
Abstract
Habitat loss from forest conversion to agriculture threatens tropical biodiversity. Despite documented risk-avoidance behaviors, some species may adopt riskier strategies to gain access to food. Recent conversion of a protected area in southern Cameroon to an agro-industrial plantation coincides with increased sightings of forest elephants near human settlements, which is unusual and suggests a drastic change in their habitat use. This study aims to examine the influence of human activity on forest elephant habitat use and evaluate the effectiveness of two survey methods in documenting elephant and human occurrence. Twenty-one camera traps were deployed along the border between the declassified protected area and the community land, and reconnaissance walks were conducted between camera trap stations. Results from both methods were compared. Elephant occurrence tended to be negatively affected by human activity, and elephants were inactive during peak human activity. However, their presence near human settlements suggests a general risk-taking behavior in habitat use. Moreover, reconnaissance walks proved more effective than camera traps in providing a greater amount of data. This risky proximity to humans points to a complex trade-off between risk and access to food resources, where the nutritional benefits and easy access of crops and secondary forest resources may outweigh the perceived human-mediated risk. At the same time, elephants may adopt strategies to minimize direct interactions with humans. Further habitat fragmentation and human encroachment on wild areas are expected in the near future. As elephant presence near human settlements often lead to increased conflict, continued monitoring of elephant habitat use in human-dominated landscapes using efficient survey methods is crucial to design up-to-date and effective management and conservation strategies.
Introduction
Habitat loss through land-use change, particularly through the conversion of forest to agriculture, is one of the most prevalent threats to tropical biodiversity (FAO, 2020; Hald-Mortensen, 2023; Nunes et al., 2022; Perrings & Halkos, 2015). As wildlife habitat shrinks, animals may be forced to occupy areas in closer proximity with nearby human settlements and agricultural fields (Liu et al., 2017; P. Sharma et al., 2020), intensifying human-wildlife tensions and undermining conservation efforts (Dickman, 2010). Elephant movement patterns are driven by food and water distribution (Adams et al., 2022; Beirne et al., 2020; Blake, 2002; Blake & Maisels, 2023; Djoko, Weladji, Granados, et al., 2022), yet when elephant and human habitat overlap, human activity may outweigh the ecological determinants of their distribution (Blake, 2002; Blake & Maisels, 2023; Buij et al., 2007). Although elephant spatial risk avoidance through avoidance of areas and infrastructures associated with humans is well-documented (Blake et al., 2008; Buij et al., 2007; Gaugris et al., 2022; Laurance et al., 2006; Wrege et al., 2024), risk-taking behaviors often emerge in human-dominated landscapes as elephants venture into human areas and crop fields in search of food and water (Djoko, Weladji, & Paré, 2022; Gaynor et al., 2018; Mills et al., 2018; Poulsen et al., 2011; Shaffer et al., 2019; Tiller et al., 2021), when the easy access and nutritional benefits of crops and secondary forest resources may outweigh the risk of interacting with humans (Djoko, Weladji, Granados, et al., 2022; Mills et al., 2018; Tiller et al., 2021). Direct interactions with humans can present a risk to elephants, as they may be chased away or harassed by people protecting their farms (Nelson et al., 2003; Ram et al., 2021). However, elephants may mitigate this risk by adapting their activity patterns in areas associated with humans, such as by increasing night-time activity (Adams et al., 2022; Gaynor et al., 2018; Hahn et al., 2022; Smit et al., 2023; Tiller et al., 2021; Wrege et al., 2010, 2024).
In the Campo-Ma’an Technical Operational Unit (CMTOU) in southern Cameroon, the critically endangered forest elephant (Loxodonta cyclotis) faces a rapidly changing environment. In recent years, the area has come under significant pressure from the development of large-scale exploitation projects, including the establishment of an agro-industrial plantation in a previously protected area in the Campo subdivision (African Wildlife Foundation, 2022; Engolo et al., 2024; Forje et al., 2021), which had revealed to be home to an important density of forest elephants (African Wildlife Foundation, 2022; Ayuk et al., 2023; Beukou-Choumbou et al., 2021). Prior to the establishment of the agro-industry in the CMTOU, Djoko, Weladji, Granados et al. (2022) found that elephant relative abundance was negatively affected by human activities such as hunting and logging. However, elephants were more likely to be present in areas with higher human presence, suggesting a trade-off between risk associated with human presence and access to food resources in secondary forest. Elephant temporal response to risk in this area has not yet been studied. The establishment of the plantation in this human-dominated landscape is bound to exacerbate existing or generate new human-wildlife dynamics (Ayuk et al., 2023). This is already suggested by the increasing number and frequency of reports of elephants venturing into community crop fields (African Wildlife Foundation, 2022; Mimeault, 2025; Urbain Nzitouo, Head of the Community and Participatory Management Service of MINFOF, personal communication, May 2023), which points to a drastic shift in their habitat use patterns as they adapt to the new landscape. The main aim of this study is to explore whether elephants in the Campo subdivision of the CMTOU exhibit spatial and temporal risk-avoidance or risk-taking strategies in response to humans. To do so, we will examine the influence of human activity on forest elephant occurrence and assess the overlap in diel activity patterns between the two species.
Monitoring wildlife in densely forested habitats may be challenging using direct observations due to limited visibility in thick understory and the elusive nature and sparse distribution of many species (Zwerts et al., 2021). Over the past decades, camera traps have proven to be a valuable tool in tropical forests by providing direct observations that would otherwise be challenging to collect (Chakraborty et al., 2021; Collins & Weladji, 2024; Djoko, Weladji, Granados, et al., 2022; Sanderson & Trolle, 2005; Tanwar et al., 2021; Wearn & Glover-Kapfer, 2019). Research in Central African rainforests also highlight the benefits of reconnaissance walks (also referred to as “recces”), to enhance data collection efficiency, notably on forest elephant populations (Blake et al., 2007; Fai et al., 2022; Gaugris et al., 2022; Hedges, 2012; Walsh & White, 1999; Whytock et al., 2021). Given the varying results regarding the effectiveness and accuracy of camera traps compared to other survey methods, depending on species, objectives, cost-to-time ratio, and environment (Gaugris et al., 2022; Nuñez et al., 2019; Wearn & Glover-Kapfer, 2019), our second research objective is to compare results from camera trapping with those from recces to evaluate the effectiveness of each method in documenting elephant and human occurrence. Understanding how forest elephants use space and interact with humans in light of recent rapid habitat loss is crucial for the development of up to date and effective management and conservation strategies (Shaffer et al., 2019), and selecting the appropriate method to report on it given the research objectives and resources at hand is imperative (Wearn & Glover-Kapfer, 2019; Zwerts et al., 2021).
Materials and Methods
Study Area
The study was carried out in the Campo subdivision (∼300,000 ha) of the Campo-Ma’an Technical Operational Unit (CMTOU) (771,000 ha) in southern Cameroon (Figure 1). This subdivision is located on the Atlantic coast, north of the mouth of Ntem River which marks the border with Equatorial Guinea. The climate is coastal equatorial, which is characterized by two dry seasons (July to mid-August and November to March) and two rainy seasons (April to June and mid-August to October) (Ayamba et al., 2024). The average annual rainfall generally decreases with increasing distance from the coast, with an average annual rainfall of 2800 mm in Campo (MINFOF, 2014). Temperatures range between 24°C and 28°C along the coast. The CMTOU comprises unfenced land uses such as the Campo Ma’an National Park (CMNP, 264,000 ha), Forest Management Units (FMUs) for timber production, agro-industrial plantations, and a multipurpose Community Land area (CL) where farming, housing, infrastructures and activities for domestic purposes (hunting, fishing, artisanal logging of wood, and gathering) are permitted (African Wildlife Foundation, 2022; Djoko, Weladji, Granados, et al., 2022; MINFOF, 2014). Agriculture, primarily practiced as shifting cultivation using slash-and-burn methods, is the main local economic activity in the area (MINFOF, 2014). Staple food crops such as manioc, plantain, corn, yam, and cocoyam, are grown during the two annual rainy seasons, and farmers experience interactions with wildlife from the Park all year round (Eyebe et al., 2012). Study Area in the Campo Subdivision of the Campo-Ma’an Technical Operational Unit, Cameroon, Displaying the Forest Management Unit that was Allocated to an Agro-industry Currently Being Converted Into a Plantation, the Community Land, and the Camera Trap Stations (in Between which Reconnaissance Walks Were Conducted).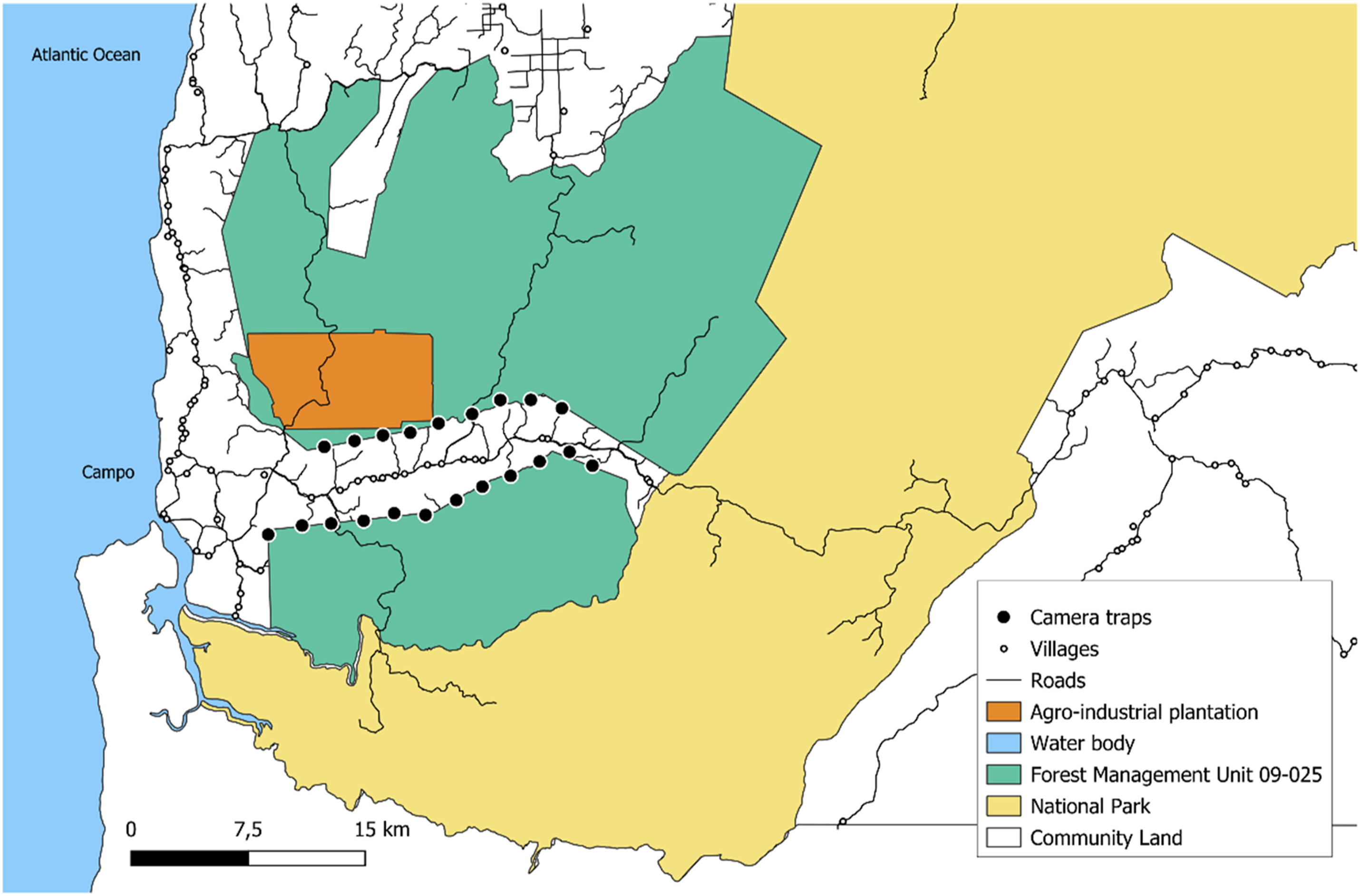
The Campo subdivision population is estimated at 7,000 habitants, reflecting an important increase over the past decade due to the growing demand for labor for large-scale projects in the CMTOU (Adolphe Edjabe, Election Cameroon manager for the Campo subdivision, personal communication, March 2025). However, it has a low population density compared to the rest of the CMTOU (MINAT, 2014). The CMTOU has been facing significant pressure from the development of large-scale projects such as the Kribi deep seaport, the Memve'élé hydroelectric dam, the agro-industrial development of HEVECAM (rubber company) and SOCAPALM (palm oil company), and mining exploration activities of SINOSTEEL (Engolo et al., 2024; Forje et al., 2021). Moreover, in 2019, the 60,000-ha protected area FMU 09-025 in the Campo subdivision was allocated to the agro-industry CAMVERT-SA for forest conversion into a palm oil plantation (Engolo et al., 2024; Forje et al., 2021; Mesmin et al., 2021). The latest inventory estimated the CMTOU forest elephant population at 243 individuals, representing a decline of 55% in under six years (Beukou-Choumbou et al., 2021). With the second largest number of elephants within the CMTOU found in the FMU 09-025, after Dipikar Island in the CMNP (Ayamba et al., 2024; Beukou-Choumbou et al., 2021), increasing reports of elephants visiting the small-scale farms in the Campo subdivision coincide with the establishment of the plantation, suggesting a drastic change in elephant habitat use which leads to increased human-elephant interactions (African Wildlife Foundation, 2022; Dhiaulhaq et al., 2024).
Data Collection
Camera Trapping
We deployed 21 Bushnell camera traps in the Campo subdivision of the CMTOU from May to October 2023. Camera trap stations were systematically placed at 2-km distances from one another along the border between the declassified FMU 09-025 and the Campo CL (Figure 1). If the grid point was deemed unsuitable to position the camera, a more appropriate area was chosen within 100 m of the initial point based on accessibility and wildlife or human activity potential. Camera trap stations were placed along trails created and maintained by wildlife or near fruiting trees, at a distance of 5 to 15 meters from the target feature (trail or tree). Because one camera trap did not work for the totality of the survey duration, we dropped data from the camera trap and reported data from the remaining 20 camera traps stations. Three cameras got stolen and did not get replaced, as survey ended shortly after. Camera stations were visited on average once every 35 days to replace batteries and SD cards. Cameras were set at 80-90 cm height, angled horizontally, and set facing north or south to avoid low sunlight interfering with the cameras. Small trees and high foliage were cleared from in front of the cameras to avoid image obstruction. Cameras were active 24 hours/day, and motion sensors were set to trigger immediately when movement is detected, with a delay of 1 second between consecutive photos while movement was still occurring. Photos were date and time stamped (see Figure 2). To account for bias arising from multiple triggers of the same individual or group by the same camera, photos at a site within a 30-minute interval were counted as a single independent detection event (Chakraborty et al., 2021; Collins & Weladji, 2024; Djoko, Weladji, Granados, et al., 2022; Li et al., 2012; Tanwar et al., 2021). Camera trap images were processed using Timelapse 2.0 v 2.3.0.8 (Greenberg, 2022). Sample Camera Trap Images of Forest Elephants From the Campo Subdivision, Campo-Ma’an Technical Operational Unit, Cameroon.
Reconnaissance Walks
Reconnaissance walks are often used in tropical rainforests, notably to monitor forest elephant populations (Blake et al., 2007; Fai et al., 2022; Gaugris et al., 2022; Hedges, 2012; Omoregie et al., 2020; Walsh & White, 1999; Whytock et al., 2021). As line transects are inefficient in thick forest vegetation, walks were conducted following a general compass bearing using a path of least resistance along hunting trails and natural features like watercourses (Hedges, 2012; Walsh & White, 1999). This method minimizes damage to vegetation and creation of new access routes for hunters and paths for wildlife. Recces were carried out between consecutive camera trap stations (2-km transect length) when stations were visited to replace batteries and SD cards. Trained team of two surveyors walked at a pace of approximately 1 km/h recording elephant signs (e.g., footprints, dung, trails, direct contact, vocalization and smell, feeding activity, CT damage, tree used for scratching) and anthropogenic signs (e.g., hunting and logging trails and camps, snares, direct contact, gunshot and chainsaw sounds, knife, empty cartridge shell, tree cut, stolen CT) (Djoko, Weladji, Granados, et al., 2022; Gaugris et al., 2022), but only fresh signs were analyzed as to be able to avoid recounting the same sign during subsequent sampling periods (Fragoso et al., 2016; Hedges, 2012). Same species’ signs found within 15 m of each other were treated as a single encounter (Gaugris et al., 2022). Each 2-km recce transect was surveyed within 16 hours, with interruptions for camera trap maintenance and for setting up camp at night when a suitable site was found.
Permits and Ethics
Authorizations required for this study were provided by the Cameroonian Ministry of Scientific Research and Innovation (MINRESI), Ministry of Forestry and Wildlife (MINFOF), and Concordia University. Chiefs from the villages we would be crossing during our survey were met beforehand, informed of our work, and shown the corresponding authorization letters. We also took the time to visit them each time we passed through their village. Following the ethical guidelines suggested by K. Sharma et al. (2020), community members were informed that their privacy would be respected, and no camera trap images would be published or shared with park staff for prosecution. Field assistants were local community members, contacted directly by our research team, and were paid for their work.
Data Analysis
Influence of Human Activity on Elephant Occurrence
We were interested in testing the effect of human activity on elephant occurrence using direct (camera traps) and indirect (recces) observations. We ran a separate model for each field method. To allow non-linear relations to be considered, we performed generalized additive mixed models (GAMMs) using the MGCV package version 1.9-1 (Wood, 2023) with the restricted maximum likelihood method. Models were fitted with a negative binomial distribution and logistic link function to account for the overdispersion in our data. Covariates were tested for multicollinearity using Variance Inflated Factor (VIF) using car package version 3.1-2 (Fox & Weisberg, 2019). For each model, the independent variable of human activity was log-transformed to provide a better fit model than untransformed data. The smoothing parameter was set to be k = 4 based on the expected degrees of freedom (Pedersen et al., 2019). Increasing k did not change the fit of the model.
Camera Trapping
Number of independent elephant detections per month at each camera station was used as the response variable, and human detection rate was used as a fixed effect (continuous variable). Human detection rate was determined as the number of independent human detections per month at a camera station, divided by the number of camera trap days multiplied by 100 (Collins & Weladji, 2024; Djoko, Weladji, Granados, et al., 2022; Li et al., 2012; Rovero & Marshall, 2009). The number of independent detections of a species and the corresponding detection rates can be interpreted as an index of species activity (Sollmann, 2018) and density (Rovero & Marshall, 2009) at a given site when subject to rigorous sampling procedures, and may be used to examine whether covariates explain variation in detection rates (Collins & Weladji, 2024; Sollmann, 2018). Camera trap station ID and month were modeled as random effects to account for spatial and temporal autocorrelation, and the log-transformed number of camera trap days per month at the station (a measure of the effort) was included as an offset term.
Reconnaissance Walks
Number of encountered elephant signs along recces per month within each camera zone was used as the response variable, and human sign density was used as a fixed effect (continuous variable). For easier comparison of results between camera traps and recces, the number of encountered human signs along recces was divided by each camera trap zone area (Ahrestani et al., 2018; Djoko, Weladji, Granados, et al., 2022; Walsh et al., 2001; Yackulic et al., 2011), i.e. 1-km2 centered on each camera station. Zone ID and month were modeled as random effects, and the log-transformed estimated linear distance traveled per month per zone (a measure of the effort) was included as an offset term. Due to hardware problems, we did not have access to the tracking data for each month, so we estimated the linear distance traveled by measuring the distance between all recorded signs using QGIS 3.32.3 (QGIS© software by QGIS Association). All types of activity signs were weighted equally in the analyses. To further compare the results obtained from the two survey methods, Spearman correlation coefficients were measured to assess the relationship between the number of detections and number of sign encounters for elephants and humans separately.
Activity Patterns of Elephants and Humans
We were interested in comparing the diel activity patterns of elephants and humans using our camera trap data. Following the procedure developed by Ridout and Linkie (2009), activity patterns were estimated nonparametrically using kernel density estimates (probability of a detection event occurring at any given time). Overlap coefficient between activity patterns, which ranges from 0 (no overlap) to 1 (identical activity patterns), was then determined. The estimator of the overlap coefficient (
Results
Our camera trap survey included 2164 camera trap days, with elephants being detected in 3001 photos for a total of 44 independent events, and humans in 217 photos for a total of 28 independent events (see Supplemental Table 1). Elephants were detected at 11 of 20 operational camera trap stations (55%), while humans were detected at 6 stations (30%). In all, around 335 kilometers were surveyed over 39 days for recent elephant and anthropogenic signs in recces, where 439 elephant signs and 102 human signs were encountered. Recent signs of elephants and humans were recorded in 20 of the 21 survey zones (95% each).
Influence of Human Activity on Elephant Occurrence
Camera Trapping
Human activity had no statistically significant effect on forest elephant occurrence (edf = 1.795, x2 = 4.873, p = 0.115), although the observed trend suggests a non-linear negative impact of human activity on elephant occurrence once detection rate exceeds a certain threshold (Figure 3). Relationship Between Forest Elephant Detection Count (Partial Residuals) and Human Detection Rate (Count per 100 CT Days). Model Estimates are Based on a Generalized Additive Mixed Model Regression Model. Shaded Area Corresponds to the 95% Confidence Interval.
Reconnaissance Walks
Human activity had a significant negative effect on forest elephant occurrence (edf = 1.00, x2 = 3.856, p < 0.05) (Figure 4). Relationship Between Forest Elephant Sign Encounter Count (Partial Residuals) and Human Sign Density (Count per km2). Model Estimates are Based on a Generalized Additive Mixed Model Regression Model. Shaded Area Corresponds to the 95% Confidence Interval.
Correlation Between Survey Methods
There was a positive Spearman correlation between elephant detection and sign encounter counts (ρ = 0.421, p < 0.001). There was however no correlation between the human detection and sign encounter counts (ρ = -0.108, p = 0.373).
Activity Patterns of Elephants and Humans
Elephants and humans had a low activity overlap ( Kernel Density Estimates of the Daily Activity Patterns of Forest Elephants and Humans. Shaded Area Under Both Curves Represents Overlap Between Activity Patterns. The Rug at the Bottom of the Plot Shows the Distribution of Independent Observation Events.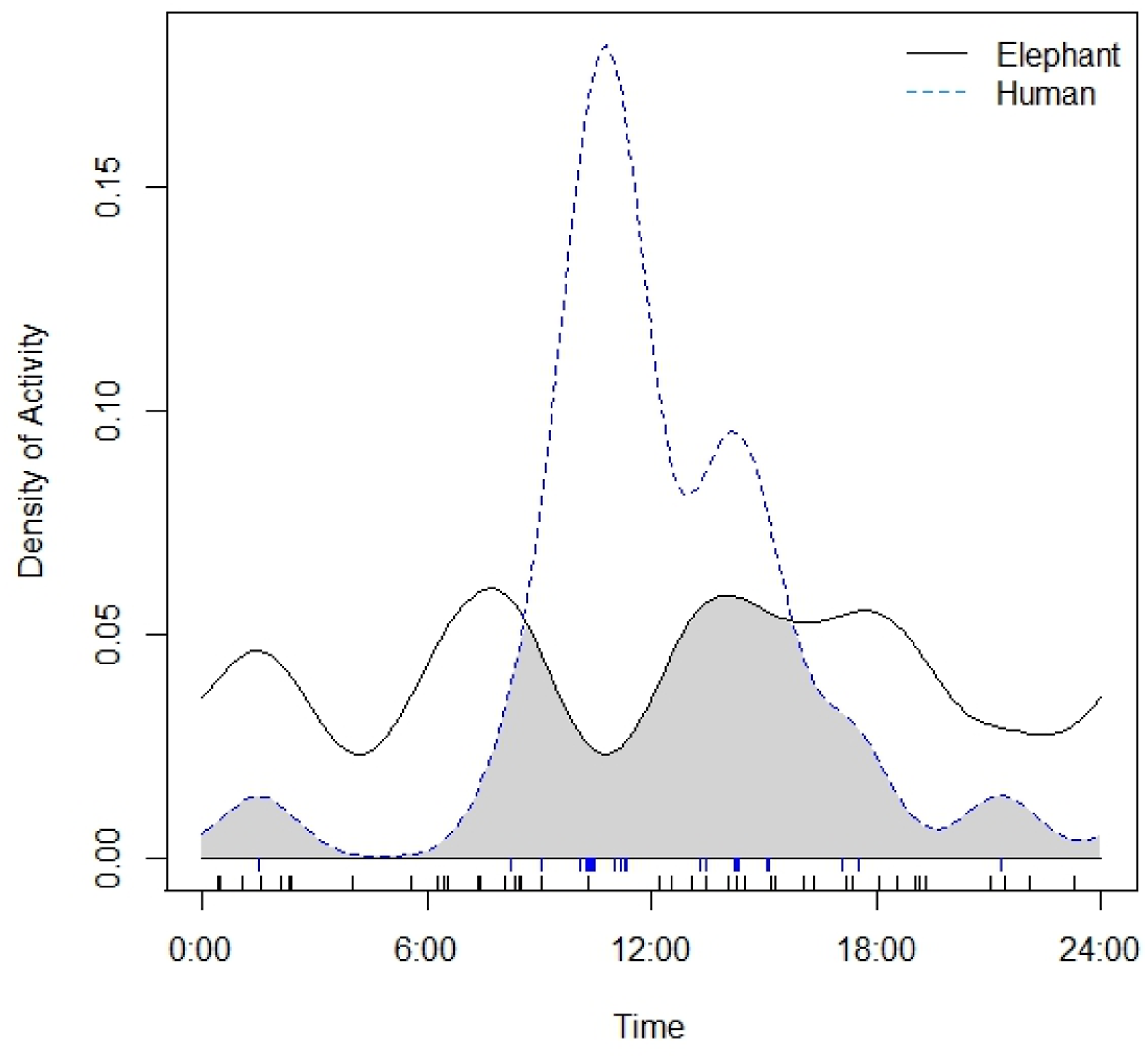
Discussion
Influence of Human Activity on Elephant Habitat Use
Our findings reflect the complexity of the forest elephant’s response to human activity in a human-dominated landscape, illustrating a multifaceted interaction between human-mediated risk and food resources. We found some evidence of reduced elephant occurrence as human activity increases from the two different survey methods, suggesting that human and forest elephants may spatially avoid each other to some extent. Our results align with findings in the CMTOU prior to the establishment of the agro-industrial plantation, where elephant relative abundance was negatively correlated with human activity such as hunting and logging (Djoko, Weladji, Granados, et al., 2022). However, the presence of elephants along the border between the Campo CL and the declassified FMU, hence within 2 to 2.5 km from the villages, suggests that elephants may be adopting a general risk-taking tactic by staying near human settlements, at least during the harvest period, which coincided with the timing of our study. This elephant presence is consistent with reports of elephants visiting villages and small-scale crop fields around the CMNP in recent years (African Wildlife Foundation, 2022; Dhiaulhaq et al., 2024; Mimeault, 2025; Urbain Nzitouo, Head of the Community and Participatory Management Service of MINFOF, personal communication, May 2023). It also reflects previous findings in the CMTOU, where the likelihood of elephant presence was higher in the CL than in the FMUs or CMNP, especially during harvest period when crops are mature and farmers report more frequent crop-raiding incidents (Djoko, Weladji, Granados, et al., 2022). This risky proximity to human settlements may be coupled with temporal avoidance, as our findings indicate distinct activity patterns between the two species. Elephants displayed reduced activity during the midday peak in human activity and were active at night while humans were asleep. This suggests that elephants may adjust their activity patterns to minimize encounters with humans. These results align with Davis et al. (2023) in Kasungu National Park, Malawi, who found that while savanna elephants did not avoid sites with higher human activity, they did avoid peak human activity times and exhibit nocturnal behavior when using roads. In areas associated with humans, elephants are indeed found to increase night-time activity (Adams et al., 2022; Gaynor et al., 2018; Hahn et al., 2022; Smit et al., 2023; Tiller et al., 2021; Wrege et al., 2010, 2024), as well as decrease crop raiding during the full moon to avoid higher visibility (Gunn et al., 2014), increase walking speed at night (Hahn et al., 2022) or when traversing agricultural areas (Songhurst et al., 2016; Tiller et al., 2021) and roads (Mills et al., 2018), and visit villages for shorter durations (Mbamy et al., 2024; Tiller et al., 2021). Our findings highlight the importance of considering both the spatial and temporal dimensions of forest elephant responses to human activity, as it enhances our understanding of the multifaceted strategies this species employs to mitigate risk in human-dominated landscapes.
Our results suggest a complex interaction between human-mediated risk and food resources, where elephants may be balancing the risks associated with proximity to human settlements against the benefits of accessing high-value food areas like crop fields and secondary forest. At the same time, they may employ temporal strategies to minimize direct interactions with humans. As crops can contain higher digestible energy than wild food plants (Branco et al., 2019), and clumped resources like crop fields provide a high nutritional payback with relatively lower search times (Blake & Inkamba-Nkulu, 2004; Branco et al., 2019), the nutritional benefits and easy access of crops may therefore outweigh the perceived risk of areas with higher human activity (Mills et al., 2018; Tiller et al., 2021), especially when crops are mature (Branco et al., 2019; Djoko, Weladji, & Paré, 2022; Matsika et al., 2024; Tiller et al., 2021). Elephant habitat selection and movement patterns can be influenced by seasonality (Djoko, Weladji, Granados, et al., 2022; Loarie et al., 2009; Mills et al., 2018), yet if the availability and quality of food in the elephant range declines, notably due to human encroachment, elephants may crop-raid throughout the whole year (Tiller et al., 2021). The observed proximity to human settlements could also be partially due to secondary forest near villages, which has been shown to attract wildlife, including forest elephants (Blake, 2002; Breuer & Ngama, 2021; Djoko, Weladji, Granados, et al., 2022; Poulsen et al., 2011), due to the higher abundance of young trees and secondary species (Poulsen et al., 2011).
Moreover, the presence and development of large-scale projects in the CMTOU implies a population increase in the area due to the demand for labor, which may, in turn, drive greater encroachment on wild areas (Billah et al., 2021; Forje et al., 2021), with humans and elephants competing for resources and space (Billah et al., 2021; Granados et al., 2012). This increased human activity and habitat loss in the CMTOU may overall heighten the perceived human-mediated risk level of the area, which could lead to a change in elephant habitat use through increased risk-taking behaviors to gain access to food. Tiller et al. (2021) reported a similar phenomenon in African savanna elephants, where spatial and temporal patterns of crop raiding by savanna elephants had changed seemingly in response to increased agriculture expansion and increased cattle grazing within protected areas in the previous 20 years. However, at fine-scale, human-mediated pressure might be low enough to allow elephants to occur near villages and allow for constant feeding opportunities with low risk (Djoko, Weladji, Granados, et al., 2022), especially as the population density outside key cities like Campo is low (Tchouto et al., 2006). The intensive maintenance and vigilance required by certain mitigation methods to deter elephants from fields at night, such as night-time guarding and chili fires, can create barriers to effective mitigation (Denninger Snyder & Rentsch, 2020; Parker et al., 2007). As such, none of the mitigation techniques implemented in the CMTOU were found to effectively reduce crop raiding by elephants and other wildlife (Djoko, Weladji, & Paré, 2022; Mimeault, 2025). Moreover, hunting regulations prohibit to hunt forest elephants (Ngandjui & Ringuet, 2010) and regular anti-poaching effort is conducted in the CMNP (African Wildlife Foundation, 2022), potentially lowering the perceived and real human-mediated risk in the area (Blake & Maisels, 2023). Yet, wildlife poaching offenses in the CMNP has increased drastically from 2008 to 2020 (Engolo et al., 2024), including some elephant poaching occurrences (Djoko, Weladji, & Paré, 2022).
Comparison Between Survey Methods
Recces proved more effective than camera traps in detecting both species in a greater number of sites and providing a greater amount of data overall. Although camera traps are valuable for detecting a large number of species (Wearn & Glover-Kapfer, 2019) and quickly and confidently identifying them (Thomas et al., 2020), sign encounter data can prove more efficient and effective for assessing species occurrence when number of sites where a species is detected matters (Fragoso et al., 2016, 2019; Gaugris et al., 2022). While the two survey methods produced consistent results for detecting elephants, elephants were detected in a greater number of zones using recces than camera traps, and encounter rates were higher than detection rates. Humans were also detected in a greater number of zones using recces than camera traps, although the two methods did not produce consistent results for detecting human activity. This discrepancy could be attributed to humans avoiding camera traps after first encountering them, especially since cameras may capture illegal activities, making people fearful of being recorded (Sandbrook et al., 2018). This explanation is further supported by the theft of three camera traps during our survey, although community members were informed that their privacy would be respected. Moreover, it was difficult to determine age of certain human signs, such as tree cutting, camps and empty cartridges. Discarding them from the analysis may have led to an underestimation of indirect signs of human activity. Our study highlights the importance of understanding the requirements of different survey methods to select the most appropriate ones based on research objectives, including focal species and environmental context. When possible, researchers still recommend the combination of different survey methods to maximize detection of all species and generate a more complete estimate of the target variables, especially in study sites with low species densities and activity (Fragoso et al., 2016; Moore et al., 2020; Nuñez et al., 2019; Plumptre, 2000).
Implications for Conservation
The spread of industrial logging has significantly transformed elephant habitats in Central Africa (Laporte et al., 2007), with Africa having the highest annual rate of net forest loss in 2010–2020 (FAO, 2020). However, in recent decades, oil palm plantations have increasingly become controversial due to their role in tropical deforestation and biodiversity loss (Oosterveer, 2015). This concern is particularly pronounced in Cameroon, which has the greatest amount of deforestation in Africa due to palm oil plantation expansion (Ordway et al., 2017). Accordingly, it is anticipated that palm oil plantations will soon surpass logging as a primary source of landscape disturbance in the CMTOU (Djoko, Weladji, & Paré, 2022). The subsequent population increase in the area due to the demand for labor may also increase demand for meat (Nlom, 2021), potentially contributing to rising poaching levels and threatening the survival of critical wildlife species (Engolo et al., 2024). As human disturbances in the CMTOU are increasing drastically as human population and agro-industrial activity increase, our findings highlight the possibility of habituation of elephants to human presence if the potential nutritional rewards outweigh the perceived dangers. That complex interaction between response to risk, food availability and quality, and forest elephant movement likely plays a significant role in one of the most critical issues threatening their conservation: crop raiding and increasing human-elephant conflicts (Blake & Maisels, 2023). Human-wildlife conflict fosters frustration among local communities and may justify retaliatory killings and negative perceptions towards conservation efforts (Dickman, 2010; Djoko, Weladji, & Paré, 2022). This could threaten the long-term survival of forest elephants, a critically endangered species. This further highlight the need to continue monitoring forest elephants in human-dominated landscapes, as assessing the impacts of habitat changes on their spatial and temporal patterns and strategy choice is essential for developing effective and up-to-date management and conservation strategies (Breuer et al., 2016; Djoko, Weladji, Granados, et al., 2022).
Furthermore, monitoring wildlife in densely forested habitats may be challenging due to limited visibility and the elusive behavior of many species (Zwerts et al., 2021), yet wildlife monitoring is essential for conservation research and evidence-based decision-making. In parallel, given the often limited resources available and insufficient financial capacity for monitoring biodiversity, particularly in tropical countries (Achieng et al., 2023; Gordon & Newton, 2006), it is essential to use the most efficient survey methods. Maximizing data collection while minimizing costs and time can make research more accessible and ultimately support better decision-making in wildlife management and habitat preservation.
Supplemental Material
Supplemental Material - Forest Elephants in a Human-Dominated Landscape: are They Risk-Takers?
Supplemental Material for Forest Elephants in a Human-Dominated Landscape: are They Risk-Takers? by Lea Mimeault and Robert B. Weladji in Tropical Conservation Science
Footnotes
Acknowledgements
We thank Patrick Paré and Mélissa Loiseau of Zoo de Granby for their support and helping with data collection, respectively. We are grateful to the Ministry of Forestry and Wildlife (MINFOF) of Cameroon for facilitating the research to take place in the CMTOU. The authors appreciate the Zoo de Granby, Quebec Centre for Biodiversity Sciences, and Concordia University for the financial support. Finally, we thank the field assistants Louis Desiré Dontégo Kafack, Chatelin Ndongo, Théo Tokari Lando, Bienvenue Mvogo, Dieudonné Assembe, Alexandro Awouma, Yann Menye, Lovy Engon, Moussa Roméo Tokari Lando, Claude Prévot Eyi Mengue, Serge Iya Ngongo, and Jean Paul Loïc Menye.
Statements and Declarations
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by the Zoo de Granby, Quebec Centre for Biodiversity Sciences (QCBS), and Concordia University.
Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.