
Abstract
Neotropical snakes have extremely low detection rates, hampering our understanding of their responses to habitat loss and fragmentation. We addressed this gap using a limited sample (50 individuals, 16 species) across 25 variable-size insular forest fragments within a hydroelectric lake and four adjacent mainland continuous forest sites, in Central Brazilian Amazonia. The number of species recorded on forest islands (1.55
Keywords
Introduction
Habitat loss and fragmentation worldwide are driving unprecedented biodiversity losses (Haddad et al., 2015; Sala et al., 2000). In the tropics, hydropower development has become one such driver, often creating extensive archipelagic landscapes of forest islands. These islands consist of forest patches corresponding to previous hilltops, that are now isolated within a hostile open-water matrix (Jones et al., 2016). Analogous to patterns observed for oceanic islands (Wilson & Willis, 1975), the effects of insular habitat fragmentation can be assessed in light of Island Biogeography Theory (MacArthur & Wilson, 1967), which predicts lower numbers of species in small, isolated habitat remnants. Although there is a growing number of studies examining species responses to this type of habitat disturbance, most of them are skewed towards birds and mammals (Jones et al., 2016).
Within reptiles, the cryptic habits, effective camouflage, and typical nocturnality of snakes render them rarely detectable, exacerbating the costs of field surveys in tropical forests (Fraga et al., 2014). As a consequence, notwithstanding insights from temperate (Kjoss & Litvaitis, 2001; Luiselli & Capizzi, 1997) and sub-tropical regions (Wang et al., 2012), snake responses to habitat loss and fragmentation in the tropics are still poorly understood. Given their characteristics, including specific habitat requirements (Fraga et al., 2011, 2018), thermal constraints and limited mobility (Fitch & Shirer, 1971), snakes are expected to be particularly sensitive to habitat disturbance. In fact, declines in snake populations have been widely reported (Doherty et al., 2020), which are considered to exert serious consequences on ecosystem functioning given their ecological roles as meso- and top-predators (Zipkin et al., 2020).
Given the overall lack of knowledge on snake resilience to habitat fragmentation and the extremely high cost associated with their surveys in tropical forests (e.g. up to US$120 per individual encountered; Fraga et al., 2014), we here assess to what degree snake responses to habitat loss and fragmentation can still be detected based on an incomplete landscape-scale species inventory that was incidentally collected using a survey design primarily targeting lizards and small mammals. Data on snake assemblages were collected using 100 L-pitfall trapping which, despite the low trapping success (Ribeiro-Júnior et al., 2011, but see Cechin & Martins, 2000), are particularly useful to record small-bodied and (semi-) fossorial snakes (Ribeiro-Júnior et al., 2008). We surveyed 25 variable-size islands and four continuous forest sites within the Balbina Hydroelectric Reservoir and its immediate surroundings in Central Brazilian Amazonia. By the time sampling was carried out, islands had been isolated for 27 years which, given the longevity of relatively small bodied-size snakes (e.g., six to 10 years for Colubridae and Dipsadidae, Ernst & Zug, 1996; ∼3 years for Elapidae, Oliveira et al., 2005), is enough to allow up to nine generations to elapse. We therefore assumed that snake assemblages surveyed in this study had reached an extinction-colonization equilibrium. We hypothesized that many species have been gradually driven to local extinction in smaller and more isolated islands (MacArthur & Wilson, 1967), which are also subjected to more intense edge effects (Benchimol & Peres, 2015a; Malcolm, 1994).
Methods
This study was carried out in the archipelagic Balbina landscape in central Brazilian Amazonia (centroid coordinates: 1°41′33″S, 59°38′57″W; Figure 1A). The Balbina reservoir was formed in 1986, following the completion of a major dam on the Uatumã River, a left-bank tributary of the Amazon River. Within the 443,772-ha hydroelectric reservoir, a primary forest area of 312,900 ha was flooded (FUNCATE/INPE/ANEEL, 2000) and 3,546 forest islands were created. Most islands consist of dense closed-canopy terra-firme forest. The mean annual temperature in this region is 28°C and the mean annual rainfall is 2,376 mm (IBAMA, 1997).

Overview of the study area and snake species recorded. (A) Surveyed sites within the Balbina Hydroelectric Reservoir of Central Brazilian Amazonia: 25 islands (in red and highlighted by a 1,000-m buffer contour) and four continuous forest sites (CF1, CF2, CF3 and CF4; indicated by red rectangles). (B) Aerial photographs illustrating a mainland continuous forest site (from Google Earth: earth.google.com/web/), a large, a midsize and small island (photo credit: E. M. Venticinque). (C) Example of species recorded: only in continuous forest sites (1st row), in both continuous forest and islands (2nd row), only at islands (3rd row) and the two unique species recorded on mid-size islands (4th row). No species were recorded on small islands (≤30 ha).
We surveyed 25 islands ranging in size from 0.55 to 14,660 ha (mean ± SD: 199 ± 344 ha), and isolation distances from each focal island to the nearest mainland CF ranged from 44 to 11,872 m (4,351 ± 3,386 m). As a baseline, we also surveyed four control continuous forest (CF) sites that are widely distributed throughout the reservoir and spaced apart by 200 m and 1,500 m from the nearest reservoir margin. Overall, CF sites were characterized by the nearby presence of riparian habitats (<500 m). In the particular case of CF2, half of the sampling sites therein (N = 5) were positioned alongside riparian forest habitat along ∼7 m-wide perennial streams. Due to logistical constraints, riparian habitat could not be sampled at other CF sites, so that all but five sampling sites within CF2 were positioned across an elevation gradient comparable to that of the surveyed islands.
Terrestrial and fossorial snakes were surveyed using 60-m long traplines, each of which consisting of three pitfall traps (buried 100-L buckets 68 × 57 cm) spaced 20-m apart and connected by a 50-cm high plastic fence, as adapted from Ribeiro-Júnior et al. (2011). The number of traplines placed at each sampling site was approximately proportional to its area. This allowed us to obtain a higher number of individuals at larger forest sites, where overall trap density, and consequently the probability of an individual moving near a trap, was lower (Table S1 in Supplementary Material 1). When analysing the data, however, we accounted for potential effects of a higher sampling effort at larger sites by including the corresponding sampling effort as an offset variable. Given the spatial restrictions on small islands, alternative smaller traplines were established therein (see details on sampling effort per site in Table S1).
Surveys were carried out from April to November in 2014 and in the same season in 2015, thereby excluding months of particularly high precipitation in the study area. We sampled 67 traplines during 16 consecutive days at each site over two field seasons, except for two CF sites which were sampled only during either the first or second field season (Table S1). In total, we sampled 5,952 pitfall-trap-days. In each field season, we sampled simultaneously a set of 10–15 trap-lines. Different sets of trap-lines were surveyed during separate occasions, yet in the same order in both sampling seasons to minimise any seasonal effects. All traps were checked on a daily basis, captured snakes were identified in situ, considering the most recent taxonomic updates (Uetz et al., 2020), and immediately released in the trapsite vicinities. Snakes of a given species and similar body size were never captured along the same transect, so it is highly unlikely that any given individual was double-counted. This study followed the guidelines provided by the American Society of Ichthyologists and Herpetologists (Beaupre et al., 2004) and the institutional animal care and use committee of the Federal University of Rio de Janeiro (CEUA-CCS UFRJ, 2010), and was approved by the appropriate Brazilian government agency (SISBIO license No. 39187-4).
Based on high-resolution RapidEye© satellite imagery and using the ArcMap software (Environmental Systems Research Institute, 2018), at the landscape-scale, we calculated the Proximity index, which takes into account the distance to nearby islands and their aggregate area within a 500-m buffer around the sampling site (
Due to the low numbers of snakes recorded, we summed the number of records for each sampling site during the two field seasons, and within sampling sites where more than one trapline was deployed. Also, given the overall low sampling sufficiency observed (see estimated sample coverage in Table S1 and rarefaction curves in Figure S1), we calculated the estimated number of species using the function ‘ChaoRichness’, an asymptotic diversity estimation function corresponding to the order of Hill numbers: q = 0, using the ‘iNEXT’ R package (Hsieh et al., 2020). For subsequent analyses, we retained both the observed (Sobs) and the estimated number of species (Sest).
Patterns and predictors of both observed and estimated numbers of snake species were examined using Generalized Linear Models (GLMs) fitted with a Poisson and a Gamma distribution with a log-link function, respectively. To overcome any potential bias due to the inevitably higher sampling effort on larger forest sites, we considered sampling effort, defined as log10 number of trap-days, as an offset variable in all the models performed. Patch and landscape variables were highly correlated when both islands and CF sites were considered, but not when only islands were considered (except for
We then repeated GLMs for Sobs and Sest considering all the uncorrelated variables –
Results
On the basis of 5,952 trap-days, we recorded 50 snakes representing 16 species, 11 genera and six families, across 11 of the 25 islands surveyed and all four CF sites. Six species were exclusively recorded at CF sites, five on islands, and another five species were common to both sites (Table S3, Figure 1C). The most abundant species were the Dipsadidae Atractus torquatus (N = 9 records) and the Elapidae Micrurus hemprichii (N = 8), while only one individual (singletons) was recorded for five species. Excluding one record of an exceptionally large Lancehead pitviper Bothrops atrox (2,100 cm), snake mean body length (±SD) was relatively small: 660
The number of species recorded on forest islands (1.55
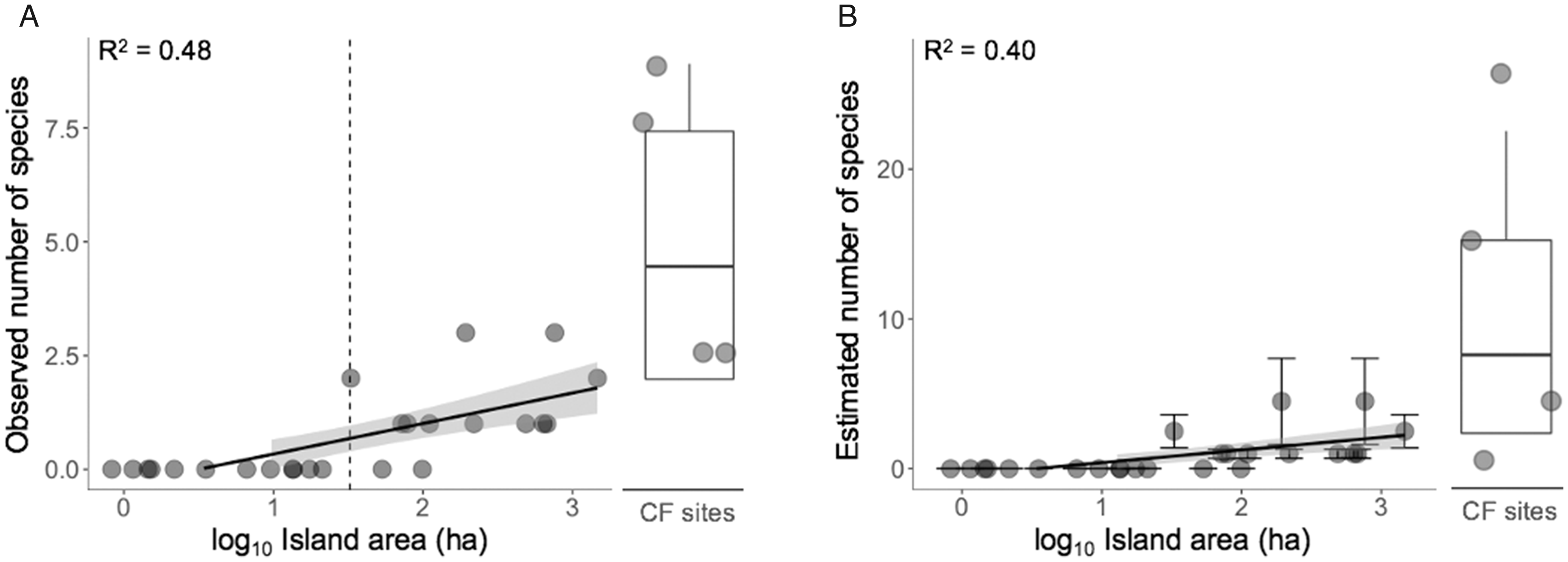
Species-Area Relationships of Snake Assemblages Across 25 Islands and Four Continuous Forest Sites Across the Balbina Archipelago Landscape, Considering the (A) Observed and (B) Estimated Number of Species. The vertical dashed line in (A) indicates the island size threshold (32.5 ha) below which all snake species could no longer be found. Lines are the model adjusted and shaded areas represent the 95% confidence regions. Boxplots indicate the median, 1st and 3rd quartiles, and minimum and maximum values of the observed/estimated number of species in continuous forest (CF) sites; dots indicate observed values.
When additional metrics related to island isolation across the 25 forest islands surveyed were considered, observed numbers of species was primarily negatively affected by distance to mainland (RI = 0.83), but positively affected by proximity to other islands or continuous forest sites (RI = 0.50) and island size (RI = 0.21) (Table S6). The estimated number of species was best predicted by island size (
Discussion
Forest patch area is the most important predictor of the persistence of multiple vertebrate taxa across nearly the same set of survey sites at the Balbina archipelago, including mid-large vertebrates (91%: Benchimol & Peres, 2015b), small mammals (69%: Palmeirim et al., 2018), lizards (87%: Palmeirim et al., 2017) and birds (81%: Aurélio-Silva et al., 2016). Despite the small sample size, forest area also explained ∼50% of the diversity of snakes at the Balbina archipelago. Our results are in agreement with general power of species-area relationships in explaining the remaining species diversity retained in the aftermath of habitat loss and insular fragmentation (Drakare et al., 2006; Lomolino, 2000). Similar findings were reported for snake assemblages on reservoir islands in sub-tropical China (Wang et al., 2012) and temperate fragmented landscapes (Kjoss & Litvaitis, 2001; Luiselli & Capizzi, 1997). Considering our disconcerting ignorance on the impacts of habitat loss and fragmentation on neotropical snakes, the clear species-area relationship reported here represents an important preliminary tool in predicting extinctions of forest serpent species across archipelagic landscapes. In addition, island isolation further apparently imposed a barrier for at least the relatively small-bodied snakes recorded in this study. Indeed, smaller-bodied snakes may be less adept at crossing the aquatic matrix, as noticed for other reptile species (Jenkins et al., 2007; Schoener & Schoener, 1984), which inhibits their recolonization of more isolated islands.
Although we were still able to detect snake responses to habitat loss and insular fragmentation, the very low number of snake records (N = 50) compared against our considerable sampling effort (5,952 trap-days) suggests that pitfall trapping is relatively inefficient in comprehensively surveying snake assemblages at Balbina. Indeed, we expected to have recorded only one-quarter (16 species) of the total snake diversity (up to 65 species) that likely occurs in this region, as observed on the basis of long-term research carried out at a terra firme forest 120-km from our study area, the Duke Reserve (Fraga et al., 2013). Most species we recorded were relatively small, terrestrial or fossorial (Fraga et al., 2013), with large-bodied and most arboreal species being able to immediately exit even the 68-cm deep 100-L pitfall buckets following captures (AFP, pers. obs.). Yet, given that small-bodied snakes are also those that are least known across the Neotropics (Vilela et al., 2014), our results highlight that opportunistic data recorded from sufficiently large pitfall traps (>68 cm in depth) can be used to improve our knowledge on those smaller species, although the applicability of our conclusions is limited in terms of arboreal and aquatic species.
The markedly higher species richness recorded across mainland CF sites is likely to result from the availability of riparian habitats therein, which have virtually disappeared from the archipelagic landscape given the widespread inundation of lowland areas. In Amazonian forests, riparian habitats harbour a unique set of snake species (Fraga et al., 2011). Therefore, the consistent suppression of lowland forest areas by hydropower infrastructure, including major riparian habitats, is expected to have an enormous impact on snake assemblages, as already observed for lizards (Palmeirim et al., 2017). Further investigation is still needed to obtain consistent information on snake responses to habitat loss and fragmentation in island systems. Notwithstanding, to prevent further severe biodiversity loss, including those of poorly known rare taxa, conservation policies should avert the additional construction of mega-dams that create a myriad of small islands, in addition to extensive reservoir lakes from which riparian habitats are eliminated.
Supplemental Material
sj-pdf-1-trc-10.1177_19400829211017366 - Supplemental material for Species-Area Relationships Induced by Forest Habitat Fragmentation Apply Even to Rarely Detected Organisms
Supplemental material, sj-pdf-1-trc-10.1177_19400829211017366 for Species-Area Relationships Induced by Forest Habitat Fragmentation Apply Even to Rarely Detected Organisms by Ana Filipa Palmeirim, Rafael de Fraga, Marcus V. Vieira and Carlos A. Peres in Tropical Conservation Science
Footnotes
Acknowledgements
We thank all 15 volunteers/field assistants who assisted in the field; the Reserva Biológica do Uatumã and its staff for logistical support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Áreas Protegidas da Amazônia (ARPA), Amazonas Distribuidora de Energia S.A., Associação Comunidade Waimiri Atroari; Rufford Foundation (grant 13675-1); Idea Wild; CNPq (grants 308.974/2015-8, 312465/2019-0 to MVV), and FAPERJ (grant E-203.045/2017 to MVV). AFP was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)—Finance Code 001, and the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No. 792678.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.