
Abstract
Highlights
• 24 wild mammal species use small oil-palm plantations in a highly deforested region in Mexico • Oil-palm plantations closer to forest remnants had higher mammal diversity • Presence of some species was associated to forest cover, distance to forest, and plantation age • Farmers associate mammals with both benefits and costs; these perceptions need to be considered when designing conservation measures • Complementing ecological sampling with interviews can improve our understanding of the oil-palm socioecosystem
Introduction
Tropical forest regions harbor the greatest terrestrial biodiversity on the planet, but they also suffer serious threats from human activities, in particular agriculture (Pendrill et al., 2022). Thus, a major challenge is to design landscapes that meet a balance between agricultural production, and biodiversity conservation (Arroyo-Rodríguez et al., 2020). This is particularly relevant for animals that have large area requirements, because their conservation must also occur beyond the boundaries of forest remnants (Payán & Boron, 2019). Wildlife conservation in human-modified areas is strongly affected by landscape composition and configuration (Almeida-Maués et al., 2022; Arroyo-Rodríguez et al., 2020), but it also depends on social factors that shape human-wildlife coexistence (Pooley et al., 2021).
One of the most rapidly expanding agricultural covers in humid tropical landscapes is oil palm (Elaeis guineensis). Despite its profitability, oil palm is controversial because in some countries it has been a major driver of deforestation and biodiversity loss (Savilaakso et al., 2014; Meijaard et al., 2018, 2020). However, in other regions (e.g., some parts of Mesoamerica) most plantations are established on lands that were previously used for agriculture (Furumo & Aide, 2017). In such cases, oil palm may even replace agricultural covers that are less biodiversity-friendly, such as cattle pastures (Boron et al., 2019; Gilroy et al., 2015).
Much of the information about the negative effects of oil palm on biodiversity comes from very large estates (Meijaard et al., 2018, 2020). Although less studied, oil palm can also be produced in smallholdings embedded in more heterogeneous landscapes. It has been argued that smallholdings have a higher potential for both conserving biodiversity and alleviating rural poverty (Azhar et al., 2017; Saadun et al., 2018). In several Mesoamerican regions, smallholdings are the dominant production system (Furumo & Aide, 2017). For example, in Mexico 95% of the 112,452 ha of oil palm is in smallholdings, one quarter of which is in the state of Tabasco (SIAP, 2020). Tabasco is a biodiversity hotspot with a long history of deforestation. Approximately 70% of Tabasco’s area is currently covered by agriculture (Hidalgo-Mihart et al., 2016), and oil palm is expected to expand (SAGARPA, 2017).
In this study, we collaborated with a local association of smallholders in Tabasco to increase our understanding about mammals in small oil-palm plantations, with the aim of guiding future conservation efforts. We had three specific objectives: (1) determine which mammals are using small oil-palm plantations in the study region; (2) assess the effects that landscape variables (forest cover and distance to forest) and plantation variables (area and age) have on the diversity and composition of mammal assemblages; and (3) determine farmers’ perceptions regarding benefits and/or costs associated to mammals. To meet our objectives, we gathered data through a combination of camera trapping and interviews. Collaboration with local farmers is key to developing participatory processes that might facilitate the co-design of conservation efforts (Saadun et al., 2018; Massarella et al., 2021).
We focused on non-volant mammals with body masses > 0.5 kg for several reasons: (i) they have been less studied in the context of oil palm in Latin America (Pardo et al., 2019), particularly in small plantations (Knowlton et al., 2019); (ii) they perform important ecosystem functions but are highly threatened by anthropogenic disturbances (Bogoni et al., 2022; Lacher et al., 2019), thus constituting a good proxy of a landscape’s value for biodiversity conservation (Payán & Boron, 2019); (iii) many have large area requirements, such that management at the landscape level is crucial (Brennan et al., 2022); (iv) some are charismatic species that are readily identified by people, which may help sensitize the local population about conservation needs (López-del-Toro et al., 2009); (v) they can be sampled with camera traps, which is a method that local community members could easily implement for long-term monitoring.
We expected that, by combining ecological and social sampling methods we might record more mammal species in oil palm than previously reported in Tabasco (Knowlton et al., 2019). Also, given the importance that the amount of remaining habitat and habitat connectivity have for biodiversity conservation in anthropic landscapes (Arroyo-Rodríguez et al., 2020; Watling et al., 2020), we expected to find negative effects of distance to forest and positive effects of the amount of forest cover. Also, we expected that older plantations, having more developed canopies, might favor mammal presence, compared to younger plantations. Regarding plantation area, we did not have an a priori expectation because, although plantations varied almost an order of magnitude in area, they were all smallholdings (< 60 ha). Finally, due to the interest of the local farmers in certifying their plantations as sustainable, we expected them to have perceptions that would be congruent with human-wildlife coexistence.
Methods
Study region and study plantations
Our study was conducted in the palm production region known as “De los Ríos” in the municipalities of Tenosique and Balancán (state of Tabasco, Mexico; 17°43’26.64’’-17°17’0.36’’ N, 91°28’20.15’’-91°05’22.73’’O; Figure 1). This region of Tabasco has lost most of its forest cover (77%) and has ∼ 8200 ha planted with oil palm (SIAP, 2020). The landscape is a mosaic of small forest fragments (mostly secondary forest in different stages of succession) embedded in an agricultural matrix dominated by cattle pastures and sugar cane. In this region, the owners of 118 oil-palm plantations (all, except one, are plantations < 60 ha) are organized in a local association, “Palmeros de los Ríos S.P.R. de R.L.”. One of the goals of the association is to be certified by the Roundtable of Sustainable Oil Palm (RSPO). RSPO is a global organization that aims to develop and implement guidelines for producing sustainable palm oil (Roundtable of Sustainable Oil Palm, 2023). This certification could benefit both farmers (e.g., certified palm oil has a higher price than non-certified oil) and biodiversity (e.g., certified palm oil comes from deforestation-free cultivation). (a) Map showing the location of the 11 small oil-palm plantations (black circles in large map) where mammals were sampled with camera traps, in a highly deforested region near the town of Tenosique (grey polygon in large map), in the state of Tabasco in southeastern Mexico (red circle in small inset map of Mexico). (b) Camera traps captured terrestrial/scansorial mammals > 0.5 kg in oil-palm plantations, such as the coyote shown here. (c) During interviews, farmers reported which mammal species they had seen in their oil-palm plantations, and they also described perceived benefits and/or costs associated with those mammals.
With the help of the local oil-palm association and satellite images (year 2020), we selected plantations that met the following criteria: (i) owners were willing to participate; (ii) they were undergoing the process of RSPO certification; (iii) they had a minimum of 5 ha and a maximum of 60 ha, and were separated by at least 2 km from any other selected plantation; and (iv) they were at least 5 years old, to ensure sufficient vegetation cover for wildlife. We selected 12 plantations distributed in an area of ∼970 km2 (Figure 1). We lost one plantation because a camera trap was stolen; the remaining plantations varied in size between 6.7 and 55.5 ha and were between 6 and 20 years old at the time of the study (Table S1).
Mammal sampling with camera traps
We sampled terrestrial/scansorial mammals with body masses > 0.5 kg using camera traps (Browning Strike Force BTC-5HDPX, Bushnell Core 24 MP, Bushnell Trophy Cam 8MP-UV 557, and Acorn 6210) without bait. We deployed 2 camera traps in plantations ≤ 25 ha and 3 camera traps in larger plantations (with ≥ 200 m between camera traps), for a total of 26 camera traps. Camera traps were placed 0.5 m above the ground and remained continuously active for 12 months (27 October 2020-11 December 2021). The total sampling effort was 8,662 trap-days. We used the software digiKam 7.5.0 (PIXLS.US, 2022) to generate and assign species labels to photographs. Subsequently, the metadata of the images (date, time, species) were extracted with the ‘camtrapR’ R package (Niedballa et al., 2016) to obtain the number of independent records per species. Photo records were considered independent when ≥ 60 minutes had elapsed since the previous photograph of the same species.
Landscape and plantation predictors
To obtain the values of both landscape variables (forest cover and distance to forest) we used a Sentinel 2 image (downloaded November 2021 from https://earthexplorer.usgs.gov), which was analyzed (including an atmospheric correction) with the software Quantum GIS 3.24.3 (QGIS Development Team, 2021). To estimate the percentage of forest cover (pooling primary and secondary forest) in the landscapes surrounding plantations, we generated an 800-m buffer around the perimeter of each plantation (i.e., the buffers had the same shape as the plantations). We chose an 800-m distance by considering two criteria: (i) a distance that allowed us to reach a compromise between larger and smaller home ranges of the different mammal species we expected to find; and (ii) a distance that avoided overlap of the landscapes surrounding study plantations. Then, we performed a land cover supervised classification using eCognition 9.0.1 (Trimble, 2014). To estimate the distance to forest, we first found the centroid of each palm plantation using the QGIS Field Calculator and then measured the distance from the centroid to the nearest edge of the nearest forest fragment that was ≥ 50 ha (either primary forest, or secondary forest with a canopy height ≥ 3 m). Values of plantation predictors (age and area) were obtained from plantation owners (Table S1).
Interviews with farmers
We conducted 12 interviews in situ (one interview per plantation). For smaller plantations, one farmer was interviewed; for larger plantations, 2-3 farmers answered the interview collectively (Frey & Fontana, 1991). For each plantation, we only interviewed farmers who had worked there for more than 6 months (Azhar et al., 2014). We obtained verbal informed consent to record the interviews. We also built rapport with our interviewees through active listening and by providing a comfortable environment. Interviews felt like an enjoyable dialogue with farmers sharing experiences and anecdotes. All participants remained anonymous and were able to reject any question or stop the interview for any reason at any time.
Interviews had two purposes: (i) to gather local ecological knowledge regarding the presence of mammal species in plantations, a method that proofed to be very useful in a previous study with mammals in oil palm (Azhar et al., 2014); and (ii) to assess how farmers perceive each mammal species in terms of benefits and/or costs, which is useful baseline information for the co-design of successful management guidelines (Saadun et al., 2018). During interviews we asked open and closed questions (Appendix S1). Of the mammals reported for the state of Tabasco (Hidalgo-Mihart et al., 2016), 32 (pooling two species of rabbits into one genus, Sylvilagus) are species with body masses > 0.5 kg that could occur in the study region (Reid, 1997); 7 of these species are mostly arboreal and were thus unlikely to be recorded by our camera traps. Following Azhar et al. (2014), we showed a photograph of each of these 32 species to farmers, using their common names in Spanish. For each species, we asked them if they had seen it in the plantation and/or in other types of vegetation in the region. For those species they had seen in the plantation, we asked them if it consumed the oil-palm fruit and if they thought the species was beneficial or harmful in other ways (Appendix S1).
Data analyses
To determine the list of mammals found in the oil-palm plantations studied (objective 1), we used data from camera traps and information obtained in the interviews.
To assess the effects that landscape and plantations variables have on mammal assemblages (objective 2), we only used camera-trap data, not interview data. Species diversity estimation was done using Hill numbers, considering numbers of order 0 (q0), 1 (q1), and 2 (q2). These indices vary in the importance they give to species abundances. Commonly interpreted as species richness, q0 gives no weight to abundances. The exponential of the Shannon diversity index, q1, weighs all species according to their frequency. Finally, q2, the inverse of Simpson concentration index, gives higher weight to the most abundant species. To calculate diversity estimates we used the number of independent photo records and the function estimateD in the ‘iNEXT’ package for R (Hsieh et al., 2020). To ensure our estimates of species diversity were not biased by differences in sampling effort among sites (Chao & Jost, 2012), estimation was done at a sampling coverage of 0.95.
To evaluate how species diversity is affected by landscape and plantation predictors, we fitted linear models. First, we evaluated collinearity among predictors by calculating the Variance Inflation Factor (VIF); all VIF values were < 3. Given that we had four predictor variables but only a sample size of 11 plantations, statistical inference was done by fitting models with all the possible combinations of a maximum of two predictors. These models were then compared using the Akaike Information Criterion corrected for small sample sizes (AICc). We considered as best-fitted models those with AICc values lower than the AICc of the null model. Then we used the weights of the best-fitted models, in comparison to the weight of the null model, to assess the importance of predictors included in the best-fitted models (Anderson, 2008). To fit linear models and obtain the associated AICc and weight values, we used the lm and dredge functions in the ‘base’ and ‘MuMIn’ packages for R (Barton, 2022).
The effects of landscape and plantation predictors on species composition were evaluated through a distance-based redundancy analysis (dbRDA) on a presence-absence matrix and using the Bray distance. Significance of the whole model and of the predictors was tested using anova-like permutational F tests (Borcard et al., 2018). The analysis was performed using the dbrda and anova.cca functions in the ‘vegan’ package for R (Oksanen et al., 2020). All analyses were done using the R v. 4.1.2 programming language (R Core Team, 2021).
To determine farmers’ perceptions regarding benefits and/or costs associated to mammals (objective 3), the interviews were recorded and transcribed. Information from the interviews was synthesized and organized in a table (see Results).
Results
Mammals using oil-palm plantations
Mammal species recorded with camera traps (Photo) or local ecological knowledge (LEK) in small oil palm plantations in the state of Tabasco, Mexico. For comparison, we also show species photo-captured by Knowlton and collaborators (Knowlton et al., 2019) in oil palm plantations in a different region of Tabasco. Two numbers are given per species in the Photo column: the percentage of independent photo-captures (from a total of 3,360 independent photo captures) and the number of plantations (out of 11) in which a species was photo-captured (upper and lower number, respectively). The LEK column shows the number of interviews (out of 12) in which a species was reported as having been seen in palm plantations. Numbers in the Knowlton column are also percentages of independent photo-captures (from a total of 1,010 independent photo captures; Knowlton et al., 2019). The last column indicates threat categories; codes shown in the second column correspond to those used in Figure 3. Species names follow the Global Biodiversity Information Facility (GBIF, https://www.gbif.org, January 2023).

FMammal species photographed or reported by farmers eating the oil-palm fruit.
aMexican government (TH: Threatened; SSP: Subject to special protection; SEMARNAT, 2010); IUCN Red List (EN: Endangered; CR: Critically endangered; IUCN, 2022).
bHighly arboreal species; all other species are terrestrial and/or scansorial.
Local farmers confirmed the presence in palm plantations of all mammal species photo-captured, except one, the ocelot Leopardus pardalis. Farmers also mentioned the presence of four additional species in palm plantations: the deer Mazama temama, the kinkajou Potos flavus, the cacomixtle Bassariscus sumichrasti, and the monkey Alouatta pigra. The latter three are highly arboreal (which is why they were not photo-captured) and of conservation concern. Regarding the other eight mammal species of which we also showed photos during the interviews, farmers reported having seen seven of them in the region, though not in palm plantations (white-lipped peccary Tayassu pecari, spider monkey Ateles geoffroyi, puma Puma concolor, jaguar Panthera onca, silky anteater Cyclopes didactylus, tayra Eira barbara, and Baird’s tapir Tapirella bairdii). The long-tailed weasel Mustela frenata was the only species that no farmer reported seeing, either in palm plantations or anywhere else.
Effects of landscape and plantation variables on mammal assemblages
Species diversity.- For the three diversity metrics (q0, q1 and q2), the best fitted models were the ones that included the distance from the forest and the second-best model was the null model. For q0 and q1 the models’ weight ratios indicated that the best fitted model had approximately twice the support compared to the null model (weight ratio for q0: 0.388/0.174 = 2.23; weight ratio for q1: 0.412/0.183 = 2.25), while in the case of q2 both models were equally supported (0.279/0.271 = 1.03; see weights of all models in last column of Table S2). Thus, for q0 and q1, but not q2, we found evidence that with greater distances from a plantation to the nearest forest remnant, the diversity of mammals using a plantation decreases (39% and 40% of the variance explained for q0 and q1, respectively; Figure 2). The other three predictors (forest cover, plantation age, plantation area) had no effect on mammal diversity metrics (Table S2). Fitted lines and their 95% confidence intervals for the relationships between mammal diversity (Hill numbers q0 and q1) and the distance between the oil palm plantation and the nearest forest remnant ≥ 50 ha (q0: R2 = 0.394; q1: R2 = 0.397). Mammals were sampled for 1 year with camera traps in 11 small oil palm plantations (< 60 ha) near the town of Tenosique in the state of Tabasco, Mexico.
Species composition.- The multivariate analysis to assess the effects of predictors on species composition yielded a significant ordination (F4,6 = 3.05, P = 0.011), in which the first axis explained 36% of the variation (F1,6 = 7.68, P = 0.002). Dissimilarities in species presence among sites were explained by forest cover, distance to forest, and plantation age, but not plantation area (Figure 3). Some species, in particular the agouti Dasyprocta mexicana, the paca Cuniculus paca, and the jaguarundi Puma yagouaroundi, tended to be present in plantations that were closer to a forest fragment and those that had higher forest cover in the surrounding landscape, while the fox Urocyon cinereoargenteus showed the opposite pattern. Finally, the presence of species such as the armadillo Dasypus novemcinctus, and the anteater Tamandua mexicana was associated with younger plantations. Ordination (distance-based redundancy analysis) of 11 oil palm plantations (black circles) and mammal species photo-captured in plantations (four-letter codes) near the town of Tenosique (Tabasco state, Mexico). Dissimilarities in species presence among sites were explained by forest cover (F1,6 = 3.54, P = 0.030), distance to forest (F1,6 = 3.11, P = 0.031), and plantation age (F1,6 = 3.29, P = 0.037), but not plantation area (F1,6 = 1.18, P = 0.355). See Table 1 for full species names.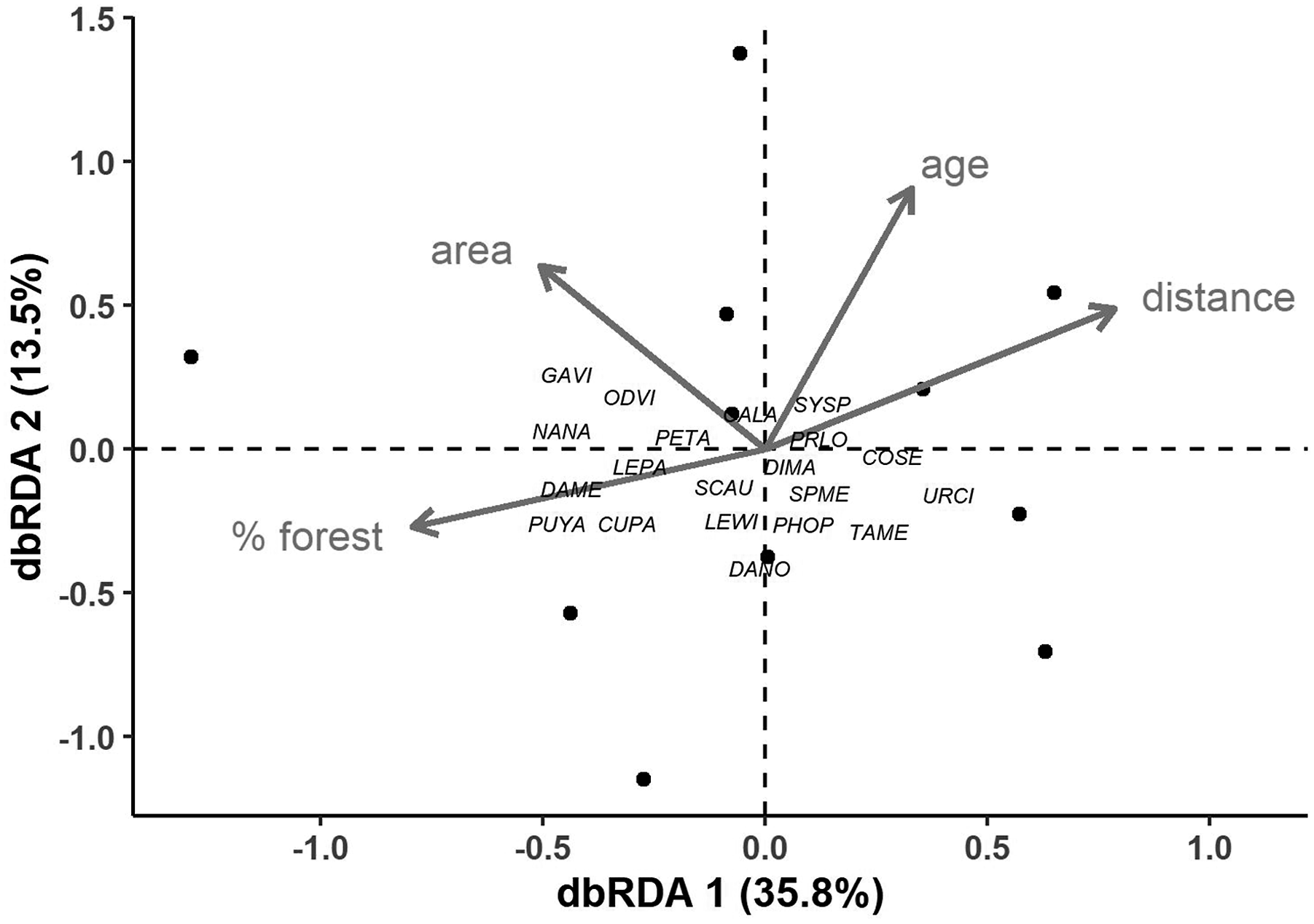
Farmers’ perceptions about the benefits and costs associated to mammals
Responses obtained from 12 interviews carried out with farmers working in oil palm plantations, to whom we asked about their perceived benefits and costs associated with mammal species they had seed in their oil palm plantations. Of the 24 mammal species recorded in oil palm plantations (see Table 1), farmers only mentioned benefits/costs for 19. We distinguish between benefits/costs that are associated to oil palm production (Palm), and those that are not (Other). In parentheses, the number of interviews (out of 12) in which a given response was recorded. See interview in Appendix S1.

Discussion
By combining camera-trapping and interviews we obtained a robust assessment of non-volant mammals > 0.5 kg using small oil plantations in Tabasco, Mexico. Our results show that characteristics of both the surrounding landscape and the plantations affected mammal assemblages, which coincides with the findings of previous studies (Meijaard et al., 2018, 2020; Payán & Boron, 2019; and references therein). Most importantly, plantations in closer proximity to forest remnants, mostly secondary forest, had higher diversity, emphasizing the importance of maintaining these patches in agricultural landscapes. Through interviews, we found that mammals were perceived as a source of both benefits and costs; the most commonly identified benefit was utilitarian. Overall, our study underscores the value of combining ecological and social dimensions for guiding wildlife conservation in anthropic landscapes.
Mammals using oil-palm plantations
The presence of mammal species recorded in oil palm through camera traps (except one) was corroborated by farmers. Furthermore, interviews allowed us to record four additional species, three of which are highly arboreal (howler monkey, cacomixtle, and kinkajou) and were thus unlikely to be photo-captured. We detected twice the number of mammal species than previously recorded in oil-pal plantation in Tabasco (Table 1). We believe that our more extensive sampling (1 year of photo-trapping in each plantation) allowed us to detect species that only occasionally use plantations and/or are rare in the region, such as the Mexican agouti, the collared peccary, the margay, and the Mexican hairy dwarf porcupine. This highlights the value not only of complementing information through local ecological knowledge (Azhar et al., 2014), but also of conducting extensive and long-term monitoring programs.
The mammals that we recorded in oil palm represent 75% of the 32 non-volant species > 0.5 kg that could potentially be found in the study region (Hidalgo-Mihart et al., 2016). On the one hand, this high percentage is encouraging, because it tells us that many mammal species still inhabit the region and are using oil palm plantations. On the other hand, however, it tells us that one quarter of the mammal fauna either avoids using oil palm or have very low abundances, including the two top predators (puma and jaguar). The mammal fauna of Tabasco has suffered a long history of decimation due to habitat loss and hunting (Hidalgo-Mihart et al., 2016). Consequently, the remaining fauna is dominated by species that are resilient to disturbances or even thrive in anthropic landscapes (Knowlton et al., 2019), as has also been found in a defaunated oil-palm region of Colombia (Pardo et al., 2019). In our study, three generalist mesopredator species (the racoon Procyon lotor, the coyote Canis latrans, and the opossum Didelphis marsupialis) strongly dominated the photo-captures (Table 1). Thus, even though we recorded a relatively high number of species, the mammal community seems to be uneven. This imbalance of ecological guilds, which threatens the adequate functioning of ecosystems, is a common pattern in hunted and deforested Neotropical landscapes, particularly where populations of top predators have suffered strong declines (Pires & Galetti, 2023).
Effects of landscape and plantation variables on mammal assemblages
Species diversity.- Of the four predictors evaluated, species diversity was only explained by distance to forest. Mammal diversity increased with decreasing distance from the plantation to the nearest forest patch ≥ 50 ha. A strong negative effect of distance to forest on mammal diversity was also found previously in Tabasco (Knowlton et al., 2019) and in Brazil (Mendes-Oliveira et al., 2017), but in Colombia the effect of this variable was weak (Pardo et al., 2018), and in Malaysia it had no effect (Azhar et al., 2014). This variability in results emphasizes the fact that wildlife responses in anthropic landscapes are highly contingent on regional characteristics, such as the species pool and the land-use history (e.g., Galán-Acedo et al., 2021).
As mentioned earlier, most forest remnants in Tabasco are secondary forests. While successional forests do not have the same value for mammal conservation as old-growth forests (Almeida-Maués et al., 2022), in highly deforested regions secondary forests are crucial for mammals, even patches that are early- or mid-successional (Knowlton et al., 2019; Pardo et al., 2018). In our study, the forest remnants that were nearest to most plantations (8 of 11) had a canopy height of only 3-5 m, indicating a relatively early stage of succession, and yet their proximity had a positive effect on mammal diversity in oil-palm plantations.
Unlike distance to forest, the amount of forest in the surrounding landscape had no effect on species diversity. This result was somewhat unexpected, because the amount of remaining habitat is one of the strongest and most consistent predictors of animal diversity in anthropic landscapes (Watling et al., 2020). However, as with the effect of distance to forest, results of previous studies in oil-palm have reported varying effects of forest cover on mammal assemblages. Two studies, one in Malaysia (Azhar et al., 2014) and one in Colombia (Boron et al., 2019) found a strong positive effect of forest cover on diversity, while another study in Colombia found only a weak effect of this predictor (Pardo et al., 2018). We acknowledge that the small number of plantations (11) in our study is a methodological limitation for assessing the effect of several predictors. It is also possible that the amount of variation in this predictor was not enough to detect its effect on species diversity (for 10 of the 11 plantations, % forest cover was between 6 and 30%; Table S2).
Plantation variables (area and age) had no effect on mammal diversity. Although the size of our study plantations varied almost one order of magnitude (6.7–55.5 ha), they were all smallholdings, and our result suggests that mammals are using plantations within this size range in similar ways. This is different from comparisons between large estates vs. smallholdings, where the latter tend to have higher levels of biodiversity, including mammals (Azhar et al., 2017). Regarding plantation age, we had thought that older plantations with higher and more developed canopies might hold more species of mammals. For the terrestrial/scansorial mammals sampled by our camera traps we did not find this, but an effect may occur for strictly arboreal mammals. For the three arboreal species reported by farmers, records were few (Table 1) and occurred in both young and old plantations.
Species composition.- Species composition was affected by all predictors, except plantation area. The two species of caviomorph rodents, the paca and the Mexican agouti, were predominantly present in oil plantations that were closer to a forest patch and surrounded by higher forest cover. In another region of Tabasco, the paca had been recorded in forest fragments but not in oil-palm plantations, while the critically endangered agouti, was not recorded in either vegetation type (Knowlton et al., 2019). Results emphasize the importance of maintaining forest fragments in agricultural landscapes, even small patches of secondary forest, for the conservation of caviomorph rodents, which are forest specialists (Mendes-Oliveira et al., 2017). We also found that some species, in particular the tamandua and the nine-banded armadillo, were more likely found in younger plantations. These mammals feed mostly (armadillo) or exclusively (tamandua) on small arthropods, and it is possible that younger plantations have a higher biomass of these animals (e.g., due to higher temperatures) than older plantations. For example, one study found higher herbivory damage in young vs. mature oil palm, suggesting higher abundance of herbivorous insects in young plantations (Woodham et al., 2019).
Farmers’ perceptions about the benefits and costs associated to mammals
Farmers were knowledgeable about which mammal species can be found in the study region. The fact that all species, but one, recorded by camera traps were also reported by farmers, reinforces the value of local ecological knowledge for informing wildlife research and management (Azhar et al., 2014; Owusu Afriyie & Opare Asare, 2020). Also, understanding how local people perceive and relate to animals can encourage community involvement and facilitate conservation in regions where humans and wildlife coexist (Pooley et al., 2021).
Interviewed farmers perceived important costs associated to mammal species that attack domestic animals and damage other crops. Although many of the mammals found in plantations were recorded to eat the oil-palm fruit (Table 1), damage to oil palm caused by these mammals was not perceived as serious. Benefits related to the ecosystem functions of mammals were not identified, with the exception of trophic control of harmful organisms. Most frequently, farmers recognized that a potential benefit of mammals was that they could be used as bushmeat. Similar responses, i.e., a certain degree of tolerance or indifference toward many mammal species, a negative perception of mammals that cause important damage, an emphasis on utilitarian benefits, and little awareness of the importance of mammals for the environment, are common in rural Neotropical settings (e.g., López-del-Toro et al., 2009).
Perceiving hunting as a benefit is problematic for wildlife conservation. Hunting is the second cause of local mammal extinctions in the Neotropics (Bogoni et al., 2022), and it needs to be controlled if conservation efforts in agricultural tropical landscapes are to succeed. In our study, however, even though farmers said that obtaining bushmeat can be a benefit of mammals, most of them also complained about external people hunting in the region. While this response might have been elicited by the fact that the RSPO certification bans hunting, it suggests a positive attitude towards wildlife, but one that they find difficult to enforce.
Implications for Conservation
Oil palm is a profitable crop and its prominence in Neotropical landscapes will undoubtedly increase (Furumo & Aide, 2017). We showed that small oil-palm plantations in southeastern Mexico are being used by a large proportion of the mammal fauna. However, we identified two key actions necessary for mammal conservation in these agricultural landscapes. The first, is to protect secondary forest fragments, which comprise most of the remaining native vegetation. Our results strongly suggest that mammals living in secondary forest fragments are venturing into nearby palm plantations to find resources. We think that a spatial arrangement in which oil-palm plantations are placed adjacent to forest fragments could have several potential benefits for wildlife conservation: (i) it could increase the carrying capacity of small fragments; (ii) it could lessen negative edge effects in forest remnants (Anderson et al., 2022), with plantations acting as a buffer between forest and harsher agricultural covers (e.g., cattle pastures); (iii) it could serve to increase connectivity at the landscape level. The second action is to work with the local community to raise awareness of the importance of mammals for ecosystem function, and to find ways in which they can successfully regulate hunting in the region. Both actions will require the concerted participation of different stakeholders, for the successful design, implementation, and enforcement of management measures.
Supplemental Material
Supplemental Material - Socioecological Assessment of Mammal Assemblages in Small Oil-Palm Plantations in a Highly Deforested Region in Mexico
Supplemental Material for Socioecological Assessment of Mammal Assemblages in Small Oil-Palm Plantations in a Highly Deforested Region in Mexico by Montserrat Franquesa-Soler, Fernando Ocampo-Saure, Francisco Mora, Gabriel P. Andrade-Ponce, and Ellen Andresen in Tropical Conservation Science
Footnotes
Acknowledgments
We thank Palmeros de los Ríos S.P.R. de R.L. for their collaboration in this project. We also thank the families of farmers for their hospitality and support during our fieldwork. We thank Jessie Knowlton and Ena E. Matas Zayas for sharing data with us and Shania Herrera Gómez for helping with ![]() . We thank two anonymous reviewers for comments that greatly improved the manuscript. M.F.-S. thanks DGAPA-UNAM for a postdoctoral fellowship and F.O.S. thanks CONACYT for a graduate fellowship (CVU 860583).
. We thank two anonymous reviewers for comments that greatly improved the manuscript. M.F.-S. thanks DGAPA-UNAM for a postdoctoral fellowship and F.O.S. thanks CONACYT for a graduate fellowship (CVU 860583).
Author Contributions
M.F.-S. and E.A. designed the project. M.F.-S. and F.O.S. collected the field data. M.F.-S, F.O.S., and G.P.A.-P. processed data from the camera traps. F.M., F.O.S., and G.P.A.-P. carried out the statistical analyses. M.F.-S. and E.A. wrote the first draft. All authors discussed the results and contributed to the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the UNAM’s Dirección General de Asuntos del Personal Académico (DGAPA), through the PAPIIT project IN211220.
Data Availability
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.