
Abstract
Changes in wetland environments can alter the dynamics of waterbird populations. We investigated the effects of hydrological and landscape variables on the abundance of resident waterbirds (ducks, fish-eaters, large waders, small waders, and vegetation gleaners) from 2003 to 2014 in Bung Boraphet, Thailand's largest freshwater wetland. Generalized linear mixed models were used to determine the effects of environmental variables on waterbird numbers, and generalized additive mixed models were used to identify the threshold for each effect. The results revealed that the population of all waterbirds declined by 27% from 2003 to 2014 with highest decline of 56% in ducks. Increasing water depth was negatively correlated with the abundance of small waders and vegetation gleaners. Higher concentrations of dissolved oxygen in the water increased the abundance of fish-eaters, while marshy areas were positively associated with the abundance of ducks, large waders, and vegetation gleaners. The abundance of fish-eaters, large waders, and small waders were negatively associated with the spatial area of waterbodies. Expanding human settlements decreased the abundance of vegetation gleaners, while vegetation infested by Mimosa pigra decreased the abundance of large waders. The study concludes that the maintenance of an optimal water depth and quality, preservation of critical marsh habitats, eradication of invasive species, and restrictions on new human settlements adjacent to wetlands are all necessary to conserve resident waterbird populations. Validating these findings in additional research sites is recommended before applying it to a broader landscape level.
Keywords
Introduction
Waterbird populations are declining globally (Hansen, Menkhorst, Moloney, & Loyn, 2015; Z. Ma et al., 2014; W. Wang, Fraser, & Chen, 2017). The decline is particularly alarming in Asia where 50% of known populations were found to be in decline (Wetlands International, 2012). Waterbirds, an important component of wetland ecosystems, are sensitive to changes in the wetland environment; they either disperse or aggregate in response to such changes (Brandolin & Blendinger, 2016; Henry & Cumming, 2016). However, in some regions, the environmental variables affecting waterbird abundance are not yet clearly understood due to the complexity of the wetland environmental systems and differences in the composition and structure of avian communities.
Several studies have demonstrated that environmental variables such as wetland hydrology and landscape can alter waterbird abundance in different ways (Alexander & Hepp, 2014; Brandolin & Blendinger, 2016; Tavares, Guadagnin, de Moura, Siciliano, & Merico, 2015; Wen, Rogers, Saintilan, & Ling, 2011). For example, fluctuations in water depth influence the physical condition of waterbird habitats (e.g., the establishment of mud-flats), affect food availability, and prey vulnerability (Bellio & Kingsford, 2013; Lantz, Gawlik, & Cook, 2011; Timmermans, Badzinski, & Ingram, 2008; Y. Wang et al., 2013; Zhang et al., 2016), which are characteristics that define habitat use by waterbirds (Jedlikowski, Chibowski, Karasek, & Brambilla, 2016). Likewise, the conversion of wetlands into agricultural areas and human settlements poses a significant threat to waterbirds. Other than direct habitat loss, such change in the wetland landscape also affects hydrological systems (Harrison & Whitehouse, 2012), and consequently, habitat use by the waterbirds (Cintra, 2015; Tavares et al., 2015). Therefore, understanding how different environmental variables affect waterbirds is important for the effective conservation of waterbirds and the management of their habitats. However, as different functional groups of waterbird respond differently to changes in environmental variables due to of varying habitat requirements, the relationship between waterbird abundance and environmental variables is complicated (Tavares et al., 2015). For instance, low concentrations of dissolved oxygen may affect waterbird species that use lake resources for foraging such as fish-eaters (Sulai et al., 2015), whereas similar changes may not affect species that use waterbodies solely as daytime roosts such as ducks. Some landscapes such as paddy fields may cause an increase in waterbird species that prefer shallow water for foraging (Acosta et al., 2010), but they are not a preferred habitat for the larger and more sensitive waterbird species like Black-headed Ibis Threskiornis melanocephalus (Sundar, 2006).
Bung Boraphet (BBP) in Central Thailand, a wetland of international importance proposed as a Ramsar site, provides an important habitat for both resident and migratory waterbirds (Office of Environmental Policy Planning, 2002). It is reported that BBP is under threat and increasing pressure due to hydrological and landscape changes such as water shortage, invasion of Mimosa pigra, and an increase in the area of land converted for agricultural use and human settlements (Chaichana & Choowaew, 2013; Sriwongsitanon, Surakit, & Hawkins, 2009). However, little research has been conducted on waterbirds and their environmental associations in BBP. There has been one study on the nest-site selection of a breeding visitor, Oriental Pratincole Glareola maldivarum, by Chaiyarat and Eiam-Ampai (2014), but no recent studies were found on resident waterbirds. Resident waterbirds might be at greater risk due to the rapidly changing local landscape and hydrology as they select a particular territory for a lifetime (West, Goss-Custard, dit Durell, & Stillman, 2005). Undesirable environmental changes reduce habitat availability (Orians & Wittenberger, 1991) and when such changes go beyond certain limits, they can lead to extinction of local populations (Peters & Otis, 2006). Therefore, a better understanding of the effects of changing hydrology and landscapes on the population trends and seasonal fluctuations of different functional groups of resident waterbirds would improve our in-depth knowledge of habitat use and enable the implementation of effective conservation measures.
This study was designed to investigate the effects of hydrological and landscape variables on waterbird abundance in BBP. To achieve this objective, we (a) assessed the long-term trends and seasonal variations in waterbird abundance in BBP from 2003 to 2014, (b) studied hydrological and land-use changes in BBP, and (c) examined the effect of hydrological and landscape variables on the abundance of resident waterbirds. Specifically, we tested the hypothesis that each of the five functional groups of resident waterbirds (ducks, fish-eaters, large waders, small waders, and vegetation gleaners) would show specific responses to changes in four hydrological variables (dissolved oxygen levels in the water, turbidity, water depth, and water temperature) and eight landscape variables (cover of fish farms, human settlements, landfills, marshes, mixed crops, paddy fields, vegetation infested by Mimosa pigra, and waterbodies). Finally, we proposed recommendations for the conservation of resident waterbirds and their habitats in BBP.
Methods
Study Site
This study was conducted in Bung Boraphet Wetland (BBP), Nakhon Sawan Province, Central Thailand (between latitude 15 °40'N and 15 °45'N and longitude 100 °10'E and 100 °23'E; Figure 1). BBP is the largest freshwater wetland on the floodplain of the Chao Phraya River, with a total area of approximately 212 km2. It was developed in 1927 by the damming of a freshwater swamp to enhance fisheries (Sriwongsitanon et al., 2009). In 1937, this area was designated as a “Conservation Zone” to prevent human encroachment and settlement in the area. Nevertheless, by 2015, more than 32,000 people were living inside that zone (Department of Provincial Administration, 2015) and were making use of its natural resources such as fish and wetland vegetation. In addition, BBP has been used for domestic and agricultural water supply and flood control. Incidents of direct waste disposal from the surrounding human settlements were also observed during the study period, which indicated that BBP was also used for waste disposal.
Location and map of the study area in Thailand. Five-kilometer buffer area surrounding the boundary of the lake, boat survey route used to count waterbird populations inside the lake, research station, and water sampling points are also shown. Photographs of the different waterbird species, wetland habitats, and threats/pressures in Bung Boraphet: (a) and (b) flocks of ducks and large waders; (c) vegetation dominated by Mimosa pigra; (d) fishermen harvesting wetland vegetation; and (e) to (h) waterbirds using different habitats (e.g., marshes and paddy fields). Photographs were taken by R. U. Haq.
BBP supports a wide variety of flora and fauna because of its distinct geographical location, its variety of landscapes, and the natural dry–wet weather cycles (Jintanugool & Round, 1989). So far, 23 species of aquatic macrophytes including emerged, submerged, and floating aquatic plants, 93 species of phytoplankton, 43 species of zooplankton, and 77 species of fish have been recorded in BBP (Chaichana & Choowaew, 2013). Additionally, BBP is an important bird area (BirdLife International, 2017) that supports large numbers of resident and migratory waterbirds; around 36,000 and 20,060 waterbirds were counted in 1991 and 1994, respectively (Lopez & Mundkur, 1997). Although BBP is very important for wild flora and fauna, it has never been managed for wildlife conservation.
BBP lies in a tropical monsoonal climate zone. The wet season is from May to October, with around 78% of the annual rainfall occurring during this period. From 2003 to 2014, mean annual rainfall of 1,240 mm was recorded, while the monthly temperature varied during this period from an average minimum of 20.9 ℃ to an average maximum of 36.7 ℃. According to the hydrological data from the Regional Environmental Office of Nakhon Sawan for the period of 2003 to 2014, the average water depth in BBP was 1.8 ± 0.7 m with a maximum water depth of 2.9 m recorded in 2011.
Waterbird Surveys
Summary of the Functional Groups of Resident Waterbirds in Bung Boraphet Wetland.

Note. CV = Coefficient of variation.
Hydrological and Landscape Variables
We considered four hydrological variables (dissolved oxygen, turbidity, water depth, and water temperature) and area of eight landscape variables (fish farms, human settlements, landfills, marshes, mixed crops, paddy fields, vegetation, and waterbodies) as predictors. Hydrological data were obtained from the Regional Environmental Office, Nakhon Sawan Province. The team from Regional Environmental Office recorded the water depth in meters at five points inside BBP (Figure 1). From the same points, dissolved oxygen levels (mg/l) and water temperature (℃) were measured in situ using a multimeter and turbidity (NTU) was determined using a black and white Secchi disc (Figure 1).
Summary of Explanatory Variables.
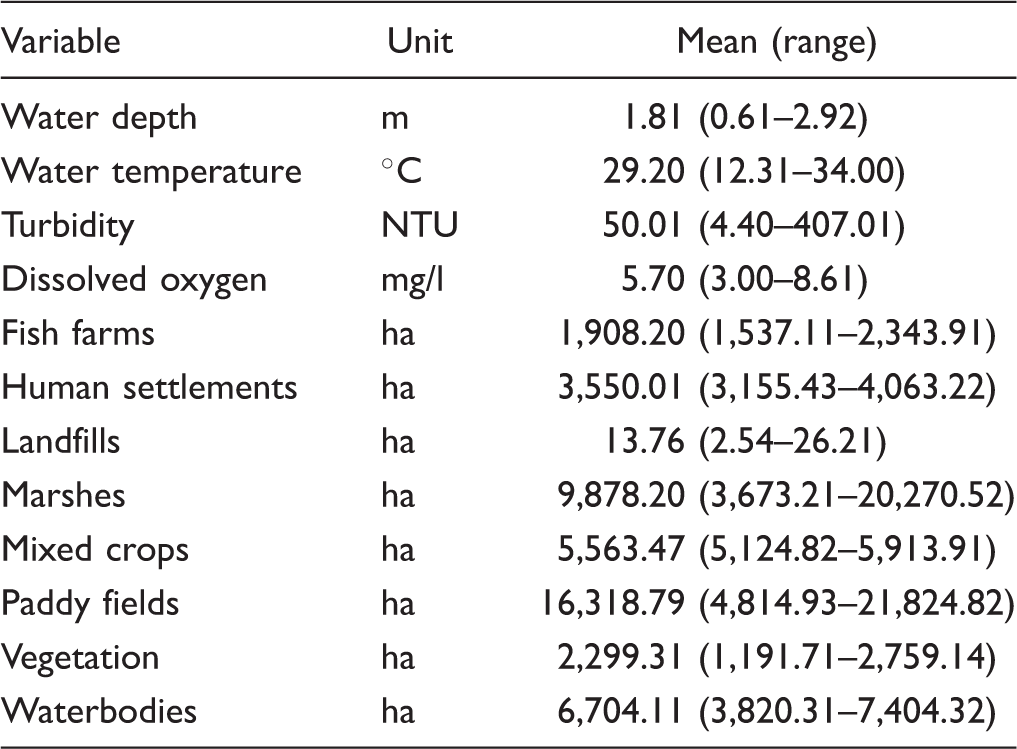
Note. NTU = Nephelometric turbidity unit; ha = hectares.
Statistical Analyses
To check the effect of predictor variables on waterbird abundance, we used generalized linear mixed models (GLMMs; Bolker et al., 2009) with a negative binomial distribution. Akaike's information criterion (AIC) was used for model selection (Burnham & Anderson, 2004); the best model is the one with the lowest AIC score. In some cases, where AIC scores for two models were not significantly different, we selected candidate models with cut-off criteria of ΔAIC<2. Model averaging was used to evaluate the coefficients of uncertain models and unconditional standard errors. We used an 85% confidence interval to recognize variables with a significant effect on waterbirds: This interim reduces model choice and makes parameter-assessment criteria more harmonious than the smaller interim widths, for example, 95% (Arnold, 2010). We used the sum of the highest number of waterbirds recorded from monthly surveys as a response variable for each functional group for GLMMs. Predictor variables were added in the models in the following sequence: (a) influence of each variable; (b) all variables in a group, for example, all hydrological variables; and (c) hydrological and landscape variables together. The variable “year” was included in all models to account for possible correlation of observations among survey years. The data for waterbirds were overdispersed, and there was high variance in the counts between the wet and dry seasons due to the difference in survey routes between both seasons which affects the survey effort. Consequently, we treated season as a random effect (intercept) in all models to account for the nuisance parameter. We checked for outliers and collinearity before performing model fits following the recommendations of Zuur, Ieno, Walker, Saveliev, and Smith (2009). We applied Pearson's correlation test to all predictor variables. Highly correlated variables (r > .5) were not used in the same model. We standardized the data to dimensionless units (Z-scores) by subtracting the mean for each taxon and dividing this by its standard deviation in order to make resulting models directly comparable.
We assessed the appropriate level of each significant variable (obtained from GLMMs) graphically by consolidating smoothing capacities into the generalized additive mixed models (GAMMs) following Tavares et al. (2015). Different values of knots (k) were tested to make effective degrees of freedom (edf) not much less than k−1 for each model following the guidelines of Wood (2006).
Statistical analysis was performed in R (version 3.3.3). We used package glmmADMB (Skaug, Fournier, Nielsen, Magnusson, & Bolker, 2013) for GLMMs; wiqid (Meredith, 2016) for model selection; MuMIn (Barton, 2016) for model averaging; mgcv (Wood, 2006) for producing the response curves through GAMMs; and ggplot2 (Wickham, 2009) for creating plots.
Results
Waterbird Abundance and Species Composition
From 2003 to 2014, 35 species of resident waterbirds from 15 waterbird families were observed in BBP. Of these, 28 species (2 species of ducks, 13 species of fish-eaters, 3 species of large waders, 5 species of small waders, and 5 species of vegetation gleaners) were considered for further analysis based on the sufficiency of available records. The total population of the studied species changed every year with a mean value of 25,300 over a 12-year period.
Overall, resident waterbirds declined by 27% from 2003 to 2014. Ducks showed a long-term decline of 56%, particularly 56% for the Lesser Whistling-duck Dendrocygna javanica and 67% for Cotton Pygmy-goose Nettapus coromandelianus. Large waders declined by 25%, with the Asian Openbill Anastomus oscitans declining by 36%. Vegetation gleaners declined by approximately 8%. The populations of fish-eaters and small waders showed seasonal fluctuations, but overall populations of these functional groups remained stable throughout the study period. Moreover, there were clear wet–dry seasonal patterns for the ducks. The other four functional groups showed no regular seasonal patterns, although there appears to be a considerable year to year fluctuation (Figure 2).
Annual mean waterbird abundance of five functional groups in Bung Boraphet Wetland during the dry and the wet season (2003–2014). Line chart showing precipitation (in millimeters) is shown. Line chart of “marsh area in hectares” is also shown indicating its association with the functional group ducks as they showed highest long-term declines.
Among the functional groups, ducks and large waders were the most dominant (60% of the total population). Among the families of waterbirds, Anatidae (ducks) were the most abundant, making up around 43.5% of the total waterbird population, followed by Ciconiidae (storks) accounting for 28%. The Lesser Whistling-duck D. javanica and the Asian Openbill A. oscitans were the predominant species, accounting for 39% and 26% of the total species, respectively. According to the categorizations of the IUCN Red List, we recorded one Vulnerable species—the Lesser Adjutant Leptoptilos javanicus—and five Near Threatened waterbird species—the Great Thick-knee Esacus recurvirostris, the Black-headed Ibis Threskiornis melanocephalus, the Oriental Darter Anhinga melanogaster, the River Lapwing Vanellus duvaucelii, and the Spot-billed Pelican Pelecanus philippensis (IUCN, 2017). The species observed within each functional group along with their annual mean abundance and global threat statuses are reported in Appendix A.
Hydrological and Land-Use Change
From 2003 to 2014, there were clear differences in water depth between seasons in which average water depth was higher during the wet seasons. Similarly, for the majority of the years, average turbidity and dissolved oxygen were higher during the wet seasons. No long-term decrease or increase was observed in the hydrological variables. On the other hand, some landscape variables showed considerable changes in the area. Land-use analysis showed that paddy fields were the most dominant landscape feature covering 47.2% of the total study area in 2014. In 2007, there was a significant decrease in the area of paddy fields due to an increase in marsh areas. The area accounted for by waterbodies declined by 31% from 2001 to 2014 (a major decline was observed after 2012 in the waterbody of BBP). The area of vegetation—dominated by Mimosa pigra—increased by 81% from 2001 to 2014. Both human settlements and fish farms showed an increase of 27% each. Marsh areas changed a little, except in 2007. After 2007, there was a decline till 2012. Also, there were changes in their distribution. The total areas accounted for by mixed crops and landfills exhibited minimal changes from 2001 to 2014. Land-use classification maps for the years 2001, 2007, 2009, 2012, and 2014 are shown in Figure 3. Seasonal variations and long-term trends in the hydrological and landscape variables from 2003 to 2014 are shown in Appendix B.
Land-use classification maps of the study area for Years 2001, 2007, 2009, 2012, and 2014.
Effects of Hydrological and Landscape Variables on Waterbird Abundance
List of the Best Generalized Linear Mixed Models (GLMMs) Explaining the Abundance of Resident Waterbirds in Bung Boraphet Wetland.

Note. K = Number of parameters estimates + intercept; AIC = Akaike's Information Criterion; Δ AIC = Difference in AIC score between ranked models; w = Model weight. Predictor variables occurring in the model sets are as follows: (a) marshes (marsh areas in hectares with emergent and floating wetland vegetation), (b) vegetation (natural vegetation in hectares including grasslands, shrubs, trees, and the invasive Mimosa pigra), (c) waterbodies (areas of open water in hectares without any vegetation), (d) human settlements (built-up or developed areas in hectares), (e) paddy fields (rice fields in hectares), (f) dissolved oxygen levels (mg/l), and (g) water depth in meters (m).
Average Estimates of the Coefficients, Unconditional Standard Errors, and Confidence Intervals of the Variables Present in the Best Generalized Linear Mixed Models (GLMMs).

Note. Uncond. SE = Unconditional standard error. Predictor variables occurring in the model sets are as follows: (a) marshes (marsh areas in hectares with emergent and floating wetland vegetation), (b) vegetation (natural vegetation in hectares including grasslands, shrubs, trees, and the invasive Mimosa pigra), (c) waterbodies (areas of open water in hectares without any vegetation), (d) human settlements (built-up/developed areas in hectares), (e) paddy fields (rice fields in hectares), (f) dissolved oxygen levels (mg/l), and (g) water depth in meters (m).
GAMMs illustrated that fish-eaters were found in greater abundance when the level of dissolved oxygen in the water exceeded 5.5 mg/l. The appropriate water depth range for small waders and vegetation gleaners appeared to be less than 1.8 m; these waterbird groups showed a decrease in abundance after average water depths exceeded 1.8 m. Ducks, large waders, and vegetation gleaners showed a positive trend when marsh areas exceeded 6,500 ha, 6,700 ha, and 10,000 ha, respectively. Fish-eaters and large waders decreased in abundance after the spatial area of waterbodies exceeded 6,700 ha, while vegetation gleaners decreased in numbers when the area of waterbodies exceeded 7,050 ha. Large waders decreased when the total vegetation area exceeded 2,300 ha. Vegetation gleaners also showed a decline in abundance when the area of human settlements around the lake exceeded 3,520 ha. Figure 4 illustrates response curves as function of the significant variables for each functional group.
Waterbird abundance responses to significant variables obtained using generalized additive mixed models. Shaded areas show 95% while circles indicate partial residuals. Blue-shaded graphs indicate hydrological variables, and green-shaded graphs indicate landscape variables.
Discussion
Our study detected a decline of 27% in the population of resident waterbirds from 2003 to 2014. Multiple studies have noted the declining numbers of waterbirds in Asia and Thailand (Mundkur, Langendoen, & Watkins, 2017; Wetlands International, 2010, 2012). The lowest abundance in overall waterbird population was observed in 2011, followed by a partial recovery from 2012 to 2014 (Figure 2). This trend could be due to a major flooding event in 2011 which overtopped the capacity of BBP. Waterbirds are likely to disperse as an immediate response to flooding; however, they return and reassemble on the main wetland area again as the flooded areas decline in extent (Poiani, 2006).
A long-term decline was observed in the populations of D. javanica and N. coromandelianus. The fifth edition of the waterbird population estimates (Wetlands International, 2012) reported a decreasing trend in the population of D. javanica and an unknown population status of N. coromandelianus in Asia. The coordinated counts of waterbirds in Asia and Australasia showed a decline of 24% in the population of D. javanica from 2008 to 2015 in Asia, but no change was observed in the population of N. coromandelianus (Mundkur et al., 2017). However, as the survey numbers fluctuated wildly from year to year between 2008 and 2015, the uncertainty across the population trends for this report is significant. Therefore, in the present study, we hypothesize that the decline in the population of ducks is associated with the changes in marsh area (Figure 2). The 25% decline in the population of large waders (primarily A. oscitans) was due to the outbreak of avian flu in 2004, which wiped out breeding colonies (Siengsanan et al., 2009; Suksatu et al., 2009). The birds that recovered from this outbreak were subsequently affected by extensive flooding in 2011, which may have either caused a population crash or reduced the availability of their principal prey Applesnail Pomacea canaliculata (Philip Round, personal communication, April 25, 2017).
Nevertheless, ducks and large waders were the dominant functional groups as BBP still offers an excellent roosting site for ducks, while the surrounding paddy fields also act as feeding grounds for ducks and large waders (Chaiyarat, Sookjam, Eiam-Ampai, & Damrongphol, 2015). For the smaller and less conspicuous species of waterbirds (e.g., small waders and vegetation gleaners), detectability differences might play a major role, particularly during the rainy season. In addition, dense wetland vegetation preferred by vegetation gleaners might also reduce their detectability.
The land-use analysis suggested that there were significant changes in the areas of waterbodies, vegetation, human settlements, and fish farms from 2001 to 2014. The total area of the waterbodies showed a decline of 31% from 2001 to 2014. Classified land-use maps showed a conversion of some waterbodies into human settlements and paddy fields (Figure 3). Rapidly expanding human settlements, unsustainable land-use practices (Chaichana & Choowaew, 2013; Sriwongsitanon, Surakit, Hawkins, & Chandrasena, 2007), and unregulated pumping of water from BBP for the cultivation of dry-season rice (Sriwongsitanon et al., 2009) were likely the main reasons for this decrease. The rapid increase in vegetation cover was due to the invasion of Mimosa pigra. In suitable habitats, where water aids seed dispersal, mimosa can out-compete local vegetation (Rijal & Cochard, 2016). Significant changes in the marsh area and paddy fields were observed in 2007. These changes were caused by two small flooding events, which transformed the paddy fields into marsh areas due to water retention. In addition, there were changes in the distribution of the marshy areas due to the shrinkage of the total waterbody area of BBP and the expansion of the paddy fields. The long-term trends and seasonal variations in the waterbird populations, decline in area of some landscape variables, and fluctuations in hydrological variables between dry and wet season appear to indicate that both hydrological and landscape variables are affecting waterbirds in BBP.
GLMMs results suggested that the assemblage of each functional group of waterbirds was influenced by a distinct set of hydrological and landscape variables. Groups of waterbirds showing seasonal fluctuations but a stable population (i.e., fish-eaters and small waders) were governed by seasonally fluctuating hydrological variables, while groups showing long-term trends (i.e., ducks, large waders, and vegetation gleaners) were governed by the changes in landscape variables. Increases in water depth caused the abundance of small waders and vegetation gleaners to decrease. Appropriate level of water is very important as it dictates the feeding success for small waders (Lantz et al., 2011) and vegetation gleaners (Zhang et al., 2016). Small waders usually prefer shallow waters because their small bills restrict the catching of prey beyond a certain water depth (Lantz, Gawlik, & Cook, 2010). Shallow waters also favor macrophyte growth (Zhang et al., 2016), which serves as a nesting and feeding habitat for vegetation gleaners, such as moorhens (Gallinula chloropus; Forman & Brain, 2004). Dissolved oxygen levels were positively associated with the abundance of fish-eaters. Sulai et al. (2015) observed a similar relationship between tropical waterbird species and dissolved oxygen levels in Peninsular Malaysia. Fish is the main food resource for fish-eaters, and sufficient concentrations of dissolved oxygen enhance fish survival (Williams et al., 2004). Greater availability of feeding resources and higher levels of dissolved oxygen were the reasons for this positive association.
Among the landscape variables, increase in the marsh area was positively associated with the abundance of ducks, large waders, and vegetation gleaners. MacDonald and Henderson (1977) explained that marshes attract more waterbird species compared with other landscapes (e.g., open water) by offering more shelter, an abundance of food, suitable nesting, and safe roosting sites. Ali, Ripley, and Dick (1987) and Upadhyaya and Saikia (2010) in their respective studies of the Cotton Pygmy-goose N. coromandelianus and the Lesser Whistling-duck D. javanica observed that both species prefer marshes compared with other habitats. In contrast to marsh areas, open water areas have higher predation risk and less food (Kleijn, Cherkaoui, Goedhart, van der Hout, & Lammertsma, 2014). As a result, increases in the spatial area of waterbodies showed negative effects on the abundance of fish-eaters, large waders, and small waders in this study. Changes in vegetation areas showed a negative effect on large waders. It is apparent that the invasion of Mimosa pigra has changed the vegetation structure so that it is no longer suitable for large waders. Mimosa pigra can transform tropical wetlands into monospecific stands that no longer support local birds and vegetation (Braithwaite, Lonsdale, & Estbergs, 1989). Moreover, waterbirds mostly rely on visual cues to detect prey, and complex vegetation may reduce prey accessibility and detectability (Pérez-García, Sebastián-González, Alexander, Sánchez-Zapata, & Botella, 2014). For example, the A. oscitans feeds majorly on the Applesnail P. canaliculata (Sawangproh, Round, & Poonswad, 2012), and vegetation reduces Applesnail's detectability by providing it with more hiding spaces (Meyer-Willerer & Santos-Soto, 2006). Human settlements showed negative effects on vegetation gleaners. Habitat degradation and competition between waterbirds and humans for the same resources are the likely reasons for this negative association. For example, water lilies provide excellent foraging and nesting habitat for vegetation gleaners. However, people around BBP also use and harvest water lilies as a food resource (Chaichana & Choowaew, 2013). The expansion of human settlements and increasing human populations inside the conservation zone of BBP are likely causing an increase in the harvesting of such resources, thus affecting vegetation gleaners.
Some important landscape variables, for example, fish farms and paddy fields, exhibited no significant effect on waterbird abundance in this study. However, studies from other regions have shown that paddy fields (Fasola, Canova, & Saino, 1996; Z. Ma, Cai, Li, & Chen, 2010) and fish farms (Feaga, Vilella, Kaminski, & Davis, 2015; Rocha, Ramos, Paredes, & Masero, 2017) can provide alternative habitats for waterbirds. We, therefore, call for further in-depth investigations so as to understand the relationships between the waterbird abundance and these variables with greater certainty. More specifically, a detailed study is needed to understand the effects of the invasive Mimosa pigra on waterbirds and wetland habitats in BBP.
In summary, our results support the proposed hypothesis that the effects of hydrological and landscape variables are specific to different functional groups of waterbirds. A strong association was observed between waterbird groups showing long-term declines (ducks, large waders, and vegetation gleaners) and landscape variables (marshes, waterbody, vegetation infested by Mimosa pigra, and human settlements), and an association was observed between waterbird groups showing stable populations (fish-eaters and small waders) and hydrological variables (water depth and dissolved oxygen). Water depth, dissolved oxygen levels, marshy areas, vegetation, waterbodies, and human settlements were critical variables for resident waterbirds in BBP.
Implications for Conservation
The study suggests that management should focus on both hydrological and landscape variables when implementing waterbird conservation and habitat restoration in freshwater wetlands. Our results also indicate that each functional group of waterbirds has its own specific hydrological and landscape needs; as a result, management efforts to conserve freshwater wetland habitats should be specific for different functional groups of waterbirds.
Among the hydrological variables, the average water depth of BBP should be maintained at a level of 1.8 m for supporting populations of small waders and vegetation gleaners. Levels of dissolved oxygen in the water should be maintained at a minimum of 5.5 mg/l to support food resources for fish-eaters. This operation can be jointly implemented by restoring the total area of the waterbodies—which was 3,820 ha in 2014—to a maximum of 6,500 ha, as above this size, the total area of waterbodies is shown to have negative effects on waterbird species. Providing this area of waterbodies will not only help to maintain water depth but will also maintain dissolved oxygen levels due to the greater surface area of the water. Regulating the water usage of rice farmers around BBP during the dry season can help to maintain the waterbody area in BBP (Sriwongsitanon et al., 2009). Maintaining an appropriate spatial area of waterbodies could also help to maintain a suitable area of marshes. Management should seek to maintain marsh areas at 6,500 ha, 6,700 ha, and 10,000 ha for ducks, large waders, and vegetation gleaners, respectively. The invasive weed Mimosa pigra should be eliminated by the regular pulling of young sprouts and the cutting of mature plants before flooding. The resprouting ability of mimosa significantly decreases underwater, leading to its death (Son et al., 2004). To reduce the resource competition between waterbirds and humans, there is a dire need to explore livelihood strategies and the factors causing an increase in human population inside the conservation zone. Furthermore, livelihood practices, which are in conflict with the waterbird conservation, need to be replaced with those practices that can benefit both people and wildlife. For example, enhancement of the habitat value of paddy fields for waterbirds in California's Central Valley has resulted in great benefits for the farmers and improved habitat to support the waterbirds (California Rice Commission, 2014). Similarly, restricting the use of pesticides and herbicides, decreasing the application of chemical fertilizers, using efficient irrigation processes, and providing economic incentives to the rice farmers who could maintain shallow waters in abandoned rice fields could provide alternative habitat for some waterbird groups. Most importantly, a proper protection status along with law enforcement is urgently required if BBP is to continue to sustain its diverse resident and migratory avifauna.
This study represents the most comprehensive assessment of waterbirds and their environmental associations yet conducted for any wetland in Southeast Asia. In general, this study will not only fill the information gap on waterbird populations in BBP and Thailand but will also contribute to waterbird conservation in Southeast Asia and along the East Asian-Australasian Flyway because waterbird populations are significantly declining in this region and quality information is lacking (Mundkur et al., 2017). Meanwhile, the quantitative approach used in this study would be of use in supporting authorities to evaluate waterbird-environment associations and to undertake strategies for effective conservation.
Footnotes
Appendix A. Annual Mean Abundance of Resident Waterbird Species Detected in the Bung Boraphet Wetland,Thailand.
Appendix B. Line charts showing trends in hydrological and landscape variables tested over 12-year study period (2003–2014).
Acknowledgments
The authors the officers and researchers of Bung Boraphet Wildlife Research Station, Department of National Parks Wildlife and Plant Conservation for assistance in fieldwork and waterbird surveys during 2014 and associate professor Philip D. Round for providing expert advice, valuable insights, and comments on this manuscript multiple times. They also thank the three anonymous reviewers for their constructive comments.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The first author is grateful to the Higher Education Commission of Pakistan and the Asian Institute of Technology, Thailand for providing funding support for this study through the PhD scholarship granted to him.