
Abstract
Keywords
Introduction
Habitat loss and fragmentation are, and will remain, leading drivers of the biodiversity crisis—ranging from local population extinctions to the disruption of ecological functions (Cardinale et al., 2012; Powers & Jetz, 2019). In response, maintaining and restoring connectivity in agricultural and human-modified landscapes has become a key strategy for wildlife conservation (Bennett & Blanch, 2004; Rudnick et al., 2012).
Well-preserved forest remnants are typically found within protected areas, which are established either to preserve a broad range of native species or to protect specific species or groups (Figueroa & Sánchez-Cordero, 2008; Guevara & Laborde, 2008). However, in fragmented landscapes where natural patches are surrounded by high levels of anthropogenic disturbance, restoring connectivity between these protected areas is critical for long-term biodiversity conservation (Halffter, 2007; Possati Vieira De Matos et al., 2021).
The concept of habitat corridors is based on the idea that connectivity enables movement between habitat patches, allowing individuals to disperse, access essential resources, and maintain gene flow—ultimately reducing the risk of inbreeding and local extinction (Frankham et al., 2002; Hilty et al., 2020; Lande, 1988; Rouget et al., 2006). However, the actual movement of wildlife across transformed landscapes remains uncertain and is often not supported by empirical evidence (Bennett & Blanch, 2004; Hofmann et al., 2021; Laner et al., 2024).
Buffer zones—transitional areas between natural habitats and human-modified landscapes—can either hinder or facilitate species movement depending on the degree of human disturbance (Bennett & Mulongoy, 2006; Guevara & Laborde, 2008). The effectiveness of biological corridors depends on the ability of species to move between habitat remnants, particularly protected areas.
In this study, we assess the permeability of the buffer zone adjacent to La Amistad International Park (PILA) for medium and large mammals by analyzing species presence across various zones and vegetation types. We also examine landscape and anthropogenic factors influencing species distribution, providing a foundation for evaluating the effectiveness of the La Amistad–Osa Biological Corridor in a human-dominated landscape.
Methods
Study Area
The Talamanca Range is one of the most biodiverse regions in Costa Rica and among the last remaining continuous habitats for wildlife in the country. It is considered a conservation priority for large mammals (de la Torre et al., 2017; González-Maya et al., 2016a, 2016b). Located in southern Costa Rica, the range spans approximately 650,000 ha, of which 197,527 ha are protected by La Amistad International Park (PILA). On the park’s Pacific slope, elevations range from 1,500 to 3,500 meters above sea level, covering 25,000 ha primarily composed of mountain cloud forest (Asociación Conservación de la Naturaleza, 2005; MINAET, 2012). Due to its ecological importance, the area has been designated both a World Heritage Site and an UNESCO Biosphere Reserve (Asociación Conservación de la Naturaleza, 2005).
The La Amistad–Osa Biological Corridor (AmistOsa), located in southeastern Costa Rica, covers 115,809 ha—approximately 10% of the Puntarenas province. First proposed in 1986 and later established in 2006 to reconnect the Talamanca Range with the Osa Peninsula over a fragmented stretch of nearly 50 km. The corridor aims to restore connectivity between PILA and forest remnants in the Osa region, including Corcovado National Park (Céspedes-Agüero, 2008; SINAC, 2018).
Field Surveys
Field sampling was conducted between July 2013 and January 2014, covering both the rainy (July–September) and dry (November–January) seasons. To estimate spatial variation in the community of medium- and large-sized terrestrial mammals outside PILA, we deployed 24 camera trap stations along four contiguous bands extending outward from the park boundary. Each band was 3.3 km wide and perpendicular to the PILA core boundary, spanning up to 10 km into the buffer zone. Six stations were installed per band, with the first band (<3 km from the boundary) located inside the park and the remaining three bands outside it. This design encompassed dispersal ranges from highly mobile species (e.g., Panthera onca) to those with limited dispersal (e.g., Philander opossum).
Each station was equipped with two motion-triggered cameras (LTL ACORN 6210 MC, PTC Technologies), placed approximately 200 m apart to maximize detection probability (Figure 1). Cameras were active for 15 days per station, per season, and were installed exclusively in forest remnants. These remnants are largely confined to riparian areas within the buffer zone. As this zone consists mainly of privately owned farms, site selection was determined by both ecological relevance and landowner permission. Progressive 3.3 km Distance Bands From the Boundary of La Amistad International Park (PILA). The First Band (A1) is Located Within the Park Boundary, While the Remaining Three Bands Lie Outside the Park: 0–3.3 km (A2), 3.3–6.6 km (A3), and 6.6–10+ km (A4) From the Park Boundary. Vegetation Types Within Each Band Are Grouped Into Two Categories: “Disturbed Vegetation” (Including Disturbed Forests, Crops, Forest Plantations, and Cattle Pastures) and “Natural Vegetation” (Including Forest, Wetlands, and Páramo). The Inset Map Shows the Study Area Within Costa Rica.
Landscape Structure and Vegetation Classification
We characterized landscape structure using ArcGIS (ESRI, 1999) and Fragstats 4.0 (McGarigal et al., 2002). For each forest patch, we measured location, area (ha), minimum patch width, and isolation (Euclidean distance to the nearest patch), as well as proximity to crop fields and human settlements. Land cover data were derived from the 2008 Costa Rica land cover layer provided by the Atlas de Costa Rica (Ortíz-Malavasi, 2009).
Vegetation types within each band were grouped into two broad categories based on structural similarity to the primary forest in PILA: (1) Natural Vegetation: Areas dominated by native ecosystems with complex structure, including primary and secondary forests, wetlands, and páramo. (2) Disturbed Vegetation: Areas significantly modified by human activities, including disturbed forests (e.g., logged or fragmented areas), crop fields (mainly coffee), forest plantations (non-native monocultures), and cattle pastures.
These categories differ in structural complexity and resource availability, with disturbed habitats typically offering fewer food sources, shelter, or breeding sites (Chiarello, 1999) and posing greater barriers to forest-dependent mammal movement (Bennett & Saunders, 2010). The percentage of natural vegetation decreased progressively with distance from the park: 94% in Band A1, 62% in A2, 46% in A3, and 45% in A4.
Community Interviews
We conducted semi-structured interviews with residents in 13 towns across four districts: (1) Biolley: Altamira, Bajo Coto, El Carmen, Colorado, Sábalo; (2) Buenos Aires: San Luis; (3) Pittier: Aguascalientes, Cotón, Fila Tigre, Fila Naranjo, Las Marías; (4) Potrero Grande: San Isidro, Sanci.
These interviews gathered local knowledge on mammal presence and perceived threats, including hunting, human-wildlife conflict, and resource use. Questions also addressed attitudes toward wildlife and conservation (see Annex 1).
Analysis
We assessed spatial variation in community attributes of medium- and large-sized mammals across the four sampling bands using camera trap data. To evaluate spatial autocorrelation among stations, we applied a Complete Spatial Randomness (CSR) test using the quadrat method, implemented in the spatstat package in R (Baddeley et al., 2022), using only one location per station.
Species richness per band was compared using one-way ANOVA, treating each camera station as a replicate. Differences in species composition among bands were assessed using the Jaccard similarity index and Sørensen similarity coefficient. Because sample sizes varied across stations, we conducted 100 random resampling iterations to standardize comparisons.
To evaluate the effects of human disturbance and landscape structure on habitat permeability for individual species, we used generalized linear mixed models (GLMMs) with a Poisson error distribution and a log-link function. Models were run only for species with more than five independent detections per season. Capture frequency (i.e., the number of individuals detected during the 15-day camera operation) was used as a proxy for site use and, by extension, for habitat permeability.
Fixed effects included season, distance to crops, number of hunters, forest patch area, isolation from other vegetation, and the width of arboreal vegetation at each camera station. Random effects accounted for the nested structure of the data, with camera stations nested within season to control for potential spatial and temporal autocorrelation (each station was sampled in both dry and rainy seasons).
Model selection was performed using a backward stepwise approach (Burnham & Anderson, 2016), testing all possible combinations of fixed variables. The best model for each species was identified using the Akaike Information Criterion (AIC), selecting the model with the lowest AIC value (Portet, 2020). Model assumptions of homoscedasticity and normality were evaluated for all final models.
All statistical analyses were conducted in R version 2.15.3 (R Development Core Team, 2011), using the lme4 package (Bates et al., 2011).
Results
Species Recorded
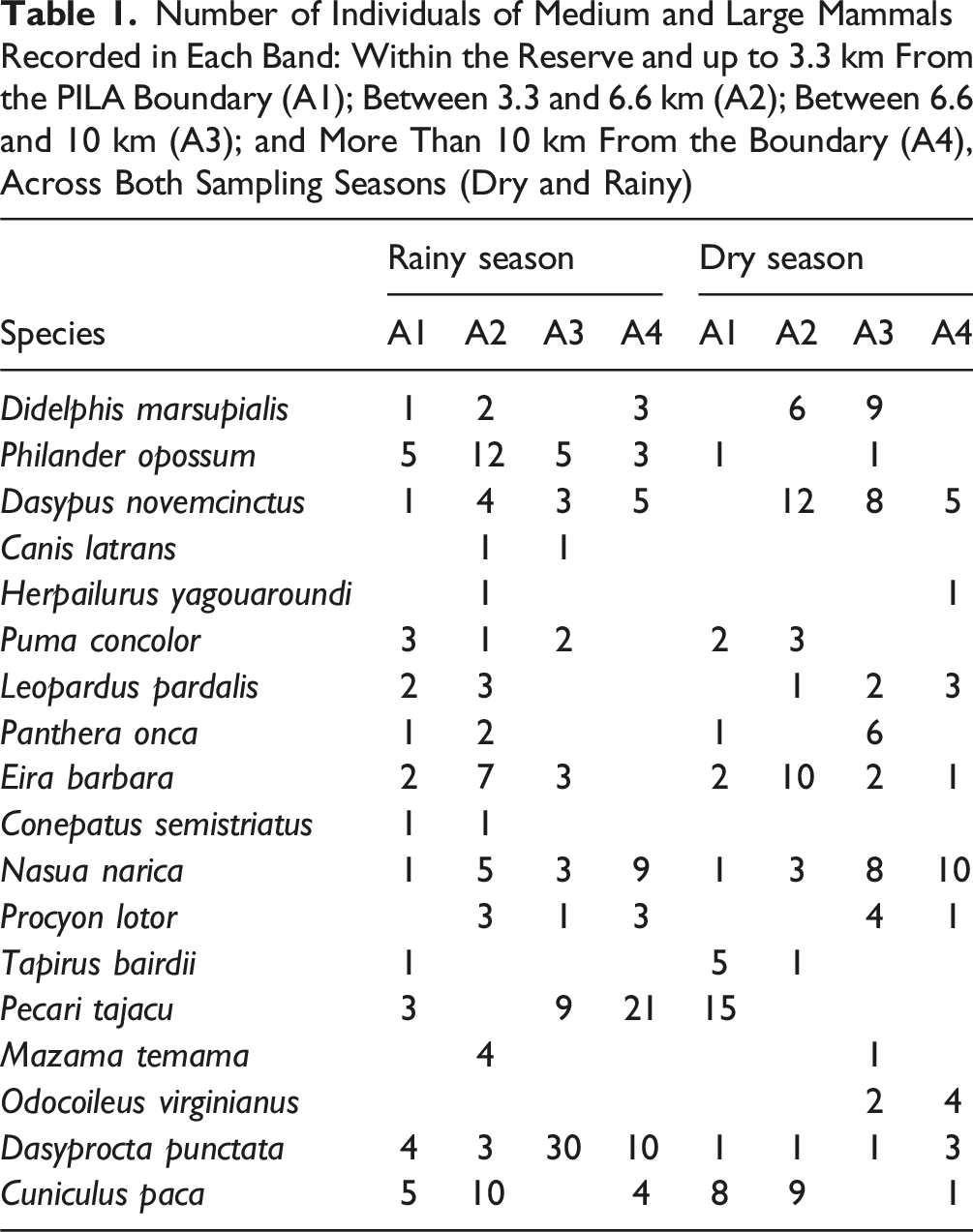
Number of Individuals of Medium and Large Mammals Recorded in Each Band: Within the Reserve and up to 3.3 km From the PILA Boundary (A1); Between 3.3 and 6.6 km (A2); Between 6.6 and 10 km (A3); and More Than 10 km From the Boundary (A4), Across Both Sampling Seasons (Dry and Rainy)
Local Knowledge and Hunting Pressure
Interviews with local residents identified the most frequently hunted species as paca (Cuniculus paca), collared peccary (Pecari tajacu), Central American agouti (Dasyprocta punctata), nine-banded armadillo (Dasypus novemcinctus), and white-tailed deer (Odocoileus virginianus), primarily hunted for bushmeat. Reported hunting intensity ranged from 1 hunter in Las Marías to 35 in Cotón, with frequent mention of poaching by outsiders. Several species were reported to have disappeared locally, including jaguar (Panthera onca), puma (Puma concolor), Baird’s tapir (Tapirus bairdii), white-lipped peccary (Tayassu pecari), and giant anteater (Myrmecophaga tridactyla)—with some sightings limited to specific areas like Fila Tigre, Cotón, Agua Caliente, and within PILA.
Residents noted that most mammal movements occur along riparian corridors, particularly in forested remnants near the Canasta, Cotón, Platanillal, Singri, and Coto Rivers.
Human-Wildlife Conflict
Problematic species reported across multiple localities included the white-nosed coati (Nasua narica), white-tailed deer, raccoon (Procyon lotor), and animals locally referred to as tolomuco (a term used for both Eira barbara and Herpailurus yagouaroundi) and caucel (used for Leopardus wiedii and Leopardus tigrinus). These were mostly associated with crops and poultry predation. In Fila Tigre, Agua Caliente, and Cotón, jaguar and puma were also reported to prey on cattle and sheep.
Changes in Mammal Assemblage Composition and Structure
Fourteen species were recorded in Band A1 (within the park), while 16, 15, and 11 species were recorded in Bands A2, A3, and A4, respectively; all located outside the park. Species richness was lowest in A4, with similar richness levels in the other three bands.
Average Jaccard Similarity Index and Average Sørensen Similarity Coefficient for Each Sampled Band (A1, A2, A3, and A4) Within and Outside La Amistad International Park, Across Both Seasons. One Hundred Random Repetitions Were Performed to Standardize Sample Size

The Sørensen similarity coefficient also reflected increasing differences with distance, indicating variation not only in species composition but also in assemblage structure (Table 2).
Effects of Human and Landscape Variables on Permeability
The Complete Spatial Randomness (CSR) test, based on the quadrat method, showed no significant spatial autocorrelation among stations (χ2 = 22, df = 24, p = 0.8415).
Summary of Generalized Linear Mixed Models (GLMMs) for the Six Species That Showed Statistically Significant Results. The Best-Fitting Model for Each Species was Selected Based on the Akaike Information Criterion (AIC). *Indicates Statistical Significance

Discussion
Limitations of Using Capture Frequency as a Proxy for Permeability
While camera traps offer crucial insights into species presence, directly translating capture frequency into a measure of habitat permeability presents inherent challenges. Several factors can influence these detection rates, including the species’ unique likelihood of being detected, localized population densities, the size of their home ranges, or even focused use of specific habitat patches for foraging or refuge. Consequently, a high number of detections at a given station does not automatically signal extensive landscape-level movement or dispersal (Cushman et al., 2013; Zeller et al., 2012). Furthermore, camera traps are generally ill-equipped to capture rare dispersal incidents or broader movement patterns, both of which are vital for grasping functional connectivity.
Even with these limitations, our findings provide a crucial initial look at how species utilize the buffer zone. They also point to which species seem more adept at handling or even benefiting from altered landscapes. While we are not directly measuring ecological connectivity here—the way you might with gene flow studies or telemetry (Hilty et al., 2020)—our work does build a foundational understanding of how this corridor functions within a highly fragmented, human-impacted area.
Changes in Mammal Assemblage Across the Buffer Zone
Buffer zones within biosphere reserve frameworks are generally seen as transitional areas. They are designed to reduce anthropogenic impacts and support biodiversity conservation beyond protected area boundaries, ideally helping native species move through human-modified landscapes (Guevara & Laborde, 2008). Our research, though, reveals a distinct shift in mammal community composition and structure with increasing distance from the PILA core protected area.
We observed a noticeable drop in species richness in the outermost zone (Band A4), with generalist species becoming more prevalent further from the park. In marked contrast, large-bodied species like Tapirus bairdii, Panthera onca, and Puma concolor were only detected in a private old-growth forest reserve right next to PILA. This suggests that most of the buffer zone is not truly functionally permeable for these wide-ranging and disturbance-sensitive animals.
Influence of Landscape Attributes and Human Disturbance
Of all carnivores, only the tayra (Eira barbara) was detected often enough to allow for habitat use modeling. Its capture frequency showed a positive link with forest patch area and riparian vegetation width, while decreasing with proximity to crops. This aligns well with prior work highlighting the tayra’s generalist behavior and its capacity to utilize both forested and disturbed environments (Eisenberg, 1989; Soley, 2012). Nevertheless, the clear preference for larger, structurally complex habitats points to a crucial need: even highly adaptable carnivores still depend on core forest elements for their continued presence.
Herbivores like Pecari tajacu and Dasyprocta punctata showed a positive response to wider riparian vegetation and less isolated patches, particularly where predators were scarce. Their adaptability and propensity for using edge habitats and agricultural resources likely explain their continued presence in these altered landscapes. Despite this, ongoing hunting pressure—which surfaced in our interviews—could still be limiting how widely they’re distributed.
We recorded omnivorous species like the white-nosed coati (Nasua narica), raccoon (Procyon lotor), and opossum (Philander opossum) more frequently in disturbed areas. Their success in fragmented environments is likely boosted by their generalist diets, high adaptability, and what seems to be predator release (Guerrero et al., 2000; Hamilton, 1936; Romero-Balderas et al., 2006). Their consistent presence near human settlements and agricultural zones really underscores the matrix’s permeability for these disturbance-tolerant species.
Corridor Effectiveness and Recommendations for Restoration
The La Amistad-Osa Biological Corridor was set up to boost both altitudinal and horizontal connectivity between PILA and the Osa Peninsula (MINAET, 2012). However, our findings suggest that current land-use patterns, especially the widespread intensive agriculture and deforestation in mid- and low-elevation areas, create significant obstacles for large mammal movement. The clear drop in species richness and changes in composition as we move further from PILA indicate that, for forest-dependent and large-bodied species, functional connectivity is currently quite restricted.
Focusing restoration efforts on increasing forest cover, especially through riparian reforestation, could be a good way to boost corridor permeability. These linear strips of habitat are among the last natural vegetation corridors left in the region (Fremier et al., 2015; Guevara et al., 2004; Harvey et al., 2005). If we expand structurally complex habitats—like those in the private reserve where we found large mammals—we might just turn parts of the buffer zone from obstacles into useful steppingstones for wildlife to move through.
Connectivity
Functional connectivity hinges on wildlife’s ability to move between habitat patches in fragmented landscapes (Bennett & Mulongoy, 2006). Our results indicate that while the buffer zone does permit movement for generalist, medium-sized species, it just is not permeable enough for large mammals. Animals like P. onca, P. concolor, and T. bairdii need large, well-connected forest patches to survive and disperse (Carbajal-Borges et al., 2014; De Angelo et al., 2011), and we only observed them in those protected, less-disturbed areas
Our observations show the smallest patch width where these large mammals were found: 190 m for P. onca, 1,116 m for P. concolor, and 646 m for T. bairdii. It is important to view these values as helpful reference points, not rigid thresholds, until further verified. Confirming these minimum habitat requirements and identifying specific movement corridors will require future studies, ideally incorporating genetic data or animal radio-tracking.
The lack of top predators can trigger a cascade of ecological impacts, such as mesopredator release and over-browse by herbivores. These effects, in turn, can significantly alter vegetation structure and regeneration dynamics (Rawinski, 2008; Terborgh et al., 1999). Consequently, restoring ecological connectivity becomes crucial not only for species conservation but also for preserving overall ecosystem function.
Despite the time elapsed since our fieldwork, the observed patterns hold significant relevance. The buffer zone around PILA, as it stands, supports only a subset of the region’s mammal fauna, predominantly generalist and medium-sized species. The distinct shifts in community composition and structure that emerged with increasing distance from the core area underscore an urgent call for targeted conservation efforts. Specifically, reforestation—especially within riparian corridors—and the expansion of structurally complex forest patches are essential steps to enhance landscape permeability and fulfill the original objectives of the La Amistad-Osa Biological Corridor. Our work provides a robust baseline for future studies and management strategies aimed at restoring and preserving mammal connectivity in southern Costa Rica.
Implications for Conservation
Although proposed way back in 1986, the La Amistad-Osa Biological Corridor’s effectiveness—meant to link La Amistad International Park (PILA) with the Osa Peninsula—had never been empirically measured for medium- and large-sized mammals until our study.
Our findings show that the buffer zone next to PILA’s core area is still permeable for some medium-sized, disturbance-tolerant mammal species. However, large forest-dwelling carnivores and herbivores—those more sensitive to habitat fragmentation and human pressures—were mostly missing outside the well-preserved forest reserve. Their limited presence strongly links to isolated forest remnants, smaller patch sizes, and greater distance from the core protected area, all factors that restrict dispersal and population viability.
To truly boost functional connectivity across this human-dominated landscape, especially within the buffer zone, targeted restoration is crucial. We should prioritize reforestation, particularly expanding riparian forest corridors, and focus on the ecological restoration of degraded patches. These steps will increase both forest cover and structural complexity. In parallel, implementing environmental education programs and compensation schemes for farmers facing wildlife-related losses could foster coexistence and lessen conflict, ultimately making the landscape more permeable for wildlife.
Our study provides a vital baseline for understanding the habitat conditions essential for mammal movement between two of Costa Rica’s most significant remaining forest strongholds. The findings directly support and inform ongoing governmental and private initiatives aimed at creating a truly functional biological corridor within the “AmistOsa” region.
Footnotes
Acknowledgments
Partial financial support was provided by the Universidad Autónoma Metropolitana-Lerma (Project 54302002). Funding for camera traps was provided by the Programa de Mejoramiento al Profesorado (PROMEP), UAM-PTC-341. The Consejo Nacional de Ciencia y Tecnología (CONACYT) awarded a postgraduate scholarship to the first author. We thank the residents of the study area, the National System of Conservation Areas of Costa Rica (SINAC), the La Amistad-Pacífico Conservation Area, and ProCAT Colombia and ProCAT Costa Rica and their partners for their invaluable help and support. We thank Kinley Reagan for her support on language reviewing and editing on the final stages of this paper.
Ethical Considerations
This study did not involve the capture, handling or interference of animals.
Author’s Contributions
AMMG, RL, JFGM, EMM, and AGR conceptualized the research and defined the methodology. AMMH conducted the fieldwork and curated the data. AMMG, EMM, JFGM, and JL conducted the formal analysis. AMMG, RL, JFGM, EMM, AGR, and JL contributed to the original draft. AMMG, JFGM, and RL reviewed and edited the final version of the manuscript.
Funding
Partial financial support was received from the Universidad Autónoma Metropolitana-Lerma (project 54302002). The Programa de Mejoramiento al Profesorado (PROMEP) UAM-PTC-341 provided funding for camera traps. The Consejo Nacional de Ciencia y Tecnología (CONACYT) awarded a postgraduate scholarship to the first author.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Appendix
Age ___ Sex ___
Education
Occupation