
Abstract
Introduction
Most apex predators are facing dramatic population declines and range reductions (Ripple et al., 2014; Treves and Bruskotter, 2014). Understanding how these species move and use space is important to develop and implement effective conservation plans. These species typically require large home ranges for their survival, and they occur at low population densities (Carbone and Gittleman, 2002; Woodroffe and Ginsberg, 1998). Diverse factors determine how much space an individual requires, such as body size, prey availability, habitat suitability, and sociability (Gittleman, 1989; Sunquist and Sunquist, 1989). Further, landscape configuration and connectivity are also critical aspects that determine the movement patterns required to access suitable habitat (de la Torre et al., 2017a). However, very little research has been conducted on how the transformation of natural ecosystems by human activities can affect the use of space by large carnivores and their survival, especially in the tropics.
The jaguar is the largest cat in the Neotropics and one of the most iconic species in the Americas. Although jaguars are not endangered at the same level as some other large carnivores, recent evaluations find that most populations are threatened (de la Torre et al., 2018), and many have gone regionally extinct (Galetti et al., 2013; Paviolo et al., 2016; Quiroga et al., 2014). Globally, the jaguar has been extirpated from 55% of its historical range and is considered as Near Threatened by the IUCN Red List (Quigley et al., 2017), but in Panama, it is classified as endangered due to its national status (Ministerio de Ambiente, 2016). The long-term viability of jaguar populations is mostly affected by habitat destruction, retaliatory killing, illegal hunting, and prey depletion (Moreno, Bustamante, et al., 2016; Moreno et al., 2015; Moreno et al., 2024; Quigley et al., 2017; Sanderson et al., 2002). Habitat destruction is caused by the expansion of livestock ranching, agriculture, and human settlements (Moreno, Bustamante, et al., 2016; Moreno, Valdés et al., 2016b). Mortality from retaliatory killings in response to livestock predation is particularly harmful to their populations, and this is now exacerbated by increased pressure from poachers for China’s illegal wildlife trade (Morcatty et al., 2020; Moreno et al., 2015, 2024; Moreno & Olmos, 2008).
Spatial ecology studies have illustrated that jaguars require extensive areas to meet their spatial requirements. For instance, home range size estimated using GPS telemetry varies from 13 to 1,154 km2 for females and from 35 to 2,914 km2 for male jaguars (Thompson et al., 2021). In tropical wetland ecosystems, such as the Pantanal in Brazil, jaguars had smaller home ranges than in humid and dry tropical forests, presumably due to differences in prey availability. Female home ranges are generally smaller and determined by the abundance of prey, which is critical to meeting the energetic requirements of reproduction (Cavalcanti and Gese, 2009; McBride Jr. and Thompson, 2019). Males have larger home ranges to overlap multiple females and defend a territory against other males (Cavalcanti and Gese, 2009). Jaguars are solitary most of the time except for periods of mating, females with cubs, although some tolerance among conspecifics is allowed (Cavalcanti and Gese, 2009; Kanda et al., 2019). Jaguars are typically nocturnal or crepuscular and reduce movements during the day (Cavalcanti and Gese, 2009; de la Torre et al., 2017b; Kanda et al., 2019), with males moving more than females (McBride Jr. and Thompson, 2019; Morato et al., 2016; Thompson et al., 2021).
Across their range, jaguars live in a variety of habitat types, and their selection for a specific resource, such as habitat type, water source, and prey changes is a function of its availability in the landscape (Morato et al., 2018). For instance, studies of jaguar movements in savannas and natural grasslands, such as in the Brazilian Pantanal, found they select forests and water sources for nocturnal activities and dry grasslands when resting (Kanda et al., 2019). While on the move and hunting, jaguars in the Pantanal choose areas with dense cover and nearby available water bodies in the landscape (Cavalcanti and Gese, 2010; Gese et al., 2018; Kanda et al., 2019; Morato et al., 2018). This suggests that the jaguar’s behavioral flexibility allows them to adapt their movements according to local conditions, while also increasing the potential for conflicts with humans and jaguar mortality, especially in areas with high livestock density (Morato et al., 2018). Further, previous studies using GPS telemetry have documented that jaguars in tropical rainforests selected areas covered with primary and secondary forests and avoided areas close to human activities such as roads and towns (Colchero et al., 2011; Conde et al., 2010; de la Torre et al., 2017a; Morato et al., 2018). However, scant information about the indirect impact of tropical rainforest transformation into cattle pastures or agriculture on jaguar spatial ecology is available (Bejarano Alegre et al., 2024). This information is crucial to identify habitat quality requirements, and areas of potential conflict, and plan potential corridors and protected areas.
This study aims to describe the spatial ecology of jaguars in a forested landscape partially transformed by human activities in Darien, Panama. Using GPS telemetry, we documented home range size, movement patterns, interactions between individuals, and habitat use. This is the first study that presents data on jaguars in a forested landscape transformed into cattle pastures using GPS telemetry, and the results presented illustrate how the species can persist in this kind of environment, which has not been documented previously. We were interested in knowing the home range characteristics and habitat use of jaguars in an area with cattle pastures combined with tropical rainforest remnants, to describe the spatial organization and how different individuals share the space. We predicted that jaguars would use the forest remnants to traverse the cattle pasture areas, as they prefer to move across areas with forest cover. We predicted that jaguars with overlapping home ranges exhibit temporal segregation in their spatial usage and would avoid utilizing the same sites simultaneously.
Methods
Study Area
Our research was conducted in the middle section of the Chucunaque River basin, in the Darién Province on the Pacific slope of Panamá. The region shares its northern border with the Guna Yala indigenous territory, the eastern border with the Tuira River and the Darién National Park, and the western border with the Sabanas River basin (Figure 1). The average elevation throughout the basin is approximately 90 meters above sea level, where the highest point is Cerro Armila, which is located to the southeast of the basin with a maximum elevation of 1,421 m.a.s.l. Throughout the basin, there is an average annual rainfall of 2,300 mm. Most of the basin was originally covered by tropical rainforest, however, this has been severely affected by anthropogenic activities. Human population density in this region ranges from 1.7 to 10.4 persons per square kilometer (CIESIN - Columbia University, 2018). Our study area includes about one hundred towns, most of which are located along the Pan-American highway, with the largest being Meteti, Yaviza, and Agua Fría. The remaining towns are situated within the territories occupied by the Emberá-Wounaan (Comarca de CEMACO) and Guna de Wargandi communities along the riverbanks. (A) Location of Our Study Area in Darien Panama; (B) Land Use in Our Study Area and Home Ranges of Jaguars Estimated Using the Autocorrelated Kernel Density Estimation (AKDE).
We studied jaguars in the “Agua Buena” area, 10 km north of the town of Meteti. Our study area encompasses ∼ 2,500 km2, where some of the forest in this region has been converted into pastures for cattle and forest plantations (Figure 1). Based on the land use classification for our study area (Correa, 2023), we found that most of the region is still covered by primary and secondary forests (65.4%). Other land use categories include pastures for cattle (21.0%), forest plantations (6.5%), regrowth areas (2.0%), various types of crops (1.6%), and urban and infrastructure areas (0.4%). The majority of the ranches in this area belong to immigrants who arrived in Darien from western Panama, called “Colonos” (settlers). They arrived in search of virgin lands to cultivate, leading to a broad and disorganized process of human colonization. We have been working in the Agua Buena area since 2018 to study jaguars on farms and implement measures to mitigate human-jaguar conflict.
Data Collection
Our capture campaigns were implemented specifically at Finca Los Lagos, which has 101 hectares that include a matrix of cattle pastures and tropical rainforest. We captured jaguars using foot snares, and we placed a VHF radio transmitter at each foot-snares to monitor if the traps were triggered (de Araujo et al., 2021). The traps were checked every 2 hours during the day and night. When an animal was captured, we used a dart fired from a CO2 pistol or rifle to immobilize it. The dart contained a combination of Butorphanol (0.4 mg/kg), Azaperone (0.15 mg/kg), Medetomidine (0.08 mg/kg), Ketamine (2 mg/kg) and to reverse the dose it was used Atipamezole (4:1) to reverse the Medetomidine and Naltrexone (3:1) to reverse the Butorphanol. All captured animals were carefully supervised by a team of veterinarians. During immobilization, we assessed individual body condition and estimated age based on coat color, tooth wear (Stander, 1997), and gum-line recession (Laundré et al., 2000). We recorded body mass, linear measurements, and weight. Animals captured were deployed with a satellite global positioning system (GPS) collar Telonics® Gen4 GPS-Iridium, model TGW-4577-4. We programmed GPS collars to acquire a location every hour (24 locations/day), and to send data through to be archived at Movebank (Kays et al., 2022).
Data Analyses
Home Range Size and Movement Rates
We used the Autocorrelated Kernel Density Estimator (AKDE) as our measure for home range, using the ctmm package (Calabrese et al., 2016) implemented in MoveApps (Kölzsch et al., 2022; Sciaini and Signer, 2023). We first evaluated semi-variograms to confirm the animals were range-resident, then selected the best movement model based on AIC and calculated the 95% ADKE. We chose the AKDE method for evaluating home ranges because it addresses potential underestimation issues and considers autocorrelation (Calabrese et al., 2016; Morato et al., 2016; Noonan et al., 2019). However, it is important to note that AKDE is an extrapolation method and may overestimate home range areas. We calculated the speed between locations at different times of the day using MoveApps to determine the movement rate and activity periods of the animals tracked (Scharf, 2023).
Interactions
We evaluated spatial and dynamic interactions among the jaguars tracked simultaneously. We evaluated the AKDE estimates with bias-corrected Bhattacharyya coefficient (BC) to quantify the spatial overlap (Winner et al., 2018). The BC is a product based on two utilization distributions and measures the affinity between two individuals, and it assumes that individuals use space independently of one another. Index values in BC range from 0 (for two home ranges with no overlap) to 1 (for two home ranges with the same utilization distribution). We used the function “overlap” in the “ctmm” package for R 4.1.1 to estimate the BC (Calabrese et al., 2016). To evaluate whether the animals tracked interacted actively during their simultaneous tracking, we used the proximity analysis (Prox) (Bertrand et al., 1996; Long et al., 2014). The proximity analysis determines the proportion of simultaneous fixes (ST), which are proximal fixes based on a given distance threshold (dc). The Prox index is calculated as: Prox = ST/dc. We defined the distance threshold between two simultaneous fixes as 1000 m and we defined the temporal threshold between simultaneous fixes as 24 hours. We chose these wide thresholds to provide distance and time intervals that were sufficiently large to determine conservatively that the two animals tracked were using the same space simultaneously. We only included in this analysis the simultaneous fixes between the tracked animals that were obtained during the same time slot as defined previously. We used the package “wildlifeDI” (Long et al., 2014) for R 4.1.1 to perform the proximity analysis.
Step Selection Functions
List of Landscape Covariates Used to Evaluate jaguars’ habitat use in Darien, Panama.

We resampled the data to constant 2 ± 0.1 hours intervals. We simulated 19 “available” steps for each “used” step; since our GPS telemetry dataset has many locations per individual, and a low ratio of simulated to used steps it is sufficient for parameter estimation (Thurfjell et al., 2014). Step lengths were drawn from the empirical movement data using a Gamma distribution with rate and shape parameters estimated from the empirical data of step length distribution of the tracked jaguars. Turning angles were also drawn from the empirical data for the collared jaguars using a von Mises distribution. We used the “amt” package (Signer et al., 2019) for R version 4.0.2 (R Core Team, 2016) to generate the random steps.
For each used and available step, we calculated the values of the landscape covariates at the endpoint of the steps. All explanatory landscape covariates were rescaled by subtracting the mean from each variable value and then dividing them by the standard deviation. We constructed several SSF models using the “amt” package (Signer et al., 2019). We built several models with different combinations of the covariates and then used the Akaike Information Criterion (AIC) to identify the best SSF (Burnham and Anderson, 2002), and these analyses were implemented using the MuMIn R package (Bartoń, 2019). We selected the best-fitting models using AIC with a difference of ΔAIC < 2, calculated the model average coefficients for the best candidate models (Burnham and Anderson, 2002), and estimated the importance of predictor variables by the Sum of Weights (SW = 1; Galipaud et al., 2014). These analyses were implemented using the MuMIn R package (Bartoń, 2019). We tested all explanatory variables for multicollinearity using Pearson’s correlation matrix, and we did not include in the same candidate model variables that were correlated at |r|> 0.5.). For example, the covariates percent forest, percent grassland, and distance to forest were not included in the same models as they were highly correlated.
Results
Home Range Size and Movement Rates
Tracking Effort and Descriptive Movement Metrics for the Five jaguars in the Darien of Panama.

Results From the Home Range Model Showing the Size of the 95% AKDE (and Confidence Intervals), Home Range Crossing Time (tau Position), and Effective Sample Size (Degrees of Freedom).

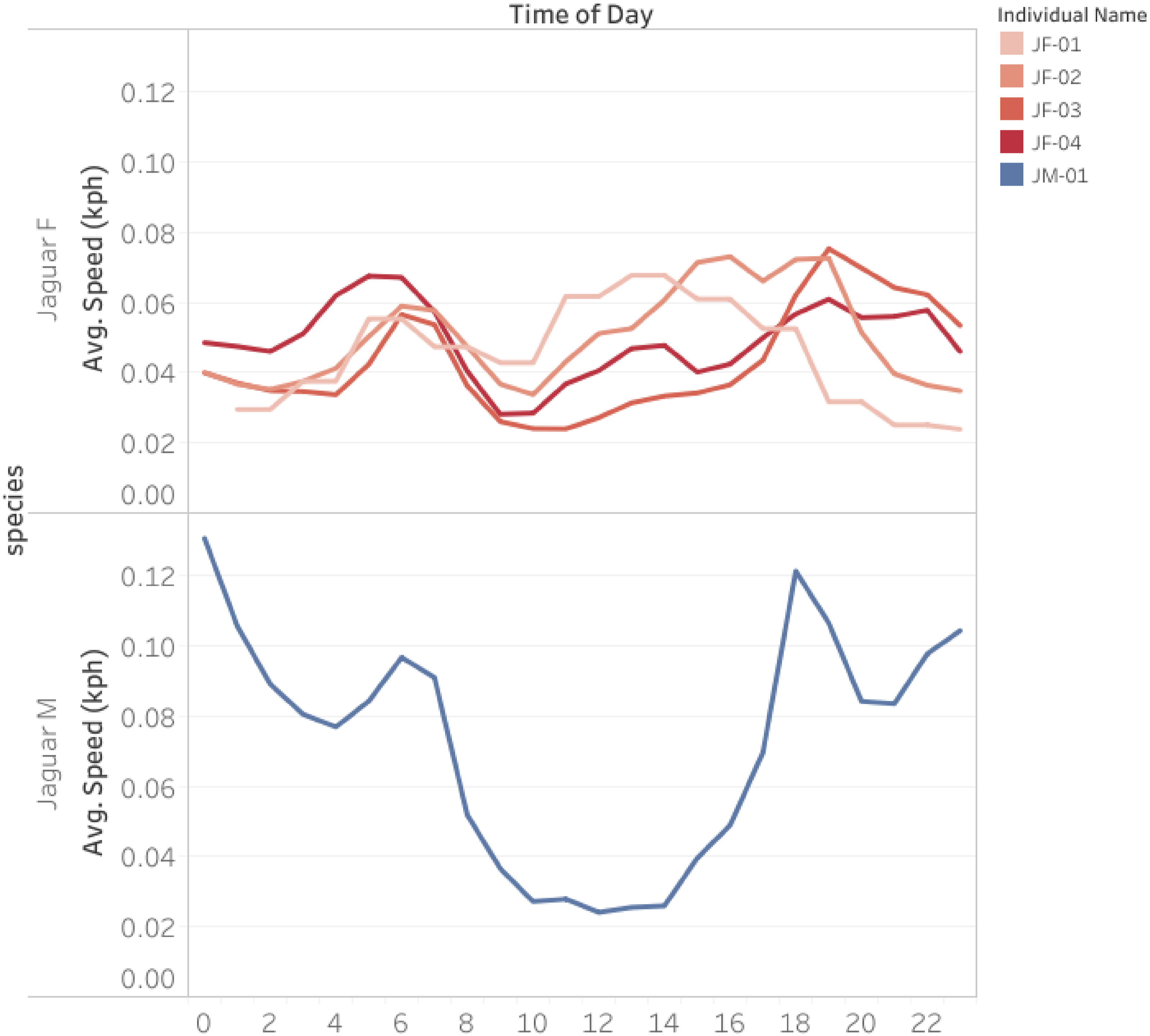
Average Movement Speed Across the Day for the Female Jaguars (Red), and Male Jaguar (Blue). The Graph Shows a Running Average of 2 Hours.
Interactions
Percentage of AKDE Overlap and Results of the Proximity Analysis (Prox) for Jaguars Tracked With Overlapping Home Ranges.


Pairs of Individuals Tracked Simultaneously Showing the Distance Between Simultaneous Fixes and the Period That Were Tracked Simultaneously. Graphs Show That These Individuals Stayed Close to Each Other at the Same Time During the Tracking.
Habitat Use and Selection
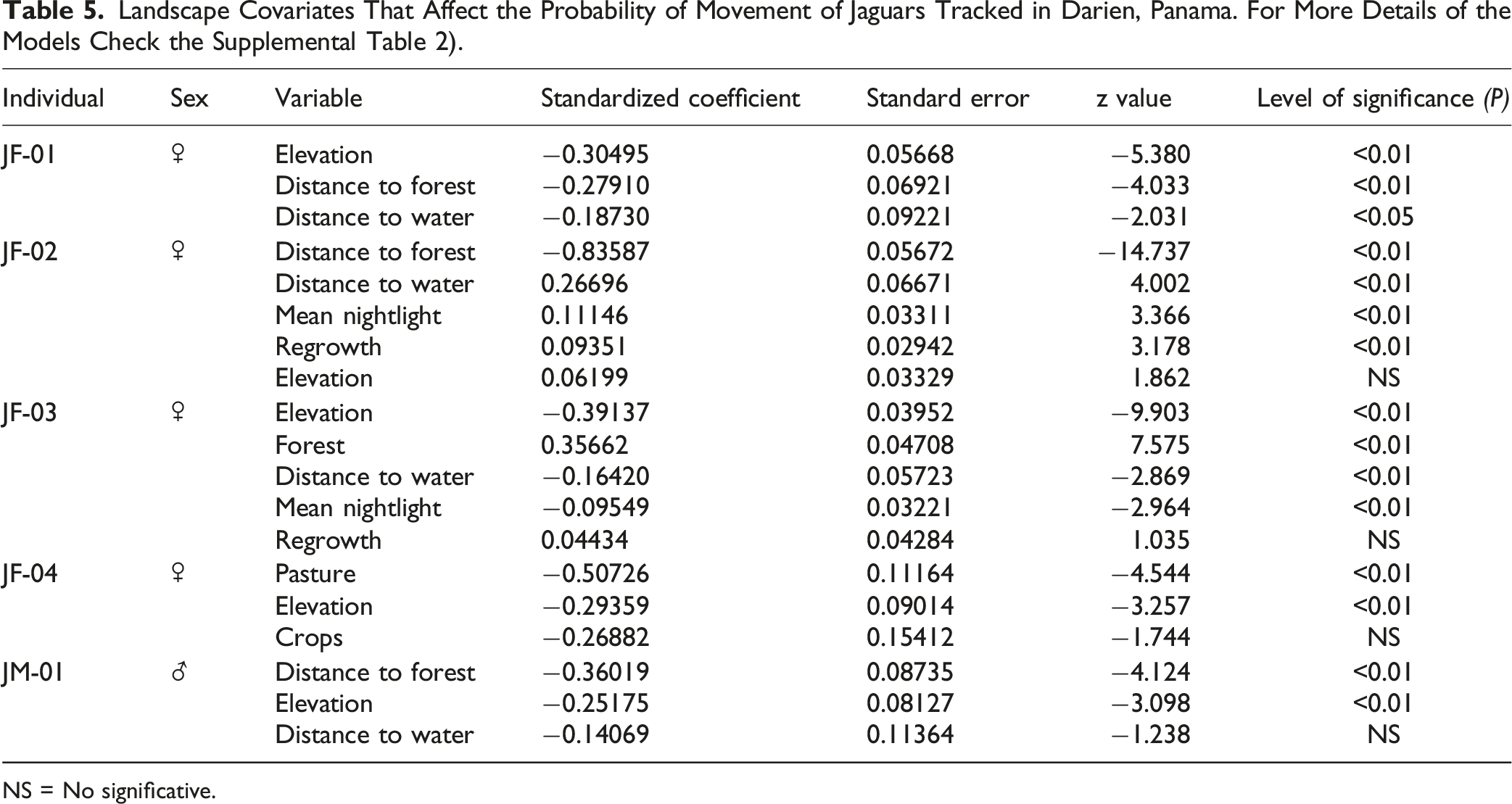
Jaguars spent most of their time in natural forested habitats (native forest, forest plantations, and regrowth), followed by agricultural areas (grass pasture, crops), and wet areas (Figure 4 and Figure S2, Table 5, Supplemental Tables 1 and 2). The step selection functions revealed some similarities between the jaguars tracked in the transformed areas of the Darien. All tracked animals selected forested areas or those that are near the forest while avoiding cattle pastures. This selection was indicated by the positive effect of the percentage of forest (Forest) and the negative effect of the distance to forests (Distance to forest) covariates (Figure 4, Table 5). This means that jaguars selected forested areas, as well as low values of percentage of pastures. However, we excluded these three covariates from the same SSF models due to their high correlation (Supplemental Table 2). Instead, we created a separate set of models that included the covariates of percentage of forest, percentage of pastures, and distance to forest (Supplemental Table 2). Further, two female jaguars also selected to move across areas of shrubland and secondary forests, where the vegetation is recovering, as indicates the positive effect of the regrowth covariate (Figure 4, Table 5). Further, four out of the five jaguars tracked actively moved near the main river of the region and across lowland areas, as indicated by the negative effect of the distance to water and elevation covariates. Only one female jaguar with an overlapping home range with the other jaguars avoided areas close to the main river and selected areas with higher elevations (JF-02) (Figure 4, Table 5). Jaguars had a complex response to the areas that showed more anthropogenic activities, with one female jaguar being attracted to areas with higher values of mean nightlights (JF-02), and another one avoiding these areas (JF-03 – Figure 4, Table 5). Significant Coefficients for Step Selection Functions of Five Jaguars Tracked in This Study. Positive Values Suggest They Use a Habitat Type More Than You Would Expect Based on Its Availability, Except for the ‘Distance to’ Covariates, for Which Negative Values Indicate They Select for Locations Closer (Shorter Distances) to That Feature. Landscape Covariates That Affect the Probability of Movement of Jaguars Tracked in Darien, Panama. For More Details of the Models Check the Supplemental Table 2). NS = No significative.
Discussion
We found important differences in the space use of jaguars in the transformed areas of Darien Panama compared to existing studies. Overall home range sizes of jaguars were smaller than those reported for other areas with tropical rainforests in Mesoamerica, such as southern Mexico and Costa Rica. We also find important differences in the average movement speed across the day for jaguars in comparison with results reported for other areas, with higher movement rates during midday and midnight for females. Our results also indicate that jaguars in this transformed landscape exhibited extensive home range overlap with their conspecifics, though the proximity analysis indicated they generally avoided occupying the same sites simultaneously. SSF illustrated how the animals tracked tend to move in the transformed landscapes in the Mesoamerican region. Overall, all animals tracked tend to move across forested areas, shrubland, and secondary forests or those that are near the forest, while avoiding cattle pastures. Further, most of the jaguars tracked actively moved near the main river of the region and across lowland areas. Animals tracked exhibited a complex response to anthropogenic activities, with some individuals being attracted to areas with higher human activity and others avoiding these areas.
According to previous research using GPS telemetry, jaguars have been found to have extensive home ranges (48 – 633 km2) in regions with tropical rainforests in Mesoamerican countries such as Mexico and Costa Rica (Cruz et al., 2021; de la Torre et al., 2017b; Montalvo et al., 2020; Thompson et al., 2021). Generally, the size of jaguars’ home ranges decreases with increasing habitat productivity and forest cover and increases with higher human perturbation (Thompson et al., 2021). Thus, we would have predicted that jaguars would have larger home ranges in these disturbed cattle pastures in Darien, Panama. It could be that the high productivity of the Darien rainforest allows jaguars to have small ranges, or that their predation on cattle from the ranches facilitates smaller ranges.
During our study, we found that all the tracked jaguars had overlapping home ranges, but they avoided using the same areas at the same time. These results indicate that while there is extensive overlap between male and female jaguars in our study area, there is also some temporal segregation, suggesting that there is territoriality and temporal separation between individuals. Other studies focusing on the spatial ecology of the species have indicated that jaguars maintain exclusive core areas within their activity areas. Solitary felids show spatial segregation, which implies territoriality. Exclusive areas are typically linked to high resource concentration, and territorial behavior affects the interactions between individuals (Azevedo and Murray, 2007; Scognamillo et al., 2003). However, it’s important to note that other individuals not tracked in our study might have occupied the area at the same time without our knowledge.
Though the male and one female jaguar showed the typical predator activity pattern of being less active during the day and more around dusk and dawn, the three other female jaguars moved more during the day. In an area with a high concentration of jaguars and significant spatial overlap between their home range areas, differences in activity patterns could facilitate the coexistence of these individuals by avoiding simultaneous activity. However, these variations in activity patterns could also be influenced by the different activity patterns of their prey (Foster et al., 2013; Harmsen et al., 2011; Karanth and Sunquist, 2000). The three female jaguars with predominantly diurnal activity included a greater proportion of pasture in their home range areas, indicating that they were more active during the day possibly due to a significant presence of cattle in their diet. Our observations also documented that at least two of these jaguars preyed on livestock on the farms they frequented (Moreno unpublished data), suggesting that their daytime activity may be driven by their predominant search for domestic livestock as their main prey. This behavior may be due to the inadequate management and surveillance on many of these farms, as they are often isolated or have minimal human presence, leaving the livestock unattended.
Our data showed that jaguars in working areas of the Darien prefer to move through forested areas or those near the forest while avoiding cattle pastures. This has significant implications for the conservation plans for the species at the landscape scale. In fragmented environments, these animals select to travel through primary forests or areas with good forest cover. These results suggest that jaguars can adapt to some degree of forest transformation due to human activities in tropical regions (Bejarano Alegre et al., 2024). However, they still prefer to move through forested areas or areas with cover or sites close to the forest. In general, jaguars in forested areas always prefer sites with good forest cover since these places provide prey availability, forest cover for stalking prey, and refuge for rearing cubs (Bejarano Alegre et al., 2024; Colchero et al., 2011; Conde et al., 2010; Crawshaw Jr. and Quigley, 1991; Rabinowitz and Nottingham, 1986). The distance to water covariate was also significant for most jaguars, with a positive coefficient, indicating that tracked jaguars prefer sites close to rivers. Jaguars are known to use riparian habitats to move through landscapes and use sites with permanent water more frequently (Crawshaw Jr. et al., 1991; Schaller and Crawshaw, 1980). Elevation and slope covariates were also included in the best habitat model for several of the individuals tracked and had a negative coefficient for most of the jaguars, indicating that jaguars infrequently used higher elevation sites (de la Torre et al., 2017a). Jaguars have been associated frequently with lowland areas, and these results suggest that jaguars in the Agua Buena Darien region avoided sites at higher elevations. Only one female jaguar (JF-02) avoided areas near the main river (Río Chucunaque) and used the higher areas elevations within its home range. This female jaguar extensively overlapped her home range area with two other female jaguars and potentially using sites with higher elevation and away from the main river allowed her to also be spatially segregated from the two other female jaguars that occurred in the area.
We acknowledge that our results may be biased due to the small number of individuals we tracked. This sample size is likely not representative of the overall population of jaguars in our study area. Specifically, we tracked only one male jaguar whose behavior may be atypical. Additionally, the five animals we monitored were observed in a relatively small region, while we are drawing conclusions about their habitat use on a much larger scale. However, our data provides valuable insights for the first time on jaguars in areas that were once tropical rainforests but have since been deforested for cattle pastures and crops. This shows how these big cats can thrive in these landscapes despite human activities.
Implications for Conservation and Management
The results of this study suggest that jaguars can persist in areas transformed for livestock farming in the tropical forests of Mesoamerica. Even these areas can also be important breeding sites for jaguars in the region. Since the study began, we have documented that all female jaguars monitored with satellite collars (JF-01, JF-02, JF-03, and JF-04) in the study area have had cubs in the farm areas. However, for them to remain in these areas, it is necessary to maintain remnants of the forest through which they can move and take refuge (Bejarano Alegre et al., 2024), to have a base of natural prey within these areas, and also for the people who live in these areas to tolerate jaguars in their ranches. Understanding the movements of jaguars in these areas helps to dimension the scale at which we should work to resolve conflicts between jaguars and people (jaguar predation on domestic animals) and to develop mechanisms that allow the local people to improve the coexistence with the species (de la Torre et al., 2021; Moreno, Valdés, et al., 2016, 2024; Moreno & Olmos, 2008). In this study, we evaluate the movement of jaguars in a transformed landscape in Darien Province Panama, and this information should serve as a guide to the conservation planning of jaguars and their habitat in the region. Agencies such as MiAmbiente Panama should incorporate this information into their landscape planning for the region, to ensure the long-term conservation of jaguar habitat and corridors. It is essential to work with local ranchers to reduce the risk of jaguar predation on domestic cattle to ensure the persistence of jaguars in these areas (Moreno et al., 2015, 2016b).
Supplemental Material
Supplemental Material - Jaguar Space Use in the Working Landscape of Darien, Panama
Supplemental Material for Jaguar Space Use in the Working Landscape of Darien, Panama by Ricardo Moreno, J. Antonio de la Torre, Josué Ortega, Natalia Young, Arturo Puertes, and Roland Kays in Tropical Conservation Science
Footnotes
Acknowledgments
We extend our gratitude to the following organizations and individuals for their funding and support: the National Secretariat for Science, Technology and Innovation of Panama (SENACYT FID 22-015), the Ministry of the Environment of Panama, the U.S. Fish and Wildlife Service (USFWS), the National Geographic Society, the UNDP Small Grants Program with GEF Fund, The GEF 7 Jaguar project of Panama with MiAmbiente/Yaguará Panama Foundation/PNUMA, Project Survival Cat Heaven, Pedro and Pituka Heilbron, Louisa and Mike Weinrib and the Bailey Wildlife Foundation. We would also like to thank the people who supported the jaguar capture campaigns, including Wildlife Pharmaceuticals Mexico veterinarians Ivonne Cassaigne and Susana Ilescas. Additionally, we express our appreciation to Leonardo Prettelt, Jack Gewart, Mariana Parks, Roberto Martinez, Bernardo Castillo, Ricardo Jimenez, Dora Dutary, Sharon Abramovich, Valentina Ellis and Eloisa Astorga. Special thanks to Dr. Claudia Rengifo and the students of the Faculty of Veterinary Medicine at the University of Panama, as well as the biology students at the University of Panama and those at UNACHI in Chiriquí for their support during the monitoring of the tracked animal. We would also like to recognize the De Leon family from the Los Lagos ranch for their unwavering support. Furthermore, we express gratitude to Milton Solano, Edgar Sanchez, Benjamin Walker, Elliot Brown, John Cleghorn, Ilina Cisneros, Alejandra Florez, Guillermo McPherson, Ana Dutary, Francisco Samuel Alvarez, Vayron De Gracia, Jean De Leon, Pablo Guerra, Annie Lopez, Yarisbeth Alain, Hilda Moran and Rafael Avilez from ISAE University in Darien and the community of Agua Buena and Bella Vista.
Statements and Declarations
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Bailey Wildlife Foundation, The GEF 7 Jaguar project of Panama with MiAmbiente/Yaguará Panama Foundation/PNUMA, Secretaría Nacional de Ciencia, Tecnología e Innovación, UNDP Small Grants Program with GEF Fund, U.S. Fish and Wildlife Service, National Geographic Society, Ministry of the Environment of Panama, Project Survival Cat Heaven.
Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.