
Abstract
Keywords
Introduction
Forest-savanna ecotones are characterised by a mixing of tree functional types with forest species co-occurring with forest-savanna transitional species (henceforth, “transition species”) and savanna species (Ametsitsi et al., 2020; Armani et al., 2018; Schrodt et al., 2015; Swaine, 1992; Torello-Raventos et al., 2013). Predicting ecotone movement, which occurs occasionally in response to global environmental change factors, such as climate change, requires insights into changes in environmental factors that influence tree recruitment as well as into responses of seedlings of the co-occurring tree functional types (TFTs) to changes in environmental factors.
Fire differentially affects tree species recruitment in forest-savanna ecotones due to differential tolerance among co-occurring tree functional types within the landscape (Issifu et al., 2019, 2020). Fire has a greater effect on forest species and possibly explains the generally lower abundance of seedlings of forest species, compared to savanna and transition tree species in forest-savanna ecotones (Armani et al., 2018; Azihou et al., 2013; Issifu et al., 2019). In the forest-savanna ecotone, however, we find dry forests with specific species composition (referring to species in drier forest types including those regarded as transition species) that can establish under prevailing fire regimes under which tree species from moister forest types fail to establish (Cardoso et al., 2016; Swaine, 1992).
In addition to fire, dry season drought constitutes an important constraint to tree seedling establishment success (Engelbrecht et al., 2007; Hoffmann et al., 2004). Soil moisture may vary substantially in the dry season even between adjacent forest and savannas via influences of microclimate and soil properties (Ametsitsi et al., 2020; Issifu et al., 2019). Due to differences in drought tolerance among tree species, effects of dry season drought on tree recruitment differ among tree functional types (Hoffmann et al., 2004). Thus, tree species which originate from forest environments may be less tolerant for both fires and prolonged drought periods relative to species from drier forest types.
Differences in responses of the different tree functional types to fire and/or dry season drought are linked to differences in carbohydrate storage in roots. This has been empirically tested (and supported) for forest and savanna tree functional types (Hoffmann et al., 2004; Issifu et al., 2019; Tomlinson et al., 2012; Boonman et al., 2020). The amount of stored carbohydrates in roots may be dependent on plant size, although size-dependent mortality may differ between tree functional types (Gignoux et al., 2009), suggesting that not all species and functional types increase stored carbohydrates in roots with increasing plant size. Some species may invest in higher root carbohydrate storage much earlier resulting in higher survival at smaller sizes than others (Gignoux et al., 2016). This may be the mechanism by which initial growth and survival differ between forest and transitional tree functional types.
In the seasonal tropics, fire commonly occurs in the dry season, irrespective of distinctions made between early versus late season fires, when microclimate and fuel characteristics are suitable to sustain fires of varying intensity (Hoffmann et al., 2012b; Prior et al., 2010; Sow et al., 2013; Veenendaal et al., 2018). Consequently, in pyrogenic savannas, fire effects and those of dry season drought are naturally entangled, confounding the influences of the two factors on tree seedling survival. However, tree functional types (e.g., forest, dry forest, transition and savanna tree species) may respond differently to fire and dry season drought. Thus, the relative importance of each factor may depend on tree species or functional type. For example, species that are less drought hardy, may have a higher chance of establishment in years of higher rainfall or shorter dry seasons and in places within the landscape where soil conditions improve soil moisture availability for tree seedlings. The role of fire may be less important in all these instances.
Therefore, it is relevant to decouple influences of fire from those of dry season drought on tree seedling survival for different tree functional types. Such insights are needed for understanding forest-savanna ecotone dynamics under different global change scenarios. More so, global change factors are thought to be increasing variability (and change) in climate (Mansoor et al., 2022; Owusu and Waylen, 2009), which can in turn, exacerbate the filtering effect of fire and dry season drought on tree recruitment in forest-savanna ecotones.
Not many studies have disentangled the effects of fire and dry season drought so far. Moreover, field experiments on tree seedling establishment success in forest-savanna ecotones that do not include dry season irrigation treatments (e.g., Cardoso et al., 2016; Gignoux et al., 2009, 2016; Issifu et al., 2019; Lawes et al., 2011) actually measure the combined effects of fire and dry season drought. Here, we decouple the effects of fire and dry season drought to determine the relative importance of each factor as a constraint to seedling establishment success of forest and transition tree species in a West African savanna.
We hypothesised that (1) the effects of both dry season drought and fire are larger on seedling establishment of forest species than on transition species, given their differences in drought and fire tolerances; (2) the interaction between fire and dry season drought on tree seedling establishment is significant, with a larger negative effect size for the treatment where fire and dry season drought are combined than for each factor separately, regardless of tree species or functional type, 3) root starch content correlates more strongly with stem size in forest species than in transition species; and (4) the relationship between seedling size at onset of the dry season and post-fire survival probability is stronger for forest species than transition species, as transition species are capable of surviving fires at much smaller sizes.
Materials and Methods
Study Site
The study was conducted at an experimental site on the Nyankpala Campus (9o 25′41” N and 0o 5′ W), University for Development Studies, Ghana. The area lies within the Guinea savanna agro-ecological zone of Ghana characterized by a unimodal rainfall regime. The wet season spans between May and October, followed by a dry spell from November to April. Average annual precipitation and temperature are 1100 mm and 28°C, respectively. Soils in the study site belong to the Nyankpala Series, described as shallow and taxonomically classified as Plinthustalf, sandy over-clayey skeletal phase (Serno & van de Weg, 1985) or Plinthic Lixisol (FAO, 2001). The soil in the experimental garden is sandy loam in texture with medium to coarse ironstones.
Tree Species
Tree Species Used in the Study and a Description of Their Origins and Mean Precipitation Under Which They Thrive. Species Origin: MSD = Moist Semi-deciduous Species, FST = Forest-Savanna Transitional Species (i.e. Transitional Species). Sources of Species Information: Hawthorne (1995); De Bie et al. (1998), Ametsitsi et al. (2020), World Agroforestry, Tropical Restoration Library and Useful Tropical Plants.

Experimental Treatments
Site preparation was done in April 2018 including ploughing of the experimental garden. In June 2018, we established 12 plots (blocks), each of about 37 m2. Twenty-four (24) seedlings were planted in rows in each plot, yielding a total of 287 seedlings in the experiment. Except for A. zygia and T. ivorensis which had fewer replications in some plots, all other species had four seedlings per plot randomly assigned to seedling positions. Final replications per treatment can be seen in Table 3. Spacing between and within rows of seedlings was 1.0 m × 0.7 m. In October 2018 (i.e., at the end of the wet season), we randomly assigned combinations of irrigation (irrigation vs no-irrigation) and fire (burn vs no-burn) treatments to plots.
We had four treatment combinations namely, “−irrigation+fire”, “−irrigation−fire”, “+irrigation+fire” and “+irrigation−fire”, respectively representing: a dry season with fire (i.e. the natural situation in fire-dominated humid savannas), dry season without fire (i.e. dry season drought in the absence of fire), fire with extended wet season (i.e. fire in the absence of dry season drought), and no fire with extended wet season (i.e. the control treatment with absence of both fire and dry season drought). Per treatment combination, three plots were assigned in a randomised block design. There were 12 seedlings per species (3 plots × 4 seedlings) in each treatment combination except A. zygia and T. ivorensis, as seedlings were insufficient for these species. Irrigation in the dry season involved giving, by means of hand watering, 33.6 L of water per plot per day from November 2018 to mid-March 2019, equivalent to 3860 mm of rainfall over the period. This amount of rainfall (although not corrected for the evaporation of the dry season) was slightly higher than the annual rainfall for the wettest forest species (Table 1). The fire treatment involved burning seedlings with pre-prepared grass fuel.
Fuel Preparation and Fire Application
Burning took place four months after transplanting and involved burning seedlings with dried grass, composed mainly of Pennisetum pedicellatum Trin. and Andropogon pseudapricus Stapf, which are the dominant grass species at the study location. We made grass fuel beds in each of the six plots to burn by placing 0.93 kg of dry grass in an area of 0.93 m2 on each row of seedlings, based on the fuel mass of ∼1 kg m-2 reported for humid savannas of West Africa (Hennenberg et al., 2006; Issifu et al., 2019). Average fuel bed height was 0.20 m yielding a fuel bulk density of 4.7 kg m-3.
Moisture content of grass (fuel) was determined on a dry weight basis from 12 samples as ((fresh weight – dry weight) / (dry weight) × 100). Burning was done on the afternoon of November 19, with all rows of seedlings (for each plot separately) burned simultaneously. Fire temperature was recorded for each burn (i.e., one measurement per plot) using an HI 935005 K-thermocouple thermometer (Hanna Instruments, Singapore). Probe cable was buried (for protection) with the full length of the probe sticking out of the ground close to the base of one randomly selected seedling to allow for the most accurate determination of fire temperature (Dayamba et al., 2010).
Plant and Biomass Measurements
Prior to transplanting, initial plant mass was determined for five random seedlings per species after oven-drying at 80°C. We measured seedling height and stem basal diameter just before implementing the fire treatment in November, after which we monitored their post-burn (re)growth until March 2019. At this time, we did the final census of live or dead seedlings in all plots and then excavated all live ones. Excavation was done as carefully as possible to recover as much root biomass as possible. We considered a seedling dead only if in addition to top-kill, the main root system near the surface had dried up. This was an appropriate definition because whereas fire usually results in top-kill, a live belowground part often results in regrowth among resprouters (Pausas et al., 2016). We measured final plant height and estimated total dry weight for all plants and regrowth (new shoot) dry weight for burnt plants after oven-drying at 80°C. We then calculated gains in plant mass as difference in mass between transplanting (species average) and final harvest (for each plant).
Measurement of Environmental Factors
Weather data for the experimental period (and few months before) were obtained from the Nyankpala weather station of the Savanna Agricultural Research Institute of Ghana-SARI (9-25-41″ N and 0-58′ 42″W), situated within 1 km of our experimental garden.
We measured volumetric soil moisture content (vmc %) at the end of the wet season (i.e., October 2018) using a theta probe (Delta-T, UK) at five locations (plot centre and four corners) within each of the 12 plots. This was to determine soil moisture content of our plots at onset of the dry season and to test if soil moisture content differed among plots assigned to receive either of the irrigation treatments. Also, at final harvest in March 2019, we measured soil moisture content gravimetrically at three depths (5-10 cm, 25-30 cm and 45-50 cm) and at three seedling positions, selected diagonally across plot (centre and two plot corners) for all 12 plots. Also, we sampled soil at two depths (10 cm and 20 cm) in three holes per plot for six plots (three each for burnt and unburnt) for analysis of soil chemical properties (Table S1). We estimated soil matric potential at onset and end of the dry season based on a drying cycle water release curve for soils in the experimental garden.
Statistical Analysis
We analysed all data in R (R Development Core Team, 2017). We tested soil moisture content (SMC) at the onset of the dry season for differences between plots to receive irrigation or no irrigation using linear mixed effects models (LMM) including plot as random factor. For this we used the lmer function in lme4 package (Bates et al., 2015) and generated ANOVA tables with statistical significance based on Satterthwaite’s method using the lmerTest package (Kuznetova et al., 2015). SMC at final harvest was analysed for fixed effects of irrigation × soil depth also in an LMM including random factors of soil pit within plot. Tukey pairwise contrasts for significant irrigation × soil depth were performed using the emmeans package (Lenth et al., 2019).
We analysed seedling survival with binomial error (logit link function) in generalised linear mixed models (GLMM) using the glmer function in lme4 package. First, we tested whether survival differed between tree functional types (TFTs) in a model that included fixed effects of TFT × irrigation × fire and random plot effect. As we also expected variation within TFT, we performed separate analyses for each TFT for fixed effects of species × irrigation × fire and random plot effect. Tukey pairwise contrasts were performed for significant species or interaction terms.
For seedling growth, we took a subset of the data (due to high post-fire mortality in some species) to include only species with sufficient replicates in the combined fire and dry season drought treatment to allow for testing interactions. We then performed an LMM separately for plant mass gain and seedling height at final harvest with species × fire × irrigation as fixed effects and random plot effect.
For both seedling survival and growth performance, we estimated effect size (Hedges’ g) for each treatment using the irrigation without fire treatment as the control treatment to estimate both magnitude (value of g) and direction (whether g was positive or negative) of effect for the other treatments. Estimating effect size is a useful addition to statistical testing because it offers a more practical meaning and is less strongly influenced by sample size unlike statistical significance.
We compared plant dry weight between irrigated and non-irrigated plants in an LMM at the TFT and species level separately. We tested root starch concentration for TFT × irrigation (for unburnt seedlings) and TFT × fire (for irrigated seedlings) in separate analyses due to insufficient replicates in some treatments at final harvest. We compared total root starch content between TFTs, using an LMM including root dry weight as covariate and random plot effect.
We assessed relationships (within each TFT) between pre-dry season stem diameter and dry season survival in separate GLMMs for fixed effect of fire × dry season with random plot effect and stem basal diameter as covariate. Prior to this, we had assessed correlations of stem diameter with other pre-dry season plant size variables (i.e., height and pre-transplantation plant mass). We also assessed the relationship of pre-dry season basal stem diameter to final root starch content to explain influences of pre-dry season plant size on dry season survival.
Results
Environmental Factors
Rainfall of the study area peaked in August and ended in October (Figure 1) with a total of 959 mm received within the experimental period of June 2018 to March 2019. Temperature, relative humidity and wind speed at the moment of the experimental burn were 30.7°C, 70% and 0.92 kph, respectively. Mean moisture content of grass used in the experimental burn was 3.0% and mean fire temperature recorded was 519°C. Soil moisture content in October (onset of the dry season) averaged 15.3% and did not differ significantly (F1, 10 = 1.70, p = 0.22) between plots to receive irrigation (at 15.7%) or no irrigation (at 14.9%). This equates to a matric potential of approximately −0.004 MPa to −0.006 MPa (i.e., no water stress for seedlings at this point). Overall, soil moisture content at the end of the experiment was lower in non-irrigated than irrigated plots, but there was also, a significant irrigation × soil depth interaction effect (F2, 68 = 20.3, p < 0.001). Soil moisture content was progressively higher with depth such that the difference between irrigated and non-irrigated plots was largest (and significant only) in the topmost (5-10 cm) layer (Figure S2), with matric potentials of −0.30 MPa for irrigated and −0.43 MPa for non-irrigated seedlings. Irrigated and non-irrigated seedlings in the 25 – 30 cm layer were under similar moisture stress (−0.20 MPa and −0.27 MPa), while matric potentials at 45 – 50 cm soil layer were −0.11 MPa and −0.42 MPa, respectively. Mean temperature and relative humidity (on the secondary axis) and total rainfall and evaporation (on the primary axis) for the experimental period in Nyankpala in the Guinea savanna ecological zone, Ghana. Red arrow indicates moment of fire event, black arrows indicate start of the experiment and final harvest. Data were obtained from Nyankpala weather station of the Savanna Agricultural Research Institute, which is located within 1 km of our experimental site. Data on total evaporation between March and August were not available.
Soils were slightly acidic to neutral and except for exchangeable Mg which was slightly higher in some burnt plots, soils in the experimental garden had low levels of available P and exchangeable Ca which were similar for all plots. Exchangeable K was generally moderate in the plots. Also, there was no effect of burning on many of the measured soil chemical parameters (Table S1).
Seedling Survival Responses to Fire and Dry Season
Statistics on Seedling Survival (From GLMM) and Plant Mass (From LMM) at Final Harvest. For Each Parameter, Comparisons are Done Between Tree Functional Types (TFT) and Then Among Species for Each TFT Separately. GLMM on Survival Included Fixed Effects of Fire, Irrigation and TFT (or Species). LMM on Plant Mass is for Unburnt Seedlings, Testing Fixed Effects of Irrigation and TFT (or Species). For Transition TFT, the Three-Way Interaction Term was Removed. Effect Sizes Associated With Survival and Plant Mass are Presented in Table 3 and Table 4 Respectively.

Survival Proportions and Effect Size Estimates (Hedge’s g) for the Various Experimental Treatment Combinations for Seven Tree Species in a Common Garden Experiment in the Guinea Savanna of Ghana. Effect Size was Estimated for Each Treatment Combination Relative to Irrigation Without Fire Which Represented the Control Treatment (With Best Possible Outcome for Seedling Survival). Effect Size Could not be Calculated for A. zygia and T. ivorensis due to Too Few Survivors at the End.

Among forest species, we observed the largest difference in seedling survival (across irrigation and fire treatments) between A. zygia (at 0.75 ± 0.31) and T. superba (at 0.42 ± 0.36) (Table 3) although species effect was not significant (Table 2). Also, there was no effect of irrigation or species × fire interaction on seedling survival (Table 2). Survival, across treatments, of the transition species was 0.96 ± 0.04 for K. senegalensis and 0.79 ± 0.14 for A. africana (Table 3), but species effect was not significant (Table 2). Across the two transition species, survival was significantly higher among irrigated (0.96 ± 0.91) than non-irrigated (0.79 ± 0.74) seedlings, but there was no effect of fire or of its interaction with irrigation on seedling survival (Table 2).
Effect size estimates (Hedge’s g) for the experimental treatments (Table 3) showed that across all species, the no-irrigation with fire treatment had the largest (adverse) effect on seedling survival. This effect was ∼ 4 times larger for the forest species (mean d = −3.70) than the transition tree species (mean d = −0.99). Between the two transition species, the effect of no-irrigation with fire treatment was ∼ 1.5 times larger for A. africana than K. senegalensis. Among the forest species, this effect was largest for T. superba as all seedlings of the species died in this treatment. For the rest of the forest species, there were only small differences in the effect size estimated for the combined fire and no-irrigation treatment (Table 2).
Also, the effect size of fire with irrigation treatment (i.e., fire alone, as the dry season drought effect is removed with irrigation) on seedling survival of forest species was 5 times larger than that of no-irrigation without fire treatment (i.e., dry season drought alone, as the effect of fire is removed). Among transition tree seedlings on the other hand, fire had no overall effect as all burnt seedlings resprouted, but the effect of irrigation was medium to large (−0.83) for both A. africana and K. senegalensis.
Seedling (re)growth Performance as Influenced by fire and Dry Season
Growth performance was analysed for the interactive effects of fire and irrigation for only three species, viz., K. anthotheca, A. africana and K. senegalensis, which had sufficient samples at the end of the fire and dry season. We found significant irrigation × species effect (F2, 86 = 7.54, p < 0.001) and fire × species effect (F2, 86 = 8.78, p < 0.001) on plant mass gain at final harvest (Table 2). Pairwise comparisons revealed lower mass gain for burnt than unburnt seedlings irrespective of irrigation for K. anthotheca and K. senegalensis, but not A. africana. Also regardless of fire treatment, mass gain was higher for irrigated than non-irrigated seedlings of K. anthotheca and K. senegalensis, but not A. africana.
We found a significant (F1, 10 = 91.8, p < 0.001) effect of fire on plant height, with unburnt seedlings being taller than burnt ones (Figure 2(a)). There was also a significant main effect (F1, 10 = 7.00, p = 0.027) of irrigation, with irrigated seedlings being taller than non-irrigated ones irrespective of species (Figure 2(a)). (a) Plant height and (b) mass gain at final harvest (after dry season/fire) for K. anthotheca (forest species) and Afzelia africana and K. senegalensis (both transition species) in burn + irrigation treatment combinations. There were enough samples after fire only for these three species. Different letters indicate significant differences among irrigation and fire treatments (Tukey pairwise comparisons at 0.05 level of significance).
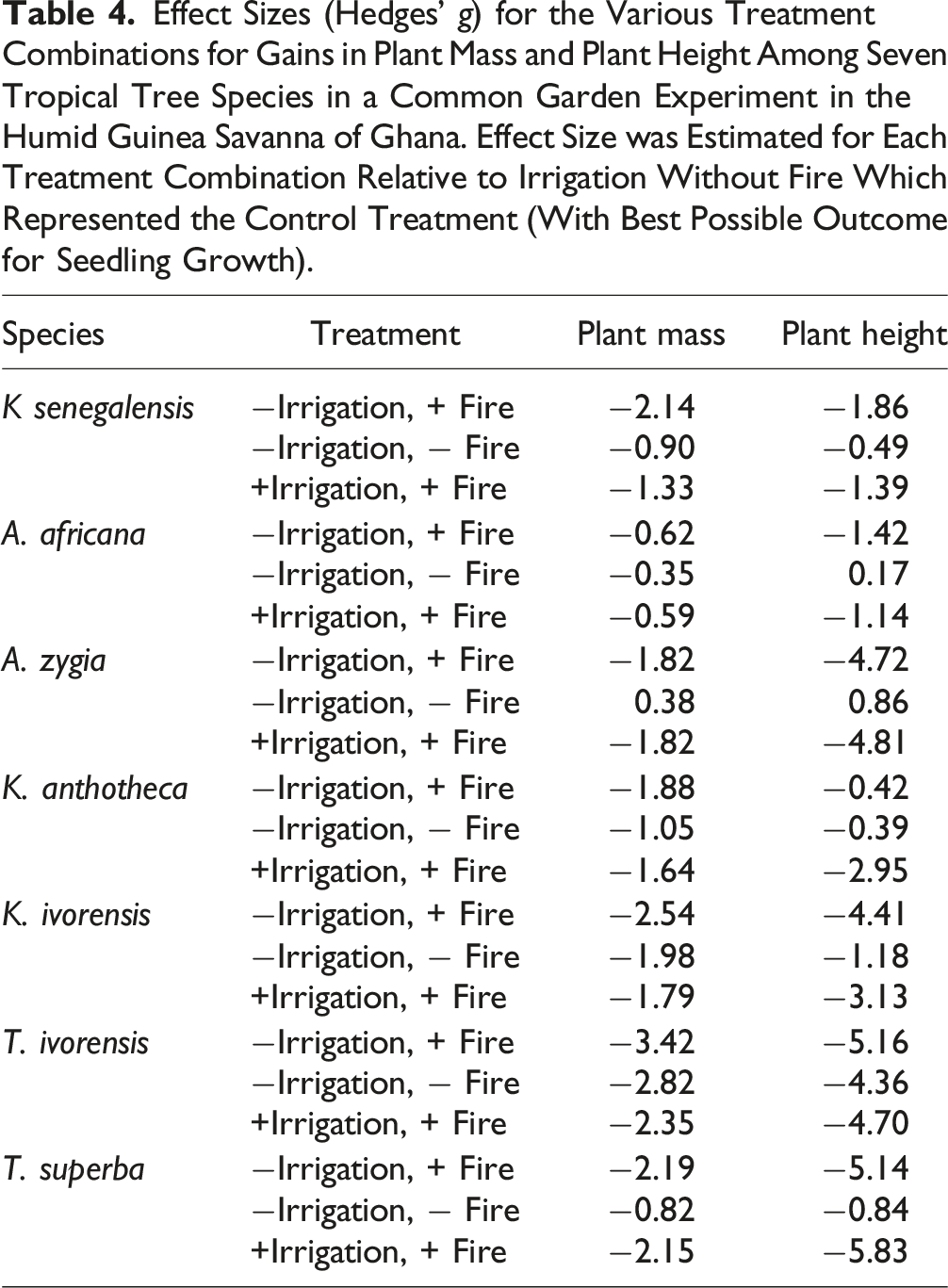
Effect Sizes (Hedges’ g) for the Various Treatment Combinations for Gains in Plant Mass and Plant Height Among Seven Tropical Tree Species in a Common Garden Experiment in the Humid Guinea Savanna of Ghana. Effect Size was Estimated for Each Treatment Combination Relative to Irrigation Without Fire Which Represented the Control Treatment (With Best Possible Outcome for Seedling Growth).
Seedling Size and Root Starch Reserves at Final Harvest
ANOVA Table From the LMM on Root Starch Concentration Compared Between Irrigated and Non-irrigated Seedlings and Only for Unburnt Seedlings.

In a separate analysis (for all species data, tree functional type regardless) for seedlings in the fire treatment, root starch concentration was significantly lower (F1, 43 = 43.2, p < 0.001) for burnt (but resprouted) (6.04 ± 4.0%) than unburnt (14.6 ± 6.5%) seedlings and there was no species × irrigation interaction effect. Root starch concentration also differed significantly (F1, 19 = 12.3, p < 0.001) among forest species, being highest for A. zygia (12.0 ± 6.0%), intermediate for K. anthotheca (9.2 ± 5.5%) and lowest for T. superba (6.8 ± 3.5%). Root starch concentration was also significantly lower (F1, 19 = 53.1, p < 0.001) overall (irrespective of species) for burnt (but resprouted) (4.8 ± 3.1%) than unburnt (12.0 ± 4.6 %) forest seedlings, and there was no species × fire interaction effect. It was significantly lower (F1, 18 = 24.5, p < 0.001) for burnt (but resprouted) than unburnt seedlings of the transition species, but with no species differences.
Root starch content (total reserves in roots) increased significantly (β = 0.10, t = 5.33, p < 0.001) with root mass. There was also an effect (F1, 54 = 7.41, p = 0.008) of tree functional type × irrigation interaction (Figure 3(b)) as the difference between tree functional types was significant only among non-irrigated seedlings. We also found that among burnt seedlings, root starch content increased (β = 0.10, t = 6.07, p < 0.001) with root mass. Burnt seedlings also had lower (F1, 41 = 23.3, p < 0.001) stored starch in roots than unburnt seedlings, but the difference between tree functional types was not significant (F1, 41 = 3.47, p = 0.069) (Figure 3(a)). Total root starch reserves for (a) burnt seedlings and (b) unburnt seedlings of forest and transition (ST) tree species in a common garden experiment. Root starch was tested for effect of burning (a) and effect of irrigation (b) in separate analyses which included only species with sufficient replicates at final harvest.
Relationship of Pre-Dry Season Seedling Size to dry Season Seedling Survival
There was a significant positive association between stem basal diameter and plant height at onset of the dry season, although the relationship was slightly stronger for forest species (β = 2.8, t = 14.4, p < 0.001) than for transition species (β = 2.2, t = 8.2, p < 0.001) (Figure S1a). Total root starch reserves, at final harvest, was significantly (β = 0.25, t = 3.20, p = 0.004) positively associated with pre-burn basal stem diameter for transition species, but not for the forest seedlings (Figure 5).
We found that survival probability for transition seedlings (across treatments) increased significantly (β = 0.84, z = 2.84, p = 0.004) with stem basal diameter, with survival guaranteed (on average) at ∼ 10 mm (Figure 4(b)). By contrast, the overall relationship between stem basal diameter and survival probability was not significant (β = 0.02, z = 0.28, p = 0.78) for seedlings of the forest species. Seedling survival probability was lowest in the +fire +irrigation combination and highest in the – fire + irrigation treatment regardless of stem diameter (Figure 4(a)). Predicted survival probability as a function of pre-burn stem basal diameter (proxy for seedling size) under various experimental treatments for (a) forest and (b) transition tree functional types in a common garden experiment in the humid Guinea savanna of Ghana.
Discussion
In this study, we decoupled the effects of fire and dry season drought on survival and growth of tree seedlings of forest and transition species of the forest-savanna ecotone. We hypothesised that the combined effect of dry season drought and fire has a greater effect than the separate effects of each factor. We expected poorer seedling performance for the forest species compared to the transition species in the 5-month long dry season. We also expected differential investment in root starch reserves to explain the differences in performance in the face of fire and dry season drought among species and between functional types.
We found that in the absence of fire, seedling survival was generally high (79% for forest species vs 90% for transition species) and did not differ between forest species and transition species, irrespective of irrigation treatment. This suggests that dry season drought alone (i.e., in the absence of fire) had no influence on seedling survival of forest species and transition species, contrary to our prediction which was based on known differences in drought tolerance between species from moist and those from drier environments (Gignoux et al., 2009; Hoffmann et al., 2004, Hoffmann & Franco, 2003). Mortality attributed to dry season drought effects have been reported for tree seedlings of forest species and transition species in other studies (Cardoso et al., 2016; Gignoux et al., 2009). However, in this study, tree seedling survival under dry season drought was high in the absence of fire for both forest species and transition species, possibly due to the large sizes attained by tree seedlings of forest species at the onset of the dry season. Some seedlings had reached a stem basal diameter of 16 mm in the four-month growth period and prior to being exposed to the five-month long dry season.
Rainfall was high at the experimental location (Figure 1) such that at the end of the experiment, soil moisture content did not differ between irrigation treatments beyond the top 10 cm soil layer (Figure S2). Therefore, our irrigation treatment appeared not to have resulted in a large enough difference in soil moisture content. Consequently, seedlings which had roots in the 25 – 30 cm soil layer and beyond (and that was nearly all species, Figure S3) possibly avoided drought stress severe enough to cause mortality. However, soil matric potentials did differ between irrigation treatments, although their difference also decreased with depth.
Additional reason for the high wet season plant growth may include the absence of competing grasses, which is linked to suppression of plant growth and survival (Barbosa et al., 2014; Issifu et al., 2021). Plants were, thus, large enough to withstand the 5-month long dry season due to the specific experimental conditions in this study. Perhaps, a longer dry season (for e.g., of six, rather than five months) drought would have made clearer soil moisture differences between irrigations treatments. This represents a limitation on a conclusion that fire is more important than dry season drought. Nonetheless, as there were differences in soil matric potentials, which are more meaningful to plant water status than moisture content alone, this conclusion may still be valid.
Fire decreased survival of tree seedlings of forest species but not of tree seedlings of transition species, consistent with our prediction, and in line with similar studies (Gignoux et al., 2009, 2016; Hoffmann et al., 2004; Issifu et al., 2021). This was expected as starch concentration in roots, which is often linked to resprout capacity, was higher for transition species than forest species as reported for savanna species versus forest species in other studies (Boonman et al., 2020; Hoffmann et al., 2004; Issifu et al., 2019; Wigley et al., 2019).
We hypothesised a positive association between seedling size and root starch concentration to explain higher dry season drought or fire survival for larger than smaller plants. We found that stem basal diameter (which was a better predictor than plant height), as a measure of seedling size, at the onset of the dry season significantly predicted root starch concentration at final harvest for the transition species, but not for the moist forest species, contrary to our prediction (Figure 5). However, larger root mass at final harvest was associated with higher root starch content (total stored starch in roots) even for the forest species (Figure 3). We had expected a poorer relationship for transition species as savanna species require less biomass, compared with forest species, to survive fire (Gignoux et al., 2016). As an important addition, our finding suggests that transition species invest in starch storage much earlier and thus have more starch reserves (at smaller sizes) than forest species (Figure 3). Therefore, survival was 100% for stem basal diameter of 10 mm (across treatments) for seedlings of transition species whereas seedlings of forest species did not attain complete survival (not even for the largest individuals) at sizes attained in this study (Figure 4). This may explain why fire and dry season drought survival of forest species did not necessarily increase with age (Gignoux et al., 2009) and why transition species required several times less biomass, than required by forest species, to survive fire (see also Gignoux et al., 2016). Relationship of pre-fire stem basal diameter to root starch content of unburnt seedlings at final harvest for forest and transition tree species. Root starch data include both irrigated and non-irrigated seedlings (not statistically different and so not distinguished here).
Due to the lack of capacity to regrow from belowground buds, forest species need to avoid aboveground tissue loss (top-kill) (Hoffmann et al., 2012a). However, compared to transition species, forest species reach the “threshold of fire resistance”, which is the size sufficient to avoid top-kill (Hoffmann et al., 2012a), at much bigger sizes. Therefore, while the forest species in this study were big enough to survive the dry season drought (in the absence of fire), they were largely killed by fire (all seedlings including those of transition species were top-killed). This finding suggests that forest species may survive dry season drought at sizes smaller than required to survive fire. Fire, thus, appears to be a stronger constraint to seedlings of forest species than low soil moisture content during the dry season where conditions are conducive to their establishment. This may justify why the high mortality of seedlings of forest species observed in dry season fire experiments is often regarded as a fire effect (Cardoso et al., 2016; Gignoux et al., 2009, 2016; Issifu et al., 2019; Lawes et al., 2011).
We observed the largest effect sizes on seedling performance in the combination treatment of dry season drought with fire. This effect, which represents the natural occurrence of fire and dry season drought, was stronger than the separate effects of fire and irrigation, consistent with our prediction. This may perhaps reflect the influence of soil moisture on survival and regrowth of resprouting plants (Pausas et al., 2016; Pratt et al., 2014) The interactive effect was also stronger for forest species than for transition species with the implication being that encroachment of forest species in savannas that are moist enough is only possible under a diminished fire regime (Louppe et al., 1995; Mitchard & Flintrop, 2013; Veenendaal et al., 2018). Otherwise, transition species (rather than forest species) may initiate forest encroachment in savannas (Cardoso et al., 2016; Hennenberg et al., 2005) and also explains the existence of dry forest communities with tree species typically adapted to regular fire (Swaine, 1992; Ametsitsi et al., 2020).
Seedling growth in the dry season was limited, possibly due to unfavourable microclimatic conditions (Figure 1) as irrigation in a common garden could only increase soil moisture without ameliorating microclimate. However, higher resprouting among irrigated and burnt seedlings yielded an overall significant effect of irrigation on plant mass and height (Figure 2). Also, higher mass gain among unburnt and irrigated than non-irrigated seedlings suggest that seedlings still grew regardless of any constraints imposed by microclimate. Differences in resprout and subsequent regrowth between tree functional types is largely attributable to differences in root starch reserves (Figure 3) also reported in other studies (see for example, Issifu et al., 2019; Wigley et al., 2019). Root starch reserves were lower for burnt than unburnt seedlings (Figure 3) likely because stored reserves were re-invested in regrowth among burnt seedlings which also explains why starch reserves (post-fire) were lower for transition species than forest species (Figure 3(a)) since regrowth was also higher for seedlings of transition species.
Overall, our study showed that irrigation during the dry season had a positive effect on seedlings of transition species but not on seedlings of forest species. Fire negatively influenced tree seedlings of forest species only. Overall, the combined effect of fire and dry season drought had the strongest effect on forest species, which thus face the greatest recruitment bottleneck in the forest-savanna ecotones.
Implications for Conservation
This study and several others (Boonman et al., 2020; Gignoux et al., 2016; Hoffmann et al., 2004; Issifu et al., 2019) have shown that recruitment of forest species is adversely affected by fire. Additionally, this study shows that fire effect is even stronger when combined with dry season drought. Thus, the mosaic of forest, transitional and savanna within the forest-savanna ecotone reflects the influence of fire and dry season drought. The implication being that any changes in these elements (fire, dry season drought and their combinations) will influence dynamics of the various communities within the forest-savanna ecotone. For example, increased fire frequency and intensity may result from global environmental change factors, particularly climate change (Mansoor et al., 2022). This could lead to transitional forest communities, composed of forest-savanna transition species, expanding at the expense of forest communities dominated by “true” forest species.
Supplemental Material
Supplemental Material - Decoupling the Effects of Fire and Dry Season Drought on Seedling Establishment Success of Tree Functional Types in Humid Savannas
Supplemental Material for Decoupling the Effects of Fire and Dry Season Drought on Seedling Establishment Success of Tree Functional Types in Humid Savannas by Hamza Issifu, Frank van Langevelde, Vincent Logah, George K.D. Ametsitsi, Jelle Klijn, Paolo Satorelli, Millicent Tanson, Hypolite Bayor, Philippine Vergeer and Elmar Veenendaal in Tropical Conservation Science
Footnotes
Acknowledgements
Wageningen University and Research with support from the Academy Ecology Fund of the KNAW made field work possible. We are grateful to Adnan Sumaila, Atampuri Amidu and Abdallah Tajudeen for assisting in field data collection and to Jan van Walsem for laboratory analysis of plant samples.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research: This work was supported by the Wageningen University and Academy Ecology Fund of KNAW [grant number J1608/Eco/G437].
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.