
Abstract
In tropical environments, the microclimatic conditions that modulate the edge influence in naturally fragmented forests have scantly been studied. Also, it is virtually unknown how microclimatic variations in the matrix-patch transition are associated with traditional harvesting practices occurring mainly at the edge of forest patches. Here, we documented three microclimatic parameters (temperature, relative humidity, and photosynthetically active radiation) in the matrix-patch transition of natural forest patches or petenes with and without palm (Sabal japa) extraction in a coastal wetland of the Yucatan Peninsula. The microclimatic parameters were recorded in six petenes with and without palm extraction (three sites per type), along three radial transects from outside (matrix) to the forest patch interior, and in diurnal periods when the environmental conditions of the wetland were most extreme but constant during the day. In contrast to relative humidity, which notably decreased during the dry season, temperature and photosynthetically active radiation showed similar variation along the matrix-patch transition in petenes with and without palm extraction. Only photosynthetically active radiation decreased toward the interior of the peten, with the highest values starting at the matrix in the rainy and dry seasons. The scant variation for temperature and relative humidity, as well as changes in PAR from the matrix to the interior of petenes, suggests an edge influence modulated by the patch’s forest structure and the presence of mangroves, even in those petenes with palm harvesting. The abiotic parameters studied in petenes suggest an edge influence mainly associated with the vegetation structure and presence of mangroves on the patch’s periphery that appears to promote the persistence of petenes in this naturally fragmented Mesoamerican ecosystem.
Introduction
The forest edge constitutes a central tenet of modern ecology (Cerboncini et al., 2016; Rinas et al., 2024) and is mainly studied in anthropogenic forest fragments. Environmental changes at the edge influence the ecological dynamic of the fragments and are primarily associated with their biogeographical characteristics and the surrounding conditions defining the environmental matrix (Benitez-Malvido et al., 2014; Laurance et al., 2007; Ries et al., 2004). In the ecological transition zone represented by the edge, differential matrix-fragment conditions produce biophysical changes that positively or negatively influence the ecological distribution and interactions of the local biota (Porensky & Young, 2013). Such biophysical changes in the edge are primarily associated with microclimatic conditions gradually changing from the border towards the forest interior (e.g.; air and soil temperature - Williams-Linera, 1990; air humidity - Kapos, 1989; solar radiation - Brothers & Spingarn, 1992), promoting environmental conditions linked to the edge influence (Laurance et al., 2002; Murcia, 1995; Watts & Kobziar, 2015).
To date, there is relatively little information about the microclimatic conditions that modulate the edge influence in natural forest patches (Franklin et al., 2021; Moen & Jonsson, 2003), where their “inherent” edges (representing long-term landscape features) could be experiencing lower edge influence than anthropogenic edges (Harper et al., 2015). In naturally fragmented forests (not resulting from anthropogenic deforestation), the interfaces between non-forested environmental matrix and forest patches are expected to be more gradual but more complex, with variable transition zones as compared with anthropogenic edges (Franklin et al., 2021; Harper et al., 2015; Moen & Jonsson, 2003). Also, it is virtually unknown how traditional harvesting practices occurring in the matrix-patch transition are associated with changes in microclimatic conditions mainly at the patch edge (Olupot et al., 2009).
In the coastal wetlands of northwest Yucatan Peninsula, forest vegetation is mainly composed of naturally formed forest patches or petenes (singular peten), which are immersed in a seasonably floodable (by rainfall and to a lesser extent, by sea water) environmental matrix (Montiel et al., 2006; Munguía-Rosas et al., 2014; Rico-Gray, 1982). In contrast with forest fragments, petenes are not the result of human activity, instead deriving from long-term geo-ecological processes that have formed an endemic naturally fragmented ecosystem with high ecological stability and persistence in southeast Mexico (Mas & Correa-Sandoval, 2000; Montiel et al., 2006; Munguía-Rosas & Montiel, 2014).
In terms of their sociocultural importance, petenes provide valuable natural resources (flora and fauna) for the rural Mayan population (García et al., 2015; León & Montiel, 2008; Méndez-Cabrera & Montiel, 2007). This is the case of guano palm (Sabal japa), a non-woody resource whose harvesting is part of the multiple natural resource use strategy characteristic of Maya culture (Barrera-Bassols & Toledo, 2005; Oliva et al., 2014). Traditionally, palm harvesting in petenes mainly occurs at the forest edge, which offers better conditions of access and mobility conditions for people during the extractive activity.
Here, we present a first assessment of microclimatic variation across the forest edge in petenes and its relationship with a traditional harvesting practice occurring mainly at the forest edge in a coastal wetland of the Yucatan Peninsula. Our objectives were to (a) evaluate three microclimatic parameters (temperature, relative humidity, and photosynthetically active radiation) in petenes with and without palm extraction (S. japa), (b) evaluate potential changes in the microclimatic parameters between the rainy and dry seasons for petenes with palm extraction, and (c) assess changes in the forest structure (number of leaves per vertical stratum and foliar interference) as a proxy for biotic factors, in the matrix-patch transition.
We expected 1) microclimatic parameters (temperature, relative humidity, PAR) to vary significantly from the exterior (matrix) towards the interior of petenes, indicating an edge influence, 2) palm extraction to increase light penetration (higher PAR) and temperature, and reduces relative humidity at the edge of petenes, 3) seasonal changes (rainy vs dry) to significantly influence relative humidity and temperature, in petenes with palm extraction, and 4) foliar interference to be negatively correlated with PAR along the edge gradient. Finally, we discuss how even in petenes with palm extraction, the forest structure and the presence of mangroves on the patch’s periphery appear to promote the ecological stability and persistence of petenes in this important naturally fragmented Mesoamerican ecosystem.
Materials and Methods
Study Area
The study was carried out in the Los Petenes Biosphere Reserve (LPBR; 20°51′30″-19°49′ 00″ N and 90°45′15″-90°20′00″ W; area = 282,858 ha, 64% marine and 36% terrestrial), located in the west coast of the Yucatan Peninsula (Figure 1). The climate of the region is warm and sub-humid, with a mean monthly temperature of 26°C and mean annual precipitation of 819 mm (Yáñez-Arancibia, 1996). Regionally, there is a dry season (December-May; mean monthly precipitation = 13.2 mm) and a rainy season (June-November; mean monthly precipitation = 149 mm) (Montiel et al., 2006). Location of Los Petenes Biosphere Reserve (LPBR) in the Northwest of the Yucatan Peninsula, Mexico. The Six petenes Selected (Stars) for the Study Are Shown Near the Northern Limit (Red Line) of the Reserve in the Projected Area. Imagens Taken From Google Earth
Due to its high biological diversity and biogeographical characteristics, since 1999 the endemic petenes ecosystem has been under federal protection and constitutes a priority conservation area in Mexico (Comision Nacional de Areas Naturales Protegidas; CONANP, 2006), having been declared a wetland of international importance (Ramsar site) in 2004. The LPBR presents terrestrial vegetation composed of at least 678 species of higher plants (103 families and 404 genera), including 24 endemic species to the Yucatan Peninsula, one endangered (Echinodorus nymphaeifolius) and five specially protected (Cupressus lusitanica, Roystonea regia, Avicenia germinans, Laguncularia racemosa, Rizhopora mangle). The coastal wetland is composed of different ecosystems such as mangroves (R. mangle, A. germinans, L. racemosa, Conocarpus erectus), grassland (sedges - Eleocharis cellulose, Cladium jamaicense; cattails - Typha dominguensis), and petenes (CONANP, 2006).
Petenes constitute the most notable characteristic of the LPBR as natural forest patches of semideciduous tropical rainforest and mangroves (with trees such as R. mangle, Manilkara zapota, Metopium brownei, Bursera simaruba, L. racemosa, A. germinans dominating the canopy and Bravaisia tubiflora and S. japa dominating the understory) (Figure 2). These forest patches (>1000 units) are semicircular in shape and with varying mean areas and isolation distances (ca. 11 ha and 200 m, respectively; Montiel et al., 2006), embedded in a seasonably floodable matrix composed of structurally contrasting vegetation, dominated by grassland (typically with aquatic vegetation dominated by T. dominguensis, the sawgrass C. jamaicense) and dispersed dwarf mangroves (Munguía-Rosas et al., 2014; Rico-Gray, 1982). Aerial Views of Petenes in a Coastal Wetland of the Northwest Yucatan Peninsula
The LPBR, unlike other reserves in the region, has no human settlements in its terrestrial area. Nevertheless, as part of its zone of influence (ca. 1125 km2 along the reserve’s eastern border) there are at least 20 rural communities that have historically reproduced the multiple natural resource use strategy characteristic of the Maya culture (Barrera-Bassols & Toledo, 2005; Oliva et al., 2014). One of the activities shaping the livelihoods of inhabitants living near the LPBR is commonly associated with obtaining guano palm (S. japa) from petenes (and to a lesser extent from communal areas surrounding rural communities) (Figure 3), mainly for handcrafts and the construction of vernacular Maya housing (Caballero et al., 2001; Sánchez-Suárez et al., 2021). Palm (Sabal japa) Fronds Traditionally Tied Into a Roll (50 Fronds per Roll; See Five Rolls in the Image) After Local Harvesting in petenes (A). Mayan-Peasant Transporting the Harvested Palm Fronds to the Community (B)
Study Sites
We selected a total of six petenes for this study based on their accessibility, shape, and size: three petenes with and without guano palm extraction (S. japa), respectively. All the sites were clearly differentiable and accessible on the ground, having a similar size (<10 ha) and shape (approximately circular), as well as bordering grassland on at least 50% of their periphery.
Sampling Design
The records obtained in the studied petenes were taken during the rainy season (October-November), and during the dry season (March) when palm extraction traditionally occurs. The first period allowed for an evaluation of the microclimatic parameters in the studied petenes with and without palm extraction, during the same season. The second period allowed for an inter-seasonal prospection (rainy vs dry) of the potential seasonal change in the microclimatic parameters along the matrix-patch transition only for petenes with palm extraction.
In each selected peten, the microclimatic parameters were recorded every three meters along three radial transects of 50 m in length from the exterior (including 10 m of the matrix) toward the interior (40 m) of the forest patch. GPS coordinates allowed that each radial transect to be established and oriented from the matrix towards the center of the peten. From the first radial transect of the matrix-patch transition, the other two transects were established at similar perimetral distances (>20 m between transects) crossing the edge of each peten (Figure 4). To reduce possible disturbances at the recording points, the measurements taken with a HOBO (data logger U30) along the transect were made systematically at the same height (1.3 m) by the observer. For the inter-seasonal prospection (rainy vs dry), in addition to abiotic parameters, only the foliar interference percentage was measured along the transects originally established in the studied petenes. Matrix-peten Transition Area where the Three Abiotic Parameters (Temperature, Relative Humidity and PAR) Were Evaluated at the Study Sites. For Illustrative Purposes, the Aerial View Projected Shows the Approximate Location on the Edge of the Three Radial Transects (Yellow Bands), From the Surrounding Matrix Towards the Interior of the Forest Patch, Established to Record the Abiotic Parameters by Transect (3 per Site) in Three petenes With and Without Palm Extraction Respectively (See Sampling Design in Methods for Details)
The microclimatic parameters were recorded continuously (ca. taking one hour per transect) between 12:00 p.m. and 3:00 p.m. when the environmental conditions of the wetland (e.g., temperature and radiation) were most extreme but constant during the day. This allowed for the exclusion of drastic variations in the matrix-patch transition associated with the time of day, which is highly desirable for edge studies (Ries et al., 2004).
Microclimatic Parameters and Vegetation Structure
For this study, three microclimatic parameters were selected: temperature (0 C), relative humidity (%), and photosynthetically active radiation (PAR; µmol/m-2 s-1). These parameters were recorded in border areas between the peten and the matrix with structurally more contrasting vegetation. Differentiation of the matrix-patch transition at the study sites was performed based on biogeographical data of the cluster of petenes (using Google Earth) and supplemented by field trips carried out at the start of the study. The recording of microclimatic parameters was calibrated based on a pilot performed on a radial transect with the greatest matrix-patch contrast.
Additionally, two variables, number of leaves per vertical stratum (total leaves touching a post marked at heights of interest) and foliar interference percentage (% = area of leaf coverage/total area for a circular photograph), were recorded jointly with the microclimatic parameters to have a proxy for biotic factors in the matrix-patch transition. The number of leaves per vertical stratum was recorded using a 5-m post placed vertically on the ground at each recording point (Schemske & Brokaw, 1981), with marks at 13 height intervals (12 equally divided between 0 and 4.5 m and 1 for measurements taken > 4.5 m). For each interval, the number of leaves in contact with the post was quantified, taking records every 20 cm between a height of 1 and 2 meters, where the measurements of the three abiotic parameters were made. The foliar interference percentage was estimated based on photographs taken at each recording point with a digital camera oriented at the canopy. To obtain an image of a circular area of a standard size for each recording point, a PVC cylinder was placed over the lens of the camera. Each photograph was subsequently converted to a binary image (using the ImageJ software package version 2017) to estimate the foliar interference percentage based on the percentage of black pixels in the image, like approaches such as those reported by Jennings et al. (1999).
Data Analysis
The variation of the three microclimatic parameters and foliar interference between types of peten (with and without palm extraction) was compared under two analytical approaches: 1) a graphical approach of the distribution curve of mean values (±SE) of each parameter recorded along the matrix-patch transition between type of peten (3 sites per type), and 2) since the data for all parameters was not found to be normally distributed (Shapiro-Wilk test: W = 0.88, 0.78, 0.6, 0.79, P < 0.05; temperature, relative humidity, PAR and foliar interference, respectively), for each case we applied an Aligned Rank Transform Analysis of Variance (ART-ANOVA), a non-parametric technique to evaluate multifactorial contrast models detecting principal effects and interactions in non-parametric data (Elkin et al., 2021; Wobbrock et al., 2011).
For this ART-ANOVA, all data sets (N = 144 units each) per parameter were averaged per site (block) acknowledging the pseudo-replication associated with the transects (n = 3 per site). Thus, for each analysis, we defined type of peten (factor A with two levels, with vs without palm extraction) and section of the matrix-patch transition [factor B, with four levels: matrix (-10 to 0 m), exterior edge (0.1-12 m), mid edge (12.1-24 m), interior edge (24.1-39 m)]. For this study, an interaction effect in the model was used for testing statistical significance (alpha = 0.05) for type of peten x matrix-patch transition for all non-parametric microclimatic parameters and foliar interference data.
In our study, defining the four sections of the matrix-patch transition allowed us to better approach the variation of the microclimatic parameters from the exterior (matrix) toward the interior of the peten, taking account of changes in their values with respect to the values recorded for the matrix (the initial 10 m section of each transect). The distances of these transect sections were defined based on a) the most conspicuous changes in PAR and forest structure noted by us during the pilot study performed on a radial transect with the greatest matrix-peten contrast, and b) published data on border-and-ecotone analysis using microclimatic parameters (i.e.; vapor pressure deficit; VDPampl.) from ecotones between forest patches and savanna (revealing that the ecotone initiates ca. 12-30 m towards the forest interior; Hennenberg et al., 2008). Considering the two previous criteria, we defined the four sections of the matrix-patch transition for our prospective patch edge analysis.
The variation in the microclimatic parameters and foliar interference between the rainy and dry seasons was only compared for petenes with palm extraction following the two previous approaches, except in this case a repeated-measures ART-ANOVA was applied: section of the matrix-patch transition (with four levels) and season (rainy vs dry), with two repeated measures per site.
In all cases, ART-ANOVA contrast models were used to determine differences between levels of the factor that included the sections of the matrix-patch transition. For the records of foliar interception of each transect, the number of leaves intercepted by height interval was compared between types of peten using the Mann-Whitney U test (Zar, 1996). All statistical analyses were performed using the ARTool package in R (Kay et al., 2025).
Results
Microclimatic Parameters Across the Forest Edge in the Rainy Season
Records (Mean ± SE) of Abiotic Parameters and Foliar Interference Along the Matrix-Forest Patch Transition in petenes With and Without Palm Extraction (PE) During the Rainy and Dry Season (See Data Analysis in Methods for Details)


Change in Abiotic Edge Parameters (Mean ± SE) in the petenes Studied, Along the Matrix-Forest Patch Gradient. Areas Shown for the Matrix (Shaded Area; Negative Values) Outside the peten and the peten Edge (0-40 m) on the X-Axis
We found PAR values differed between sections of the matrix-patch transition (F(3) = 24.98, P < 0.05), decreasing toward the interior of the peten with the highest values starting at the matrix. In this case, contrast models confirmed that the PAR values in the matrix were significantly contrasting (from 2.6 to 6 times higher) with respect to the exterior, mid and interior edge sections in both types of peten (Table 1 and Figure 5).
The number of leaves intercepted per height interval (N = 13) was similar (P > 0.05) between types of peten, showing that the vertical distribution of the vegetation presents a consistent structure in both cases, with the lowest values of foliage at 1-2 m (Figure 6). Distribution of Vegetation (Foliage) by Vertical Strata in petenes With and Without Palm Extraction. The Horizontal Bars Represent the Mean Number of Leaves for Each Vertical Stratum Recorded by Type of Peten (See Vegetation Structure in Methods)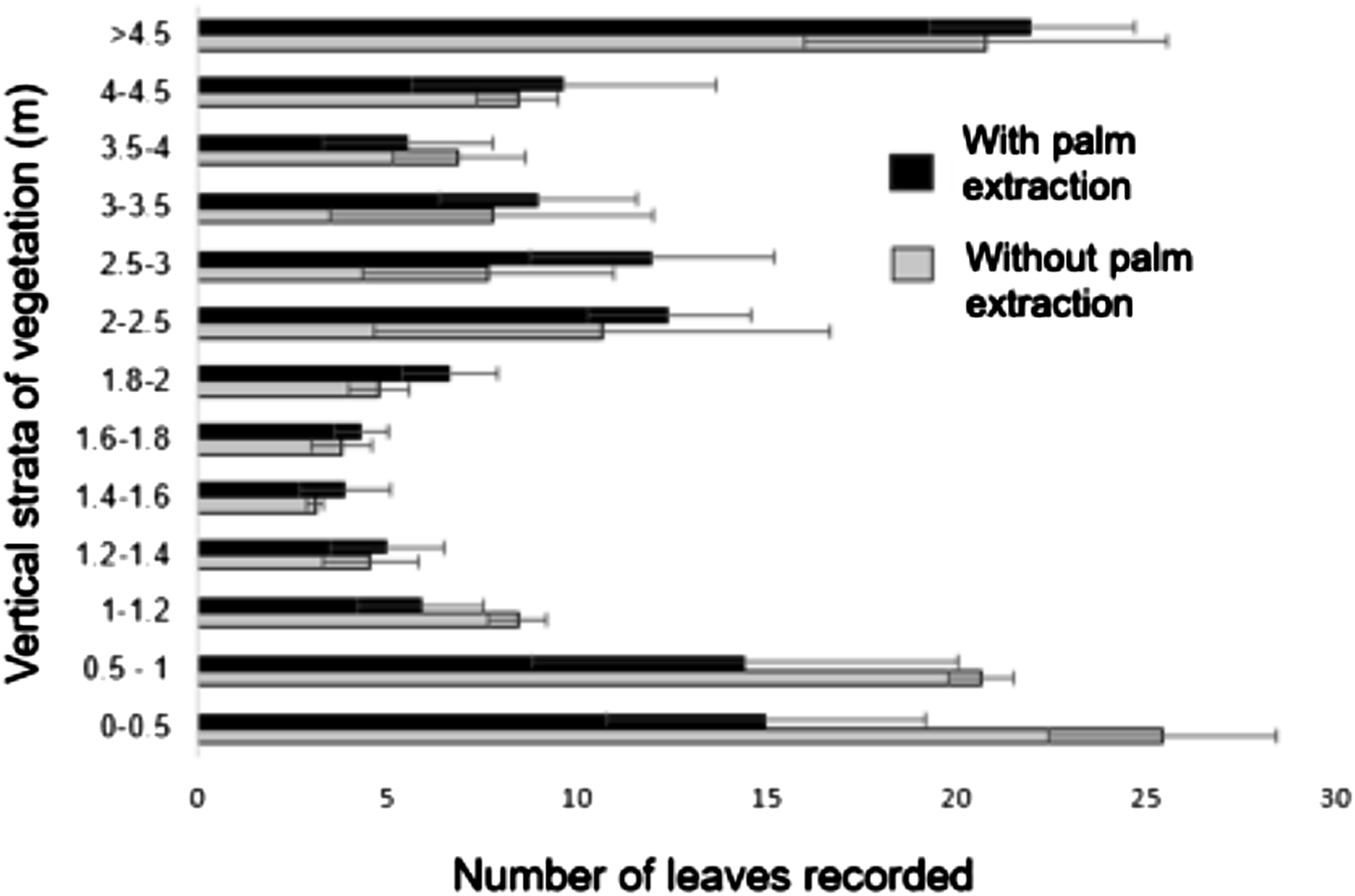
The ART-ANOVA showed that foliar interference along the matrix-patch transition was similar between types of peten (F(3) = 0.02, P > 0.05). Percentage values for this parameter notably increased toward the interior of the peten, with the lowest values starting at the matrix (Table 1 and Figure 7). In this case, contrast models confirmed that foliar interference in the matrix was at least 0.56 times lower with respect to the exterior, mid and interior edge sections, in both types of peten (Table 1 and Figure 7). As expected, PAR and foliar interference were inversely correlated along the matrix-patch transition (r = -0.86; t (22) = 8.08, P < 0.01). Change in Foliar Interference Percentage (Mean ± SE) in petenes With and Without Palm Extraction Along the Matrix-Forest Patch Gradient. Areas Shown for the Matrix (Shaded Area; Negative Values) Outside the peten and peten Edge (0-40 m) on the X-Axis
Microclimatic Parameters Across the Forest Edge Between Seasons
In petenes with palm extraction, the repeated-measures ART-ANOVA showed that there was a similar variation for temperature and relative humidity (F(3) = 0.36, 0.76, P > 0.05 respectively) along the matrix-patch transition between the rainy and dry seasons (Table 1 and Figure 8). Notably, only relative humidity values showed the lowest values for the dry season. Seasonal Change in Abiotic Edge Parameters (Mean± SE) for petenes With Palm Extraction Along the Matrix-Fragment Gradient. For This Comparison, the Records Were From Rainy Season (after Harvesting) and the Dry Season (before Harvesting) at Each Site (n = 3). Areas Shown for the Matrix (Shaded Area; Negative Values) Outside the peten and peten Edge (0-40 m) on the X-Axis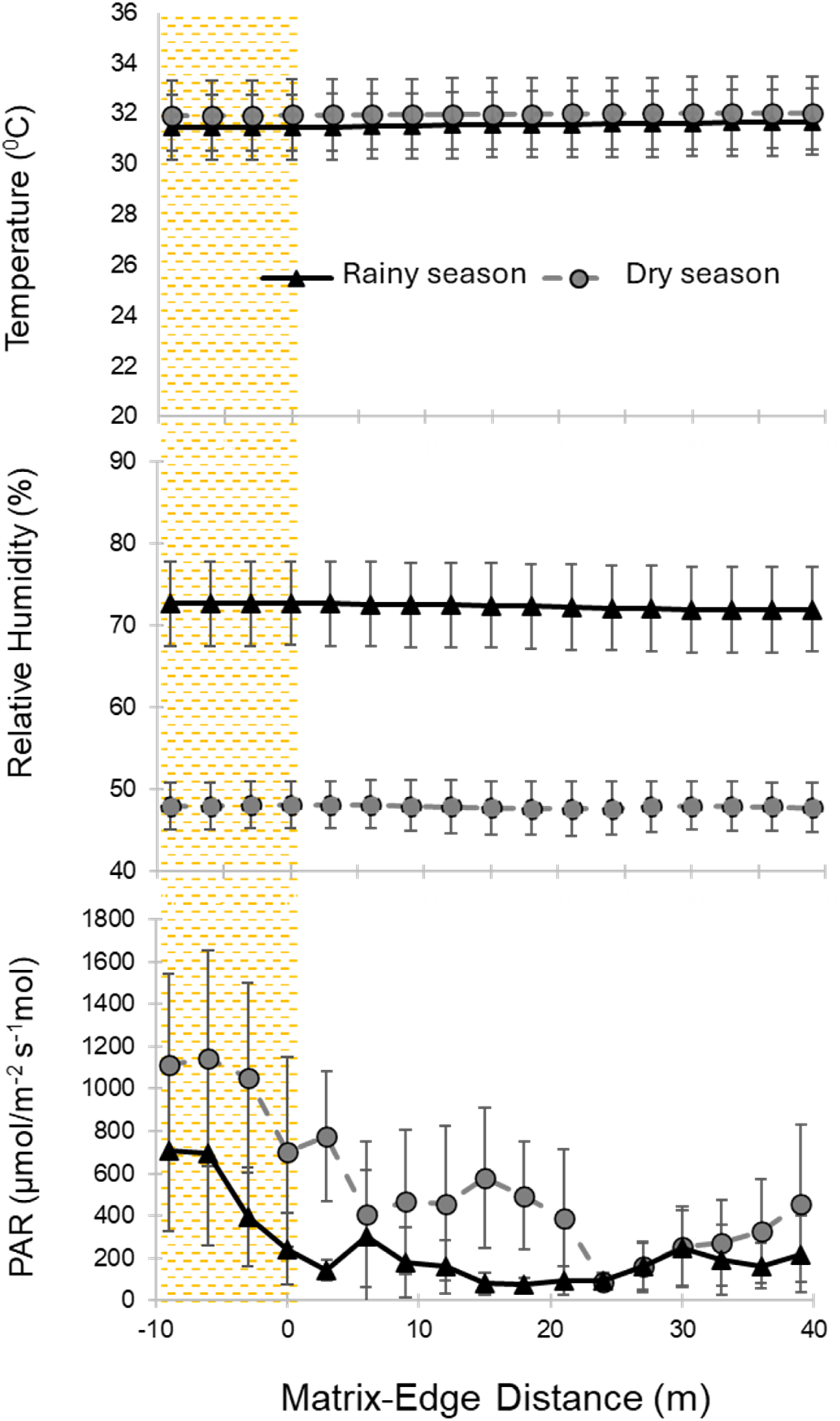
For this inter-seasonal comparison, PAR values were notably different along the matrix-patch transition (F(3) = 8.8 P < 0.05). For both seasons, PAR decreased toward the interior of the peten with the highest values starting at the matrix, particularly in the dry season. Contrast models confirmed that PAR values in the matrix were highly contrasting (from 1.9 to 6 times higher) compared to the exterior, mid and interior edge sections, in the rainy and dry seasons (Table 1 and Figure 8).
Along the matrix-patch transition, foliar interference showed significant differences between seasons (F(3) = 7.2, P < 0.05), presenting the lowest values in the dry season (Table 1 and Figure 9). For this inter-seasonal comparison, contrast models confirmed that foliar interference in the matrix was at least 0.6 times lower with respect to the exterior, mid and interior edge sections. For this parameter, a seasonal contrast was found at the mid edge in petenes with palm extraction (Table 1 and Figure 9). Seasonal Change in Foliar Interference Percentage (Mean ± SE) in petenes With Palm Extraction Along the Matrix-Fragment Gradient. For This Comparison, the Records Were From Rainy Season (After Harvesting) and the Dry Season (Before Harvesting) at Each Site (n = 3). Areas Shown for the Matrix (Shaded Area; Negative Values) Outside the peten and peten Edge (0-40 m) on the X-Axis
Discussion
Our study showed the variation in microclimatic conditions in the matrix-patch transition for the naturally formed petenes for the first time. Our findings revealed that the edge influence in petenes is primarily associated with PAR, which decreases by as much as 84%, highlighting a mid-edge (ca. 12 m from the border), from the matrix towards the interior of the forest patch, even in those sites with palm harvesting (see Figure 5). Since PAR and foliar interference were inversely correlated, the edge influence associated with PAR can be explained by structural changes in vegetation starting at the border of the peten (see Figure 7).
The inverse correlation between PAR and foliar interference may be key for understanding possible relations between vegetation changes (associated with palm extraction) and changes in others edge parameters. The vertical distribution in petenes shows that most leaves are found in the lower and upper strata of the peten (see Figure 6). If leaves are removed from the middle strata, the passage of light increases, which would cause significant variations in the abiotic parameters (Martínez-Ramos et al., 2016), particularly in PAR, but there is still cover in the upper strata where most of the vegetation is found. This applies to the sites with palm extraction, given that in this species, while of variable height, fronds are usually harvested from plants no taller than three meters. This contributes to explaining why no significant differences were found for PAR between sites with and without palm extraction (as shown in Figure 5).
In contrast to temperature, in petenes with and without palm harvesting, trends for PAR and relative humidity (to a lesser degree) along the matrix-patch transition were consistent with previous studies for the forest edge, which reported that microclimatic parameters are expected to vary gradually from the matrix towards the patch interior (Brothers & Spingarn, 1992; Kapos, 1989; Williams-Linera, 1990). The scanty variation along the gradient for temperature (and relative humidity) in both types of peten may be associated with the period of field measurements (12:00-15:00 hrs.), when these parameters are more homogeneous during the day. This latter was a criterion defined a priori to avoid possible contrasts along the gradient associated with the time of day being confused with abiotic changes of the edge, reported for parameters such as temperature and relative humidity (Laurance et al., 2002; Murcia, 1995; Ries et al., 2004). We consider this situation to be the correct method for monitoring edge changes in standard periods and with the daylong variation in the abiotic parameters of interest. Furthermore, the above made it possible to more precisely evaluate the magnitude and penetration of the edge influence based on a parameter such as PAR, which is strongly related to structural changes in vegetation in the matrix-patch transition zone.
The coastal zone of the Yucatan Peninsula, a structurally complex matrix with the presence of mangrove belts, would provide less contrasting differential conditions with the forest patch (Debinski, 2006; Murcia, 1995; Porensky & Young, 2013), forming an attenuated and softened edge influence (i.e., due to temperature and PAR) in petenes, similar to the edges found, for example, in forest fragments embedded in the Everglades of South Florida (Sternberg et al., 2007). That ecosystem has many similarities to the structure and spatial configuration of the petenes ecosystem (CONANP, 2006). Mangroves (as a vegetation type) provide the habitat with resilience and resistance to climate changes (Alongi, 2008; Marois & Mitsch, 2015), giving rise to properties that attenuate abiotic parameters (such as temperature and relative humidity) recorded in this study.
Another distinctive characteristic of petenes that may act as a buffer for temperature and relative humidity at the edge is the presence of sinkholes of fresh water in the interior of the forest patch, common to the region (Rico-Gray, 1982; Munguía-Rosas & Montiel, 2014). These sources can form bodies of water in the interior of the petenes and generate a humid environment throughout the year, as water flows toward different areas of the patch. Following the model proposed by Porensky and Young (2013), the above would cause little variation in terms of temperature and relative humidity along a matrix-patch gradient, with the existence of an effect from inside to outside the peten counteracting the differences between the matrix and fragment.
In fragmented wetlands, hydrology (e.g.; variations of water salinity) and seasonal flooding modulates the edge influence, modifying microclimatic edge parameters with respect to the dry season but without the complete disappearance of the matrix-fragment variability (Watts & Kobziar, 2015). Additionally, in forest fragments surrounded mainly by grassland like those of our study, edge penetration measured by parameters such as temperature and relative humidity can change in the dry season (Hennenberg et al., 2008). Our results were consistent with the previous expectations only for relative humidity, which notably decreased (but with the same trend) along the matrix-patch transition during the dry season, compared to the rainy season. Together with temperature, this parameter showed a similar seasonal variation from the matrix to the interior of the forest patch for petenes with palm harvesting (see Figure 8).
In both seasons, photosynthetically active radiation, the central parameter in this study, decreased toward the interior of the peten with the highest values starting at the matrix, particularly in the dry season (see Figure 8). Our results showed that trend in this parameter along the matrix-patch transition, with the edge influence due to PAR being maintained to the forest interior in both seasons. This finding can be explained by the semideciduous nature of the forest and the presence of mangrove species in petenes (Munguía-Rosas et al., 2014; Rico-Gray, 1982). Trees such as R. mangle, M. zapota, M. brownei, L. racemosa, A. germinans dominating the canopy and B. tubiflora and S. japa dominating the understory provides a similar vertical structure and forest coverage in petenes throughout the year (see Figures 6 and 9, respectively).
In petenes, the edge influence associated to PAR, which is mainly related to structural changes in vegetation in the matrix-patch transition (Brothers & Spingarn, 1992), can be appreciated starting from the “mid edge” in the dry season, while the contrasts appear from the “exterior edge” in the rainy season (see Figure 8). The edge records in the rainy season showed the expected edge influence in petenes due to the greater humidity resulting from flooding. This promotes notable characteristics in terms of vegetation structure in the forest patches, without a notable differential contrast with the matrix, as might be the case in the dry season.
In the naturally fragmented ecosystem studied, the edge does not behave abruptly as in other particularly anthropogenically modified forest ecosystems (Laurance et al., 2002, 2007; Murcia, 1995; Porensky & Young, 2013). In addition to the forest constitution and the wetland environmental conditions, the age of the edge seems to be a relevant factor for understanding the edge influence in petenes. Considering that petenes are the result of long-term geo-ecological processes (Montiel et al., 2006; Munguía-Rosas & Montiel, 2014), for these forest patches we can assume that the changes relating to the edge are largely stabilized, although external changes in land use (such as fire or the development of adjacent vegetation) could disrupt the balance, which would equate to a post-closure phase of greater than five years (Laurance et al., 2002). As such, in petenes the existence of an attenuated edge is proposed that is presumably associated with a dynamic equilibrium of the forest patch (Sternberg et al., 2007) that may be promoted by (a) long-term geo-ecological processes of the forest patches present in the coastal wetland, (b) a gradual reduction in the changes in vegetation between the matrix-peten transition, (c) a highly structured matrix with high landscape stability, and (d) the presence of water sources in their interior.
Finally, we conclude that the scant variation recorded for temperature and relative humidity, as well as changes in PAR from the matrix to the interior of petenes, suggests an edge influence modulated by the patch’s forest structure and the presence of mangroves, even in those petenes with palm harvesting.
Implications for Conservation
The abiotic parameters studied in petenes suggest an edge influence mainly associated to presence of mangroves on the patch’s periphery that appears to promote the ecological stability and persistence of petenes in this important naturally fragmented Mesoamerican ecosystem. Potential anthropogenic alterations added to seasonal variation could modify the abiotic edge parameters of the forest patches in the wetland during the dry season, but with “normal” conditions recovering in the rainy season. This would imply a dynamic ecosystem highly resistant to extractive activities (i.e.; palm extraction) on a local scale, meaning that these activities could be promoted selectively in the context of the development and conservation appropriate to a biosphere reserve.
Footnotes
Acknowledgments
The authors thank Armando Rojas for your help in collecting data in petenes and CINVESTAV-Merida for the logistical support provided for the fieldwork in wetlands. Special thanks are given to Franco Rojas for the aerial views of petenes using a Drone and Marcos Briceño for his help with complementary data analysis. Carlos N. Ibarra and four anonymous reviewers provided valuable suggestions to the final version of this manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was partially funded by a Master’s scholarship (704309) awarded to the first author (SGH) by Mexico’s Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI). The authors also thank CINVESTAV-Merida for the partial funding.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.