
Abstract
We aimed to assess how bats are affected by habitat transformation by comparing bat assemblages in four habitat types: primary forest, secondary forest, cocoa plantations and human habitations in the Dja Biosphere Reserve of southern Cameroon. Bats were sampled in the four habitat types using mist nets. During 126 nights, a total of 413 bats were captured, belonging to four families, 16 genera and 24 species. Ninety three individuals (17 species) were captured in the primary forest, followed by plantations (105 individuals, 14 species), human habitations (159 individuals, 10 species), and secondary forest (55 individuals, eight species). Megaloglossus woermanni was recorded in all the four habitats, and was the most abundant species (105 individuals). The analysis of bat assemblage between habitat types showed a statistically significant difference in species composition. The distribution of the six most abundant species (Epomops franqueti, Megaloglossus woermanni, Rousettus aegyptiacus, Dohyrina cyclops, Hipposideros cf. caffer and Hipposideros cf. ruber) was influenced by habitat types. Our results suggest that the decrease in species richness observed in disturbed habitats may be due to habitat perturbations of primary forest habitats. Therefore, it is important to examine the effects of habitat conversion at species level, as responses are often species-specific.
Anthropogenic disturbances in natural habitats are a major cause of global biodiversity crises, especially loss of tropical biodiversity (Frick et al., 2020). Among these disturbances, the rapid conversion of primary forest into agricultural areas can be considered as the main driver of tropical biodiversity loss (Ramalho et al., 2014). Sustained destruction of these complex ecosystems may lead to mass extinctions and massive loss of important ecosystem services provided by resident species (Ceballos et al., 2015; Morris, 2010). Forests provide important resources such as roosting sites and food for many bat species (Papadatou et al., 2011). Thus, the modification of the forest habitat through agriculture expansion and various developmental projects, as well as the accompanying roost site losses or disturbances have been identified as some of the major threats to bats worldwide (Frick et al., 2020). Indeed, bat species richness and abundance may be affected by these transformations, with the possible extinction of some species (Laurance et al., 2002; Turner, 1996).
The extinction of some bats species can lead to the degradation and loss of vital ecological processes (e.g. pollination, seed dispersal and insect predation), which in turn may affect various ecosystem services that benefit human societies (Frick et al., 2020). The consequence of land-use modifications in tropical forest is most often the fragmentation of forest or the degradation of initially continuous and intact native vegetation (Fahrig, 2003). Fragmentation and degradation lead to a loss of forest-dependent species and possibly favor non-forest generalists and edge species (Monadjem et al. 2010a). Changes in plant species composition and the forest structure affect bats’ food supply and their movement capacity and thus have direct impacts on maintenance of their ecological functions in forest regeneration (Meyer & Kalko, 2008). Effects of land-use intensity on species and abundance distribution lead to a stronger dominance of the most abundant species. Furthermore, species with restricted distributions are more likely to also become rare species in the local species abundance distribution and therefore are at high risk of being lost under intensive land-use (Simons et al., 2015). Though small fragments are known to support small bat population sizes and fewer species, they remain important for conservation (Loreau et al., 2001).
With about 1400 species, bats are the second most diverse group of mammals (Frick et al., 2020). Despite their high diversity, they are currently among the most persecuted group of vertebrates in the world (Bakwo Fils, 2009). Given their abundance, diversity and range of responses to habitat change, bats are recognized as an ideal group to study the effects of fragmentation in tropical forests (Meyer et al., 2010). Indeed, deforestation and habitat fragmentation is known to modify bat assemblages (Stebbings, 1995, Walsh & Harris, 1996). The responses of bats to habitat modification vary from a species to another (Cunto & Bernard, 2012). Generally, diversity and abundance of bat species decline with increasing deforestation (Duchamp & Swihart, 2008, Gehrt & Chelsvig, 2003). However, some generalist species are able to persist in modified habitats, due to the ability to access additional resources outside of remnant patches, such as artificial shelters and food resources (Duchamp & Swihart, 2008, Gehrt & Chelsvig, 2003). A study conducted in Paraguay by Gorresen & Willig (2004) revealed that, community diversity of insectivorous bats was highest in relatively undisturbed forests. Meanwhile, Cosson et al. (1999) showed that community diversity and abundance of frugivorous and nectarivorous bats was found to rapidly decline in smaller fragments. In contrast, Estrada and Coates-Estrada (2001) showed that bats within continuous forest and forest fragments in Mexico shared similar species richness.
Globally about 11.3 million ha of tropical forest is lost annually, including three to five million hectares in Africa (Food and Agriculture Organization [FAO], 2006). According to FAO (2006) deforestation rates are exceptionally high in West and Central Africa, with mean annual forest loss of 0.48–0.56% between 1990 and 2005. Marcoux (2000) estimated the annual deforestation rate in Africa at 0.7%, with Cameroon estimated to lose 80,000 to 200,000 ha annually (Ndoye & Kaimowitz, 2000). Deforestation is therefore a threat to conservation of biodiversity in Cameroon, particularly for bats conservation. Consequently, field data that helps to evaluate the responses of bat assemblages to habitat loss and land conversion due to anthropogenic activities is necessary. This data will help ecologists to understand how bat species richness and abundances are related to habitat perturbations caused by forest lost in Cameroon.
In this study, we investigated bat communities in the Dja biosphere reserve. With the aim of comparing diversity patterns between areas inside and outside the Dja fauna reserve. Specifically, we tested the following hypothesis: Abundance and richness of bats decreases over the habitat disturbance gradient. We predict that the bat species richness and abundance will be higher in primary forest than others habitats due to the greater structural vegetation complexity and stability found in this habitat. To this end, we assessed differences in species richness, abundance, and species composition between habitat types and evaluated bat responses to deforestation.
Study Site
The study was conducted in the Dja Biosphere Reserve situated in the southeast of Cameroon (Figure 1). The reserve lies in between latitude 2°40’ and 3°23’ North and longitude 12°25’ and 13°35’ East, and covers an area of about 5,26,000 hectares. It is the largest protected area in Cameroon (International Union for Conservation of Nature, 1987). The vegetation is described as semi-deciduous lowland tropical rainforest (Letouzey, 1968) and elevation is between 400 m a.s.l. and 800 m a.s.l. The climate is characterized by two wet and two dry seasons, with major and minor rainfall peaks generally occurring in October and May respectively. Four main habitat types occur in this reserve: Upland forest; Rapphia swamp; Uapaca swamp and the inselberg associated forest (Sonké 2004). During the last decades, activities such as continued human migration to the area, has resulted in vast areas of the original vegetation being cleared to provide land for commercial and subsistence agriculture. Currently, unsustainable commercial logging activities, hunting and trapping are causing serious degradation to the natural vegetation. These changes have inevitably affected the resident fauna, including bats.

Map Showed the Study Area and Sampling Sites at the Dja Biosphere Reserve (Cameroon).
Methods
Bat Sampling
We captured bats with mist-nets set at ground level in four habitat types (Primary forest, Secondary forest, Cocoa plantations and Human habitations) described based on anthropogenic disturbance and land cover (Figure 2):

Photos of Four Different Sampling Habitats Types in the Dja Biosphere Reserve (Cameroon). (a) Primary forest. (b) Secondary forest. (c) Cocoa plantation and (d) Human habitation.
Primary forest showing no evidence of disturbance. Situated at the centre of the Dja Biosphere Reserve, this part constitutes the Dja Fauna Reserve. It is characterized by the dense evergreen forest with a canopy rising from 30 to about 60m, forming a continuous closed canopy.
Secondary forest (more than 2 years old) showing evidence of disturbance resulting from wood exploitation and unsustainable commercial logging activities. The clearance of natural vegetation to provide land for commercial and subsistence agriculture, illegal hunting and uncontrolled bush burning have led to a serious degradation of the vegetation. It is characterized by road openings and had a slightly open canopy.
Cocoa plantations (more than 10 years old) characterized by the dominance of cocoa trees. Some fruits trees (Mangifera indica, Persea americana, Carica papaya, Dacryodes edulis, Garcinia kola, Cola nitida and Cola pachycarpa) are present. The canopy is slightly open due to the presence of some forest trees left specially to create shade. Other agroecosystems occur in the study site as well as coffee plantation and palm plantation. Cocoa plantation was chosen because they are polycultural agro-ecosystems contrary to the two others.
Human habitation characterized by dominance of houses. Some fruit trees (Mangifera indica, Carica papaya and Psidium guajava) are present. They are surrounded by small farms and the canopy is open.
Bats were sampled monthly between June 2018 and November 2019 in all four habitat types. Sampling was conducted at the rate of seven consecutive nights per month using 10 ground-level mist-nets per night (5 x 12 x 2.5 m, mesh, 16 mm; 3 x 9 x 2.5 m, mesh, 16 mm and 2 x 6 x 2.5 m, mesh, 16 mm). During each sampling night, mist-nets were opened from 6 pm to about 12 midnight along potential flight path of bats. The mist-nets were checked every 15 minutes in order to avoid severe entanglement to captured bats. Captured bats were held in bags prior to processing. We also investigated caves, hollow trees and other potential day roosts site for bat presence. Each captured bat was weighed (using a balance, nearest 0.5 g), measured using a Vernier caliper (Ecotone-Poland 150/0.1 mm) (Appendix A), aged (adult or juvenile based on ossification of phalanges) and sexed. Each individual bat was punched marked with a code on the wing before release in the capture site, in order to prevent data replication due to recaptures on the same night (Bonaccorso & Smythe, 1972). Bats were identified to species level in the field based on morphometric measurement aided by identification keys of Rosevear (1965), Hayman and Hill (1971), Patterson and Webala (2012), Van Cakenberghe et al. (2017) and ACR (2019). Skin samples were collected from the wing membrane of living bats for genetic analyses. The skin biopsies were collected using non-lethal Stieffel 3 mm biopsy punches (Pierce & Keith, 2011).
Data Analyses
Bat species richness for each habitat type was estimated using the observed species richness Sobs (Mao Tau) and the non-parametric estimator Chao 2 to construct rarefaction curves. These analyses were computed using software EstimateS 9.0 (Colwell, 2009). Chao 2 takes into account the number of undetectable species and differences in sampling methods. We constructed Rank-abundance curves to compare the number of species, relative abundances of each species, number of rare species and equitability in each habitat type (Stoner, 2005).
To investigate which parts of the species abundance distribution are affected by forest loss and habitat conversion, we focused on the dominant in our samples. For this, we used the dominance d (also known as Berger-Parker). Dominance d is calculated by dividing the number of individuals of the species that is most abundant (n) by the total abundance of all species in the community (N); it is independent of the abundance distribution and species richness of the sample, and is considered as informative and more robust than other more complex measures of dominance (May, 1975).
We used an analysis of similarity (ANOSIM – one way) and a Ward linkage hierarchical clustering based on Bray-Curtis distance to compare the differences in composition and species abundance between the four habitat types in the Dja Biosphere Reserve. ANOSIM combines Jaccard similarity index, which uses only presence/absence data for species and the Bray-Curtis index, which also considers abundance. The analysis produces values of R between 0 and 1. Values of R closer to 1 indicate higher dissimilarity between groups (Clarke, 1993). Those analyses were performed using PAST software (Hammer et al., 2001; Magurran, 2004). To identify the species contributing most to the differences between habitat types, we performed an analysis of similarity percentages (SIMPER; Clarke, 1993) with the R function SIMPER in vegan for each habitat type. SIMPER identifies the species that are most responsible for the observed patterns (e.g. the species that contribute the most to the dissimilarity between habitat types) by disaggregating the Bray-Curtis similarities between samples. The more abundant species is within a group, the more it contributes to the intra-group similarity, while a species with a consistently high contribution to the dissimilarity between groups is a good discriminating species (Clarke & Warwick, 2001).
Generalized linear mixed-effects models (GLMMs) were used to evaluate the differences in species abundance between habitat types (Zuur et al., 2009). Models were fitted using function glmer in the R package ‘lme4’ (Bates et al., 2015), assuming a Poisson error distribution and log-link function. Habitat type was included as a fixed effect and individual sites and month were specified as random, which allowed for appropriate control of pseudo replication within a site and to account for site-specific variance. Statistical significance of the fixed effect ‘habitat’ was determined based on a likelihood ratio test (Zuur et al., 2009). Because of low number of captures for most of the species during this study, this analysis was restricted to species with sufficient data of more than 20 captures (Hipposideros ruber, Doryrhina cyclops, Epomops franqueti, Hipposideros caffer, Megaloglossus woermanni, and Rousettus aegyptiacus). This method is more robust compared to more commonly use non-parametric tests such as the Kruskal–Wallis test.
Results
Bat Assemblages
During 126 nights, a total of 413 bats were captured, belonging to four families, 16 genera and 24 species (Table 1). The family Pteropodidae (frugivorous bats) was represented by 285 individuals (eight species), followed by Hipposideridae with 95 individuals (four species), Vespertilionidae with 27 individuals (nine species) and Nycteridae with 6 individuals (three species). Insectivorous bats represented 31.07% of all captures while frugivorous bats represented 68.93%. However, insectivorous bats had the highest species richness (66.66%; 16 species), while frugivorous bats had a lower species richness (33.34%; 8 species).
Number of Captures and Sampling Efforts Within the Four Habitat Types in the Dja Biosphere Reserve.

Fru: frugivorous; Nec: nectarivorous; Ins: insectivorous.
Quantitatively, Megaloglossus woermanni (a nectarivorous bat) was the most common species recorded (25.49%, n = 105), followed by Epomops franqueti (17.72%, n = 73), Rousettus aegyptiacus (17.23%, n = 71) and Hippossideros cf ruber (09.95%, n = 41). The other species represented 29.61% of all the bats captured. We found six caves, with five housing Hippossideros cf ruber and one housing Hippossideros cf caffer (cave 6) in primary forest. Seven trees as roost were also found in our study, where three in primary forest housing Dohyrina cyclops and four in cocoa plantation housing Dohyrina cyclops (trees 4 and 5), Nycteris thebaica (tree 6) and Dohyrina cyclops together with Nycteris hispida (tree 7). Concerning houses as roost, two houses containing Hipposideros cf ruber (house 1) and Pipistrellus nanulus (house 2) were found in human habitation (Appendix B).
Ninety three individuals (17 species) were recorded in primary forest, amongst which H. ruber was the most abundant (22 individuals), followed by Hipposideros caffer (20 individuals) and E. franqueti (19 individuals). In secondary forest, we recorded 55 individuals (eight species) amongst which M. woermanni was the most abundant species (23 individuals), followed by Doryrhina cyclops (seven individuals). In cocoa plantations we recorded 106 individuals (14 species) amongst which the most abundant species was M. woermanni (29 individuals) followed by E. franqueti (22 individuals) and D. cyclops (17 individuals). In human habitations, we recorded 159 individuals (10 species) and the most abundant species was M. woermanni (51 individuals) followed by R. aegyptiacus (45 individuals) and E. franqueti (27 individuals) (Table 1). In primary forest we recorded the highest number of species, followed by cocoa plantations, and human habitations. The secondary forest had the lowest species richness (Figure 3). Six species were sampled only in primary forest (Scotoecus hirundo, Myotis bocagei Scotophilus sp., Glauconysteris albogutata, Glauconycteris argentata and Hypsignathus monstrosus), only four species in cocoa plantations (Pipistrellus sp., Nycteris grandis, Nycteris thebaica and Macronycteris gigas), one only in secondary forest (Glauconycteris sp.) and human habitations (Myonycteris angolensis) respectively (Table 1). The percentage of singletons was highest in the primary forests (1.5), followed by cocoa plantations (0.97). In Human habitations (0.2) and cocoa plantations (0.2), we recorded the lowest number of singletons respectively.

Expected Species Accumulation (Mao Tau) Curves (Plus 95% Confidence Interval Curves) for the Four Sampling Habitats Types in the Dja Biosphere Reserve (Cameroon): Primary Forest and Cocoa Plantations Don’t Reach Asymptote; Secondary Forest and Human Habitation Reach Asymptote.
When the four habitat types are considered, the rarefaction curve (Mao Tau) of each habitat, did not reach asymptotes (Figure 3) suggesting that the sampling effort was not satisfactory. For human habitations and secondary forest the fitted rarefaction curves nearly reach asymptotes indicating that few species could still be captured with more sampling occasions. However, the rarefaction curves for primary forest and cocoa plantation did not reach asymptotes and more species would be detected with increasing sampling effort (Figure 3). The Chao 2 estimated species richness, indicated primary forest to be the richest habitat (Chao 2 = 26.63 species), followed by cocoa plantations (20.13), human habitations (10.67) and secondary forest (8.83). Based on this estimation, we achieved a level of species inventory completeness of 63.84% (17 species) for primary forest, 69.54% (14 species) for cocoa plantation, 90.60% (10 species) for human habitation and 93.72% (8 species) for secondary forest.
Rarefied bat species richness was significantly higher for primary forest and cocoa plantation than the two other habitats types. There is no statistical difference in bat species richness between primary forest and cocoa plantation (X2 = 0.29; ddl = 1; α > 0.5) and between secondary forest and human habitation (X2 = 0.22; ddl = 1; α > 0.5). The dominance was the inverse of the species richness in different habitat types in the Dja Biosphere Reserve (Figure 4). There is no statistical difference in bat species dominance between primary forest and cocoa plantation (X2 = 0.31; ddl = 1; α > 0.5), but between secondary forest and primary forest, there is a statistical difference (X2 = 5.04; ddl = 1; α < 0.025). Looking at the respective rank-abundance distributions, these differences were apparent (Figure 5).

Variation of species richness and dominance across habitat types. a: Comparison of bats species richness in cocoa plantation, human habitation. Primary forest and secondary forest. b: Comparison of bats dominance. Rarefied to equal abundances in cocoa plantation, human habitation, primary forest and secondary forest.
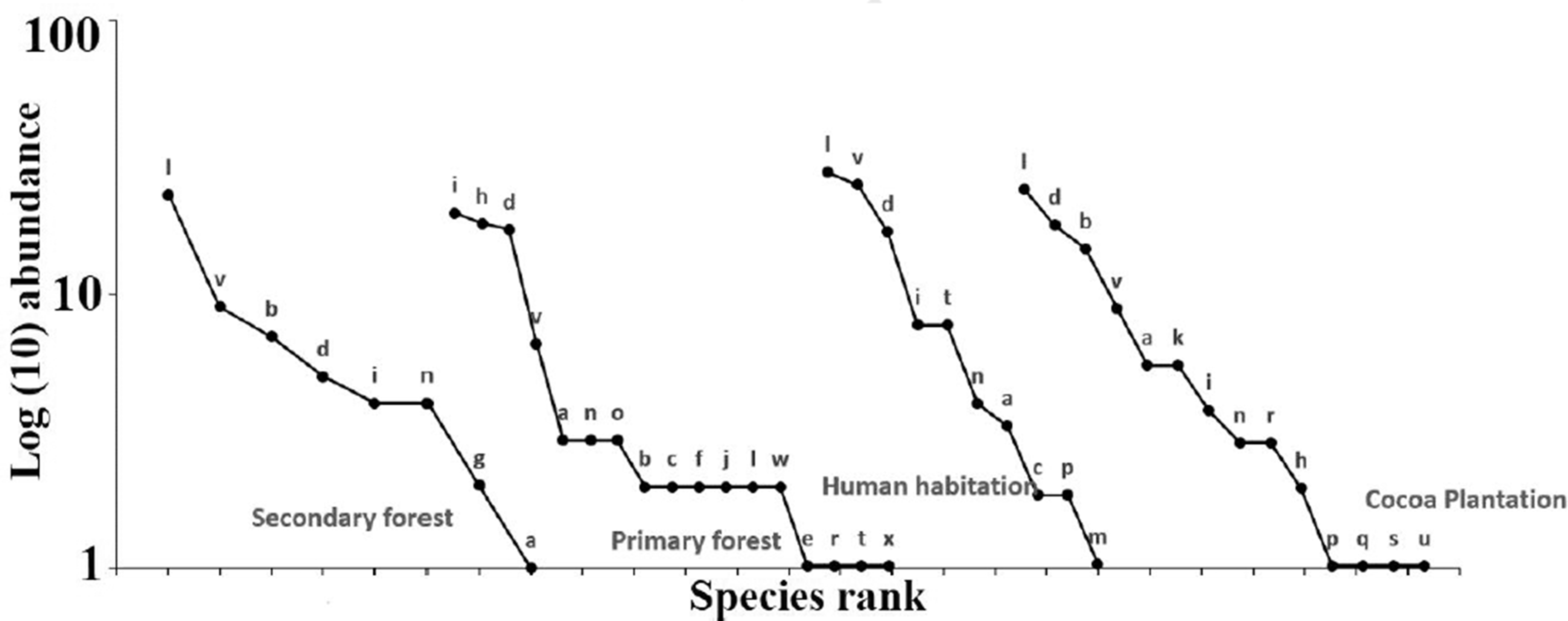
Rank-Abundance (Dominant Diversity) Curves for Each Habitat Showing Each Species Position Within the Assemblage in the Dja Biosphere Reserve (Cameroon). Letters represent the species captured. a: Casinycteris argynis. b: Dorirhyna cyclops. c: Epomops buettikoferi. d: Epomops franqueti. e: Glauconycteris albogutata. f: Glauconycteris argentata. g: Glauconycteris sp. h: Hipposideros caffer. i: Hipposideros ruber. j: Hypsignathus monstrosus. k: Macronycteris gigas. l: Megaloglossus woermanni. m: Myonycteris angolensis. n: Myonycteris torquata. o: Myotis bocagei. p: Neoromicia nana. q: Nycteris grandis. r: Nycteris hispida. s: Nycteris thebaica. t: Pipistrellus nanulus. u: Pipistrellus sp. v: Rousettus aegyptiacus. w: Scotoecus hirundo and x: Pipistrellus capensis.
Bats Biotic Dissimilarity Among Habitat Types
Differences in overall species composition were statistically significant among habitat types (ANOSIM, R = 0.12, P = 0.018). The hierarchical clustering also revealed dissimilarity between the four habitats types (Figure 6). SIMPER revealed that the species contributing most to dissimilarities between primary forest and secondary forest were H. ruber (19.03%), M. woermanni (14.85%), E. franqueti (13.31%), R. aegyptiacus (12.3%), and H. caffer (11.19%). Epomops franqueti (18.37%), M. woermanni (16.33%), H. ruber (13.48%), D. cyclops (10.24%) and H. caffer (10%), contributed to the dissimilarities between primary forests and cocoa plantations (Table 2). Megaloglossus woermanni (20.18%), R. aegyptiacus (19.78%), E. franqueti (14.39%) and H. ruber (13.65%), contributed most to the dissimilarities between primary forest and human habitations (Table 2). Megaloglossus woermanni (26.6%), E. franqueti (19.24%), R. aegyptiacus (14.34%) and D. cyclops (12.68%) contributed to the dissimilarities between cocoa plantations and secondary forest (Table 2). Megaloglossus woermanni (24.44%), R. aegyptiacus (21.15%), E. franqueti (15%) and D. cyclops (9.33%) contributed to the dissimilarities between cocoa plantations and human habitations. Megaloglossus woermanni (28.8%), R. aegyptiacus (24.45%) and E. franqueti (12.94%) contributed to the dissimilarities between secondary forest and human habitation (Table 2). GLMM analyses revealed that the distribution of the six most abundant species (E. franqueti, M. woermanni, R. aegyptiacus, D. cyclops, H. cf. caffer and H. cf. ruber) was influenced by the four habitat types (Table 3). The LSD post hoc test revealed a significant statistical difference between primary forest and cocoa plantation (P = 0.054).

Hierarchical Clustering Based on Bray-Curtis Distance Showing the Dissimilarity of Bat Assemblages Between Four Habitat Types in Dja Biosphere Reserve (Cameroon). Red numbers are AU p-values and black numbers are clusters.
Results of the Similarity Percentage Analysis (Using Simper), Showing the Importance of Dissimilarity for Different Bat Species in the Different Habitat Types Based on Capture Data in the Dja Biosphere Reserve.

Results of Generalized Linear Mixed-Effects Models Examining the Influence of Habitat Type on Bat Species Abundances in the Dja Biosphere Reserve. Values in bold indicate statistically significant differencies.

Discussion
To the best of our knowledge, studies that assess bat community response to forest loss and land conversion are lacking in Cameroon, though numerous studies have been conducted on the Chiroptera fauna of Cameroon in the past decade (eg Atagana et al., 2018; Bakwo Fils, 2009, 2010a, 2010b, 2012, 2014, 2018; Mongombe et al., 2019; Waghiiwimbom, Moise, Jules, & Tamesse, 2019a; Waghiiwimbom, Moise, Jules, Aimé, et al., 2019b). This study is the first to assess how bat species respond to forest loss and land conversion. The higher abundances of frugivorous bats in disturbed habitats can be attributed to proximity of fruit garden in which food such as Mangifera indica, Psidium guajava, Carica papaya, Annona muricata, Cola acuminate, and Garcinia kola were found. This finding is in agreement with other studies that showed that frugivorous bats diversity can increase in disturbed vegetation (Bobrowiec & Gribel, 2010; Weber et al., 2009). For example in rubber and oil palm plantations (Danielsen & Heegaard, 1995), eucalyptus plantations (Sonia et al., 2013), and complex plantation-forest landscapes (Katrin et al., 2015) abundances of frugivorous bats were higher than other guilds. This finding also corroborates with the works of Mongombe et al. (2019), Webala et al. (2019) and Avila-Cabadilla et al. (2009) that showed that disturbed habitats contained the greatest diversity of pteropodids. Our results indicate that habitat types resulting from forest disturbance can influence the abundance and composition of bats (Cortés-Delgado & Pérez-Torres, 2011).
When the various habitat types are considered, the species rarefaction curves of bat species nearly reach asymptotes for human habitations and secondary forest. This means that nearly all species were recorded in these two habitat types. According to Chao 2 species richness estimator, a single species could still be captured for both human habitations and secondary forests respectively, with additional sampling efforts. The fitted rarefaction curves for primary forests and cocoa plantations did not reach asymptotes, suggesting that not all species were recorded during this study. Approximately ten and six species were not sampled respectively in primary forests and cocoa plantations based on Chao 2 species richness estimator. According to Townsend et al. (2006), only a portion of organisms present in an area can be sampled. According to Chao et al. (2005) and Fahr et al. (2003), more sampling efforts and longer periods of sampling are highly recommended to achieve a complete inventory.
We used only ground-level mist nets to sample Dja Biosphere Reserve diverse bat fauna. This method is less efficient at capturing aerial insectivorous bats as well as bats from the families of Emballonuridae, Molossidae and some Vespertilionidae (Freeman, 1981, Portfors et al., 2000). Indeed, although acoustic monitoring requires an exhaustive and validated call library (Webala et al., 2019), many insectivorous forest bats commonly use their echolocation calls to avoid mist-nets and might be better sampled via acoustic monitoring and harp-trap (O’Farrell & Gannon, 1999). Thereby, an increase in sampling efforts using a variety of capture methods can possibly lead to the capture of more species (Colwell et al., 2004). This can explain the absence of some species (Eidolon helvum, Scotonycteris zenkeri, Hypposideros beatus, Hipposideros fuliginosus, Hipposideros curtus, Nycteris arge, Nycteris major, Mimetillus moloneyi, Tadarida thersites, Rhinolophus alcyone, Rhinolophus landeri, Saccolaimus peli) or families (Rhinolophidae and Molossidae) previously reported in the reserve (Bakwo Fils, 2009). Indeed, in addition to location and season, sampling method influences species-level detectability (Meyer et al., 2011). Even though we might have failed to capture some bats in our study, the method used is repeatable and would have introduced no systematic bias in our comparisons (Meyer et al., 2015).
When compared to previous studies of bats in the Dja Biosphere reserve, our study recorded more individuals (413) and fewer species (24) species in 126 capture nights than the 272 individuals and 29 species in 84 capture nights recorded by Bakwo Fils (2009). The difference can be attributed to rapid expansion of the agro-ecosystems in and around the biosphere reserve. Indeed, the modification of the natural forest for agricultural purposes is considered to be the main cause of biodiversity loss worldwide (Ramalho et al., 2014). Deforestation in the area is further compounded by the establishment of plantations in areas adjacent to Dja reserve encouraged by NGO’s as a means to discourage poaching within the reserve.
During our surveys, M. woermanni was the most abundant species recorded (105 individuals). It was also recorded in all the four habitat types sampled in the Dja biosphere Reserve. These findings are similar to those obtained by Bakwo Fils (2009) who noted the presence of this species in multiple habitats in the Dja biosphere reserve. This suggests that the species has a broader habitat niche and is likely to be less sensitive to habitat disturbances at the study site (Cosson et al., 1999). The number of individuals of this species recorded during our study (105) is significantly higher than the 66 individuals recorded by Bakwo Fils (2009) in the same area. These findings corroborate those of Happold (2013) that showed that this species is rare in collections but abundant in selected localities where food resources are abundant.
The three most abundant captured species were M. woermanni, E. franqueti and R. aegyptiacus. The high number of these frugivorous bats species within these habitats underlines their role in forest regeneration. Even though frugivorous bats can benefit from the conversion of primary forest to non-forest land-uses, they may also suffer when their habitats are drastically modified (Evans et al., 2018, Jones et al., 2001). This may probably be the reason for the low number of these species recorded in the secondary forest during the study period. Other studies reported more frugivorous bat species in disturbed areas than in primary forest (Rocha et al., 2017, Webala et al., 2019). In contrast, insectivorous bats preferred primary forest to disturbed habitats (Garcia-Morales et al., 2013). In this study, H. cf. ruber and H. cf. caffer were the most abundant species in primary forest. This latter record is consistent with previous findings of Waghiiwimbom et al. (2019a), who recorded more individuals of H. cf. ruber in primary habitats. This further corroborates with previous reports that revealed that H. cf. ruber is a cluttered space forager (Monadjem et al. 2010b), and supports the hypothesis that the forest offers a variety of roosting resources and hunting sites for prey (Erika de la Pena et al., 2015, Gorresen & Willig, 2004). Also, we netted two individuals of N. nana in human habitations. This is in agreement with previous findings of Waghiiwimbom et al. (2019a), who revealed that this species used uncompleted building as a night roost during night foraging bouts.
Our study found a difference in bat species composition and assemblages amongst the four habitat types sampled in the Dja Biosphere Reserve. Bat species richness was higher in the primary forest (17), than in cocoa plantations (14), human habitations (10) and then secondary forest (8).This is expected as modification of the primary forest through the establishment of plantations, man-made structures like buildings and logging would influence bat species composition. This result contrasts with the findings of Montaño-Centellas et al. (2015) in Tropical Andes which showed that bat species were more abundant in urban areas followed by plantations, primary forests and secondary forests. This difference can be explained by the fact that, their study was on phyllostomid bat species, which prefer anthropogenic habitats, particularly agro-forestry systems (Garcia-Morales et al., 2013). However, contrary to species richness, species dominance was highest in secondary forest followed by human habitations, cocoa plantations and primary forests. Generally, the dominance was low in three of the four habitat types, showing that bat population of the Dja Biosphere Reserve are co-dominated by several species. The higher dominance in secondary forest might be the result of habitat loss and interspecific competition. Indeed, the dramatic decline in resources availability, as well as the global loss or alteration of foraging habitat, may generate new competitive interactions or exacerbate existing interactions in the Anthropocene, and into the future (Salinas-Ramos et al., 2020).
Our result suggests that four of the six most abundant species (M. woermanni, R. aegyptiacus, H. cf. caffer and H. cf. ruber) generally responded to habitat conversion, despite their morphological and ecological differences with regards to food, roosting sites and foraging strategies. Frugivorous bats (M. woermanni and R. aegyptiacus) responded positively to habitat conversion while, insectivorous (H. cf. caffer and H. cf. ruber) responded negatively. Thereby the poor species richness in secondary forests and human habitations may be due to the severity of anthropogenic disturbance in these two habitat types, lack of appropriate food resources and less complex vegetation compared to primary forest and plantations (Webala et al., 2019). In fact, a greater number of species (9) out of 24 (37.5%) were shared between primary forest and cocoa plantations. Despite an increased number of shared species between primary forest and cocoa plantation, the cluster analysis revealed a dissimilarity between the two habitats. This can be explain by the fact that many species play some part in determining the dissimilarity between the two groups, and this is typical of such analyses (Clarke, 1993). In this tightly clustered situation, it is no surprise to find that the principal contributions come from species that are abundant in one habitat and rare (though not necessarily totally absent) in the other; the balance of contributions in this case is from species that are numerous in one habitat but rare in the other such as H. cf. ruber and H. cf. caffer which are abundant in primary forest (22 and 20 individuals respectively) and rare in cocoa plantation (four and two individuals respectively). Indeed, the complexity and nature of habitats constitute a valuable factor that determines the species composition and diversity in a particular area (Bellanthudawa et al., 2019, Casas et al., 2016). Some species, like the hipposiderids (Doryrhina cyclops, H. cf caffer, H. cf ruber), are clutter specialists (Norberg & Rayner, 1987), and are adapted for foraging close to or within dense vegetation (Kingston et al., 2003). Species that had the lowest abundances are sensitive to disturbance and therefore face an increased threat of local extinction (Henle et al., 2004). Cocoa plantations have gained recognition as a refuge for biodiversity within agro-ecosystems (Schroth, 2004, Cassano et al., 2009, Rice & Greenberg, 2000, Schroth & Harvey, 2007), but their ability to retain forest-dependent native species as well as to have higher species diversity is dependent on their distance to native forest patches of considerable size (Faria & Baumgarten, 2007).
Implication for Conservation
Although secondary forest can sustain a high species richness of bats, the transformation of primary vegetation into disturbed habitats seems to have a large effect on assemblage composition and abundance of some bat species. Indeed management and conservation efforts should first and foremost focus on preserving the four habitat types (primary forest, secondary forest, cocoa plantation and human habitation) already present due to their greater contribution of habitat type in explaining bat abundance responses to fragmentation. This is of critical relevance in landscapes where there is contrast between primary forest and disturbed habitats (e.g., more heterogeneous landscapes or landscapes with higher anthropogenic pressures) due to the expected increasing contribution of habitat structure characteristics. Because of the rapid conversion of rain forest into agricultural land and secondary forest, caused by deforestation rate of 0.5%/year, secondary forests will occupy a high percentage of the total forested area in the world in the next decades (Asner et al., 2009, FAO, 2010). This is not a good news for bats of southern Cameroon particularly those of the Dja Biosphere Reserve. Indeed, in this protected area, secondary forest appears to be more species poor than the other three habitat types studied. Secondary forest can present more food resources and roosts to bats than cocoa plantation and human habitation, but, as indicated in our discussion, this poor species composition is probably the consequence of human’s activities as well as wood exploitation, machines noises; because it is well known that bat are very sensitive to disturbance of their habitats. One factor that can help to boost the species richness and abundance of bats in human habitation is the maintenance of fruits trees, as they can act as attractor for bats, probably representing important food sources.
To minimize adverse effects of habitat disturbances on the sensitive bats communities, conservation and management efforts should be more oriented toward protecting habitat and resources of these key ecological mammal groups during the modification of available land uses. The exploitation of the woods around the Dja Biosphere Reserve should be reduced or stopped if possible; exploitation of primary forest should be carried out using few roads and as little mechanized equipment as possible to limit the degree of habitat disturbance (Tchoumbou et al., 2020). Awareness raising campaigns of local communities living in neighbourhoods of the Dja Biosphere Reserve on the importance of bats and forest conservation should be planned to reduce anthropogenic effects on primary forest and biodiversity. Government needs also to implement better policies to control and regulate forest exploitation.
Measurements of the Different Parameters of the Species Caught in the Dja Biosphere Reserve.
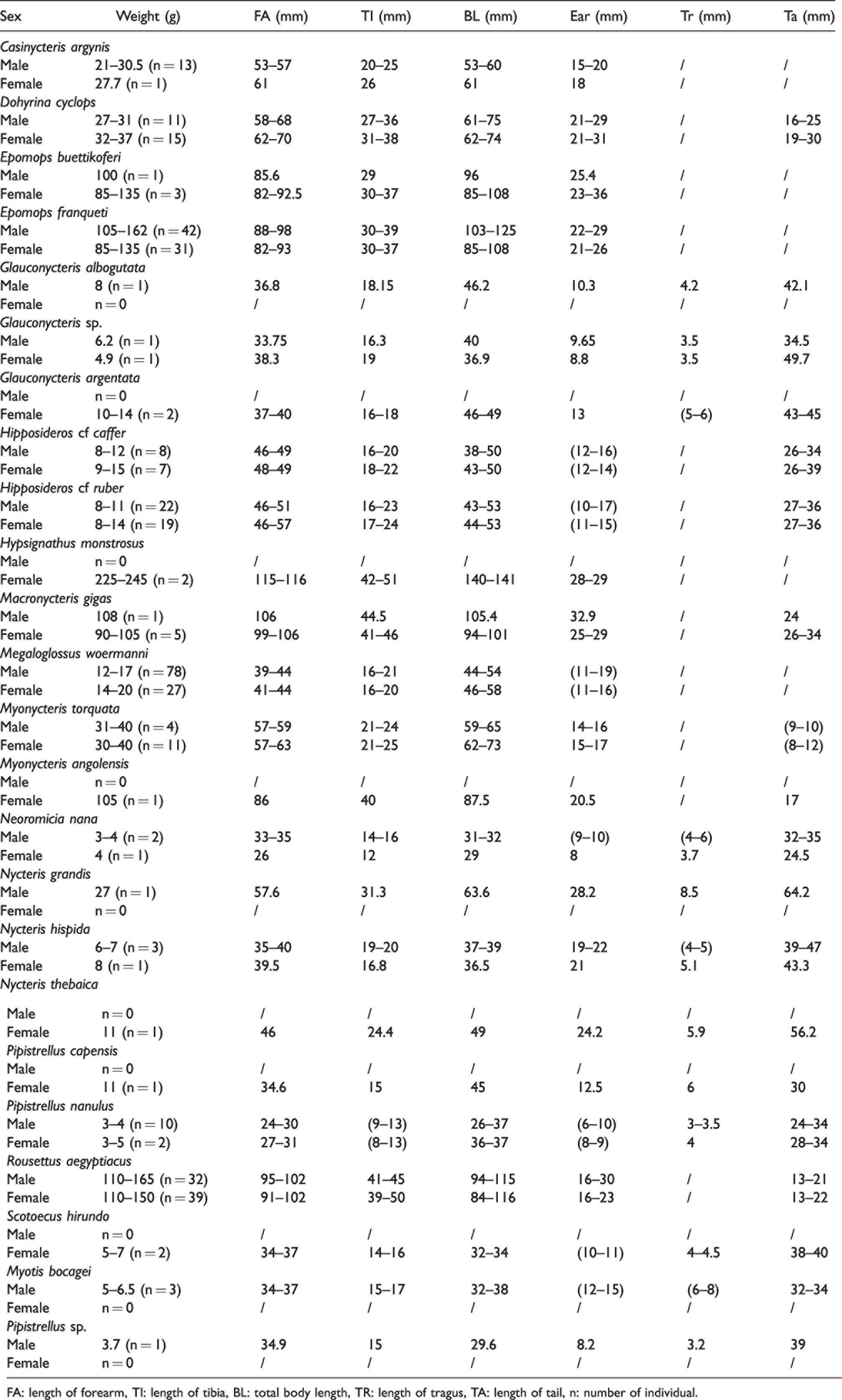
FA: length of forearm, TI: length of tibia, BL: total body length, TR: length of tragus, TA: length of tail, n: number of individual.
Bat Roosts Sites Present in the Dja Biosphere Reserve.

Appendix A
Appendix B
Footnotes
Acknowledgments
The authors are grateful for the collaboration, cooperation and support of Ernest Seamark and Jon Flanders, who facilitated the completion of this work. The authors extend their gratitude to the Minister of Scientific Research and Innovation of Cameroon for providing the research permits for this project. Special thanks also goes to the conservator and the conservation service of the Dja Biosphere Reserve, for granting access to this protected area. We are also grateful to Takwo Jean Michel for providing some field work materials support. Finally, we would like to thank villagers of all the villages around the Dja Biosphere Reserve for their hospitality.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study received financial support from Bat Conservation International scholarship 2017 (APJ) (grant number SS 1808) and also from Rufford Small Grant (APJ) (Grant number 25283–1).