of different mechanisms of resistance. 24 In addition, one patient enrolled in a clinical trial showed an initial response to PKC412 but later developed resistance. 25 Analysis of samples from this patient demonstrated a point mutation in the kinase domain of FLT3, suggesting that mutations in FLT3 itself can override the effects of PKC412 in a clinical setting, as is seen with imatinib-treated chronic myelogenous leukemia (CML) patients. These findings suggest that the identification of additional complementary and synergistic targets, especially downstream mediators such as STAT5, is of great importance for improved patient outcomes.
A major downstream target of many tyrosine kinases, particularly in hematopoietic cancers, is the transcription factor STAT5, and there are several reasons why targeting STAT5 might be particularly important in cancer therapy.26,27 First, STAT5 controls the expression of genes that directly regulate malignant cellular behavior, and it is necessary for tumor maintenance. Second, STAT5 activation is driven by many tumorigenic tyrosine kinases, and thus, specific STAT5 inhibitors may be useful in a range of cancers driven by different upstream mutations. This includes not only Flt3-driven AML but also other forms of AML, acute lymphoblastic leukemia (ALL), CML, and lymphomas, among others. Third, resistance to tyrosine kinase inhibitors often arises from secondary mutations that lead to the continued activation of STAT5, and thus, directly targeting STAT5 might circumvent a range of resistance mechanisms. Fourth, as shown here, the combination of tyrosine kinase inhibitors and STAT5 inhibitors may be a particularly effective means of achieving drug synergy. Finally, genetic and pharmacological studies have indicated that inhibition of STAT5 in normal cells appears to have few deleterious effects,15,28 and thus, it is a target with a potentially high therapeutic index.
Not only is targeting STAT5 a useful therapeutic approach in hematological cancers, but the combination of STAT5 inhibition and antagonism of an upstream tyrosine kinase may also be particularly effective. Our in vitro data demonstrated a clear synergism when pimozide and PKC412 were combined (Figs. 4 and 5). In our cellular assays, we have also shown synergism between pimozide and sunitinib (Fig. 6), and we have previously shown synergism between pimozide and either imatinib or nilotinib in CML. 15 In vivo toxicity precluded our ability to assess the efficacy of pimozide and PKC412 in vivo. However, the combination of pimozide and kinase inhibitors may be a promising therapeutic approach.
Pimozide is approved by the United States Food and Drug Administration for the treatment of Tourette syndrome, and it is used elsewhere for the treatment of schizophrenia. Although it can cause cardiac toxicity, it has not been shown to have adverse effects on normal hematopoietic cells. In fact, while pimozide can decrease colony formation of bone marrow progenitor cells derived from patients with CML, it has almost no effect on hematopoietic progenitors derived from healthy donors. 15 Given the biochemical and biological synergy seen between pimozide and kinase inhibitors, and the possibility that even brief exposures to pimozide may have therapeutic effects, 29 the use of pimozide or other STAT5 inhibitors may be a promising approach.
Pimozide was identified as a STAT5 inhibitor through a cell-based screen in which drugs already known to be safe in humans were analyzed for their ability to inhibit STAT-dependent gene expression.30,31 The advantage of this approach is that it can greatly shorten the amount of time needed to initiate a clinical trial. However, a challenge inherent with this strategy is that the direct cellular target of an active drug may be difficult to elucidate. Pimozide decreases STAT5 phosphorylation, but it does not have activity as a tyrosine kinase inhibitor. We are currently engaged in a genetic screen to elucidate the mechanism of action of pimozide and other STAT inhibitors. This approach should also identify steps in the STAT signaling pathway that are amenable to pharmacological inhibition or whose perturbation may synergize with current STAT inhibitors. Thus, in addition to its potential use as an anticancer agent, pimozide, and other STAT modulators, may help identify novel cellular targets, further reflecting the utility of this type of chemical biology approach.
In this study, we show that pimozide is a STAT5 inhibitor in AML cells containing constitutively activated FLT3 and that it is effective in cell culture and in vivo models of AML. Therefore, inhibition of STAT5 might be a useful strategy for the treatment of AML alone or in combination with other therapeutic treatments.
Materials and Methods
Cell lines
MOLM14 cells were a kind gift from Daniel G. Tenen (Beth Israel Deaconess Medical Center, Boston MA), MV411 cells were from the American Type Culture Collection (Manassas, VA), and Ba/f3 FLT3 ITD cells were obtained from D. Gary Gilliland (Brigham and Women’s Hospital, Boston, MA). 13 All cells were cultured in RPMI media supplemented with 10% fetal calf serum.
Compounds
Pimozide was obtained from Sigma (St. Louis, MO); PKC412 was from Novartis Pharma AG (Basel, Switzerland). For in vitro experiments, drugs were dissolved in DMSO and were diluted to a final concentration of 0.1% DMSO in all experiments.
Antibodies
Antibodies recognizing STAT5 (sc-835) and FLT3 (sc-479) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Antibodies to phosphotyrosine (9411 and 9416), phospho-STAT5 (9359), and phospho-FLT3 (3461) were obtained from Cell Signaling Technology (Danvers, MA).
Immunoblotting and immunoprecipitation
Immunoblotting and immunoprecipitation were performed as previously described. 14 Band intensities were quantitated using ImageJ software (National Institutes of Health, Bethesda, MD).
In vitro kinase assay
Kinase inhibitory activity of pimozide was analyzed using the SelectScreen Kinase Profiling service (Invitrogen, Carlsbad, CA).
Viability assays
The number of viable cells was measured using the CellTiter-Glo ATP-dependent luminescent assay (Promega, Madison, WI).
Statistical analysis
Synergy in cell viability assays was determined by plotting isobolograms and quantitating the CI using CalcuSyn software (Conservion, Ferguson, MO).
Apoptosis assay
Quantification of apoptotic cells was performed using the annexin V–FLUOS Staining Kit (Roche Applied Science, Penzberg, Germany). Cell suspensions were washed twice with PBS. Staining and counterstaining with annexin V–FITC and PI, respectively, were performed following the manufacturer’s instructions. Analysis was performed using the BD FACSCano II Flow Cytometer (BD Biosciences, San Jose, CA) using negative and single color controls to adjust for compensation. Apoptotic cells were quantified and differentiated from viable and necrotic cells using FACSDiva 6.1 software (BD Biosciences).
Mouse studies
Ba/f3 FLT3 ITD luc+ cells were resuspended in PBS, and a total of 106 cells were injected intravenously via the lateral tail vein into 8-week-old female NCr nude mice (Charles River Laboratories, Wilmington, MA). Baseline bioluminescence imaging, as previously described, 32 was performed 4 days after tumor cell inoculation, and mice with established disease were divided into cohorts (n = 7 per group) with a matched tumor burden. Mice were treated with either 25 mg/kg pimozide by oral gavage (PO), 25 mg/kg PKC412 (Novartis) PO, or a vehicle at a volume of 10 mL/kg PO. Treatments were administered in 10-day cycles consisting of continuous drug treatments for 7 days, followed by 3 days off. Mice with evidence of distress were humanely euthanized, and statistical significance of survival prolongation was determined by the log-rank test. All studies involving mice were performed according to Dana-Farber Cancer Institute’s Animal Care and Use Committee–approved protocols.
Footnotes
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: J.D.G. and A.L.K. have a financial interest in Novartis.
Funding
The author(s) received the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Multiple Myeloma Research Foundation (Norwalk, CT), Kittredge Foundation (Dana-Farber Cancer Institute), Brent Leahey Fund (Dana-Farber Cancer Institute), Gabrielle’s Angel Foundation (New York, NY), and the Claudia Adams Barr Program in Innovative Basic Cancer Research (Dana-Farber Cancer Institute).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.