
Abstract
The Military Macaw (Ara militaris) is a globally threatened species with a fragmented distribution, and assessing the genetics of populations could help identify conservation units. Nine microsatellites were used to analyze 86 samples in seven localities along the Sierra Madre Occidental, the Sierra Madre del Sur, and the Sierra Madre Oriental in Mexico. Results showed that the Military Macaw has moderate levels of genetic diversity, similar to that found in other macaw species in Latin America. This species shows a high genetic structure; we find a genetic break between localities separated by the Central Plateau and the Trans-Mexican Volcanic Belt, which serve as geographic barriers. However, the locations within each genetic group are not genetically differentiated. It was observed that three locations of the Military Macaw have excess homozygotes, which could indicate a small effective size of the population and in combination with genetic isolation could increase the risk of extinction of the species. We propose two genetic groups for the species, the first comprising localities in the Sierra Madre Occidental and the Sierra Madre del Sur, and the second comprising localities of the Sierra Madre Oriental. According to the genetic differentiation, which was significant between the physiographic regions, and the unique allelic richness shown in this study, these two groups should be considered as independent conservation units. We strongly recommend the conservation and restoration of the natural habitats of the Military Macaw to maintain and increase the size of its population and recover and expand its original geographical distribution.
Introduction
The analysis of genetic diversity and its distribution within and among populations of endangered species may be used to inform conservation programs (Astorga & Ortiz, 2006). Knowledge of the intraspecific distribution of genetic diversity may identify genetically distinctive groups that can be proposed as genetic conservation units to maintain genetic diversity (alleles and genotypes) of a threatened species or population (Féral, 2002; Moritz, 1994a). Conservation genetics aims to investigate these patterns and the evolutionary processes of natural populations, with particular emphasis on endangered species (Moritz, 1994a); it can also identify potential and real threats that endanger the survival of such taxa (Moritz, 1994a, 1994b), so that appropriate action can be taken for their protection (Faria, Guedes, Yamashita, Martuscelli, & Miyaki, 2008; Moritz, 1994b). However, the sampling of endangered species is often a challenge, since these species usually have small populations, exhibit high habitat specialization that restricts their distribution, and are located mostly in habitats that are difficult to access (Bates, 2000).
In the case of macaws, studies of levels of genetic diversity have been mainly in South America (Caparroz, Miyaki, & Baker, 2003; Faria et al., 2008; Myers & Vaughan, 2004; Nader, Werner, & Wink, 1999; Presti et al., 2013; Presti, Oliveira-Marques, Caparroz, Biondo, & Miyaki, 2011). These studies have found some correlation between genetic diversity and the level of threat; Spix’s Macaw (Cyanopsitta spixii, extinct in the wild), the Lear’s Macaw (Anodorhynchus leari, endangered), and the Hyacinth Macaw (A. hyacinthinus, vulnerable) are macaw species with low genetic diversity (expected heterozygosity between 0.37 to 0.47), mainly due to hunting, illegal trade, and populations drastically reduced and isolated by habitat loss (Faria et al., 2008; Presti et al., 2013). In contrast, species such as the South American Scarlet Macaw (Ara macao) and Red-and-Green Macaw (A. chloropterus) present high genetic diversity (expected heterozygosity from 0.78 to 0.87), as do Scarlet Macaw populations in Central America (Nader et al., 1999), because their populations are larger and more stable (Caparroz et al., 2003; Myers & Vaughan, 2004).
In Mexico, there have been few studies that estimate the genetic diversity in populations of Psittacids (García-Feria, 2009). Studies of the genetic diversity of other bird species in Mexico have underscored the importance of geographical barriers (extensive mountain ranges) in determining distribution patterns and the genetic population structure of resident birds (Mock, Theimer, Rhodes, Greenberg, & Keim, 2002; Proudfoot, Honeycutt, & Slack, 2006). Two clear examples are the Ferruginous Pygmy Owl (Glaucidium brasilianum) and the Wild Turkey (Meleagris gallopavo), both found in the United States and Mexico. Within Mexico, populations of the Ferruginous Pygmy Owl of Sonora and Sinaloa were found to be genetically distinct from those of other populations (Proudfoot et al., 2006). Wild Turkey populations in eastern and western Mexico occur in distinct clades, showing a high genetic differentiation (Mock et al., 2002). For both species, a combination of geographic barriers and shifting vegetation regimes caused by environmental changes may have restricted gene flow between populations (Mock et al., 2002; Proudfoot et al., 2006). Data from mitochondrial sequences for the Military Macaw (Ara militaris) show a phylogenetic structure with geographical concordance for Mexico, supporting a disjunct distribution corresponding to the two extensive mountain ranges (Sierra Madre Oriental and Occidental) (Eberhard, IñIgo-Elias, Enkerlin-Hoeflich, & Cun, 2015).
The Military Macaw has a disjunct geographic distribution; it is found in Mexico as well as in South America (Venezuela, Colombia, Ecuador, Peru, Bolivia, and Argentina). Within each of these seven countries, the species has an irregular distribution coinciding roughly with that of tropical dry forests (Collar, 1997). This emblematic bird, it is considered vulnerable due to habitat destruction and illegal trade (Birdlife International, 2013). The species is listed as vulnerable by the International Union for Conservation of Nature (CITES, 1998) and is listed in Appendix I of the Convention on International Trade in Endangered Species of Fauna and Flora (CITES, 1998); in Mexico, it is listed as endangered (NOM-059-SEMARNAT-2010) (SEMARNAT, 2010).
Both the global population size and geographic range of the Military Macaw are in decline (Birdlife International, 2013; Collar, 1997). In Mexico, the species is found in the mountains of the Sierra Madre Oriental (Gulf of Mexico slope) (Arizmendi & Márquez, 2000; Carreón, 1997; Howell & Webb, 1995; Iñigo-Elías, 1999, 2000a, 2000b; Marín-Togo et al., 2012; Peterson & Chalif, 1989; Rivera-Ortiz, Contreras-González, Soberanes-González, Valiente-Banuet, & Arizmendi, 2008; Rivera-Ortiz et al., 2013), and the Sierra Madre Occidental and the Sierra Madre del Sur (Pacific slope) (Gaucín, 2000; Howell, & Webb, 1995; Peterson & Chalif, 1989; Rivera-Ortiz et al., 2013), with roosting populations of 20 to 78 macaws. This reduced population is due mainly to the loss and fragmentation of habitat. There is an annual rate of 3.27% to 5.21% deforestation of Military Macaw habitat in Mexico (Iñigo-Elías, 1999). In a period of almost 30 years, 29% to 32% of Military Macaw distribution area has been deforested and local extinctions have occurred along the Pacific slope (Marín-Togo et al., 2012; Ríos-Muñoz & Navarro-Sigüenza, 2009; Rivera-Ortiz et al., 2008; Rivera-Ortiz et al., 2013).
Given these population characteristics, we would expect the Military Macaw to present high genetic diversity (heterozygosity), more like species of the genus Ara of South America than the highly threatened endemic macaw species of Brazil. We would also expect populations in the Sierra Madre Occidental and Sierra Madre Oriental to be genetically distinct and that populations might be separated genetically by physiographic regions, in light of the findings of Eberhard et al. (2015) for the Military Macaw and of studies of other species (Mock et al., 2002; Proudfoot et al., 2006).
In the present study, we evaluated the population distribution in Mexico of the genetic diversity of the Military Macaw to identify conservation units and offer a proposal for long-term conservation of the species. We conducted a broad sampling across its current geographic distribution to assess the levels of genetic diversity and genetic structure that characterize remnant populations (Iñigo-Elías, 1999, 2000a; Ríos-Muñoz & Navarro-Sigüenza, 2009; Rivera-Ortiz et al., 2013).
Methods
Area of Study and Sample Collection for DNA Analyses
The study area included three physiographic regions: the Sierra Madre Occidental, Oriental, and Sur; and macaw feathers were collected at site in each of the following states: Sonora, Sinaloa, Nayarit, Jalisco, Oaxaca, Tamaulipas, and Queretaro. All sample sites were in tropical deciduous or semi-deciduous forest, with an elevation above sea level of 800 a 2200 m, present a wide range of total annual rainfall ranging from 60 mm to 1800 mm, and an annual temperature of 22℃ to 26℃ (Appendix A).
Feathers were collected to serve as sources of DNA for genetic analyses. Primary and secondary feathers accumulating on the ground below nest were sampled. Each nest was occupied by a specific pair of birds, and we genotyped only one feather per nest, assuming that it represented one of the pair. We collected a total of 54 feathers from different nests in seven geographically isolated populations (Gaucín, 2000; Gómez-Garduño, 2004; Jiménez-Arcos, Cruz-Padilla, Escalona-López, Arizmendi, & Vázquez, 2012; Rivera-Ortíz et al., 2013; Rubio, Beltrán, Aviléz, Salomón, & Ibarra, 2007) (Appendix A). In addition, 32 feathers were sampled at a distance from the nests, at sites visited by foraging macaws. All of the genotypes identified from 86 feathers were statically contrasted to discard possible identical genotypes. The feathers sampled were cleaned in the field with 90% alcohol, individually stored in paper bags, and maintained at ambient temperature.
DNA Extraction and Genotyping
Locus Name and Sequences of the Eight Microsatellite Loci Used for Genetic Analyses of 86 Samples of the Military Macaw.
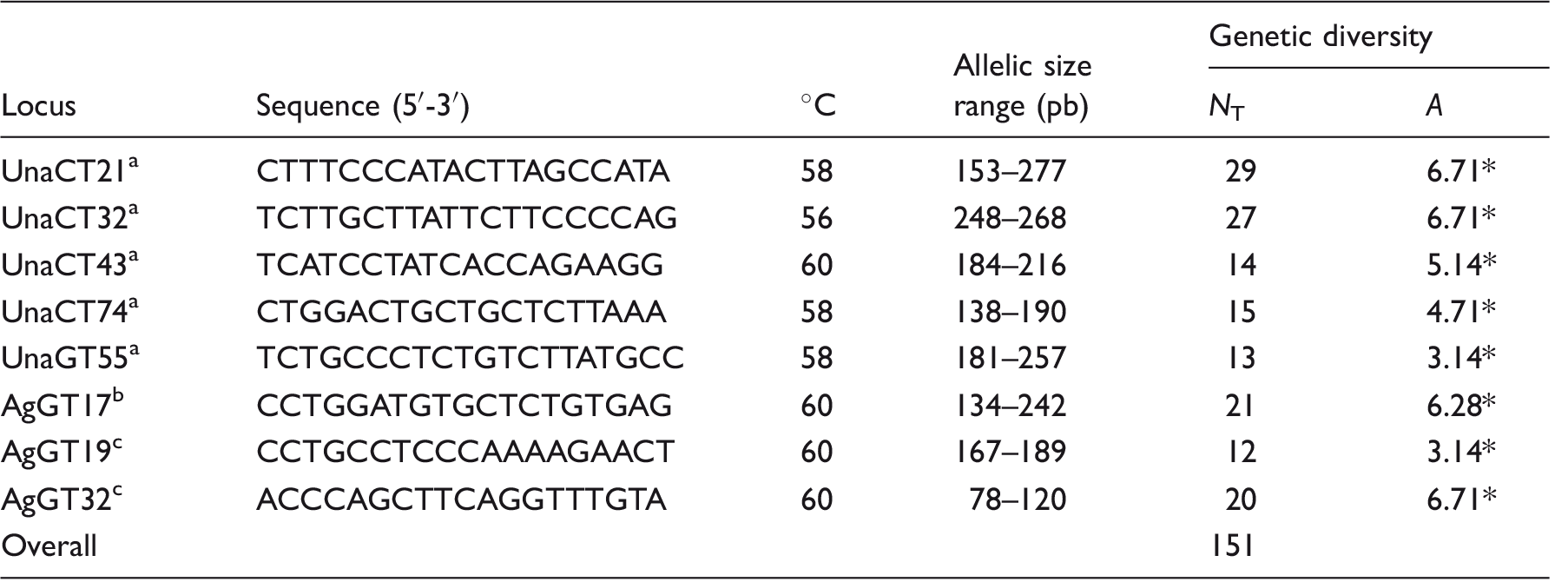
The annealing temperature and the allelic size variation correspond to specific results for this bird species. NT represents the total number of alleles identified by each locus among the sampling localities studied. A indicates the average number of alleles per locus among sampling localities.
HWD, Bonferroni correction, p < .0001.
The electrophoretic analyses were carried out in the ABI PRISM 3100-Avant sequencer (Applied Biosystems) using Gene Scan LIZ 500 to determine fragment size. The analyses of the fragments and their final size were determined using Gene Mapper 4.0 software (Applied Biosystems). Since this program automatically determines the size of the alleles, we visually checked the microsatellite electropherograms for all nine loci to corroborate their size and number. A few samples showed unclear electropherograms, and we repeated the PCR reactions to resolve these ambiguities (Morin, Chambers, Boesch, & Vigilant, 2001). The nine loci were tested for null alleles using Micro-Checker software (Oosterhout, Hutchinson, Wills, & Shipley, 2004). The UnaCT41 was the only locus discarded in the statistical analyses due to the presence of null alleles. Psex was calculated with GENELAX 6.3 software (Peakall & Smouse, 2006) and was used to detect possible individuals with identical multilocus genotypes, in order to identify multiple samples that might correspond to the same individual. No samples were found with identical genotypes, so all were used in the statistical analyses.
The statistical analyses both of genetic diversity (heterozygosity levels and allelic variation) and structure were organized hierarchically at the species level (considering all loci of all individuals from all sampling localities as a single population), at the group level by sampling localities corresponding to each of the three physiographic regions (Sierra Madre Occidental, Sierra Madre del Sur, and Sierra Madre Oriental), and by sampling locality, using ARLEQUIN 3.0 software (Excoffier, Laval, & Schneider, 2005).
Genetic Diversity
Genetic diversity was described by estimating allelic variation and heterozygosity levels. Since allelic richness (AR) and private allelic richness (PAR) estimation are affected by sample size, we employed the rarefaction method provided by ADZE 1.0 software (Szpiech, Jakobsson, & Rosenberg, 2008). Total number of alleles (NT), average number of alleles (A) by locus, total species heterozygosity (HT) (three regions together), and observed (HO) and expected (HE) heterozygosity for each sampling locality were estimated using the ARLEQUIN 3.0 software; heterozygosity values ranged from 0 to 1 (1 = highest heterozygosity) (Excoffier et al., 2005). Deviations from Hardy–Weinberg equilibrium (HWE) for loci and sampling localities were evaluated with the Markov chain method using the same software with 1000 dememorization steps, 50 batches, and 1000 iterations.
Inbreeding coefficients were calculated for each population (FIS) (Hedrick, 2005), using the Bayesian approach in INEst v. 2.0 (Chybicki & Burczyk, 2009); the estimation was conducted with 500,000 Markov Chain Monte Carlo (MCMC) cycles with every 200th updated and a burn-in of 50,000. Using the deviance information criterion, we compared the full model with a random mating model (where FIS = 0) in order to assess the statistical significance of inbreeding; if the resulting inbreeding coefficient is >0, there is an excess of homozygotes, and if it is ≤0, there is an excess of heterozygotes (Chybicki & Burczyk, 2009). We performed a Bonferroni correction test (<0.0001) as described in Sokal and Rohlf (2003) to test the significance of multiple comparisons for HWE.
The program BOTTLENECK 1.2.02 (Cornuet & Luikart, 1997) was used to infer a population bottleneck based on the significant differences between observed and expected heterozygosity of two mutation models: IAM (infinite alleles model) and SMM (stepwise mutation model). A total of 100,000 simulations were run in each sampling locality. If the p < .05 associated with the sign test the hypothesis of mutation-drift equilibrium is refused, consequently a population bottleneck might have occurred.
Differentiation and Genetic Distinctiveness
We estimated genetic differentiation (FST) among sampling localities within the three physiographic regions and also among physiographic regions (Weir & Cockerham, 1984). The estimation of FST was based on the IAM and the SMM with 10,000 permutations. The genetic distance matrix was also calculated, based on the proportion of shared allele (Bowcock et al., 1994; Milot, Gibbs, & Hobson, 2000), with ARLEQUIN 3.0 software. FST = 0 indicates no genetic differentiation in the populations studied and FST = 1 indicates genetic differentiation (Excoffier et al., 2005).
The genetic structure of the sampling localities studied was analyzed using STRUCTURE 2.3.1 software (Evanno, Regnaut, & Goudet, 2005). To choose the appropriate number of inferred clusters (K), we performed ten runs for each K from 2 to 10. All runs used a burn-in period of 500,000 steps, followed by 106 iterations under the admixture model, without any prior information. Using STRUCTURE HARVESTER software (Dent & Bridgett, 2012), we determined the most probable value of K using the maximum value of ΔK, following Evanno et al. (2005). We then estimated the cluster membership coefficient (Q) with CLUMPP 1.1.2 software (Jakobsson & Rosenberg, 2007), using the LargeKGredy algorithm with 1000 random input orders. These values were visualized using bar plots prepared with DISTRUCT software (Rosenberg, 2004).
A hierarchical test of population structure was estimated with analysis of molecular variance in ARLEQUIN 3.0 (Excoffier et al., 2005). The variance distribution between genetic groups, between localities in different groups, and within localities was compared (using the genotype groups obtained with STRUCTURE software); statistical significance was based on FST and tested with 10,000 permutations.
To determine the geographic location of the major genetic discontinuities (breaks) between populations, we used the maximum difference algorithm of Monmonier with BARRIER 2.2 software (Manni, Guérerd, & Heyer, 2004), with a matrix of average square distance (Goldstein, Linares, Cavalli-Sforza, & Fielman, 1995; Slatkin, 1995) for the seven sampling localities of Military Macaw. Resampling random subsets of individuals within populations provided 100 bootstrap replicate distances that were constructed utilizing MSA software (Dieringer & Schlötterer, 2003) to achieve statistical significance for the predicted barriers.
Results
Genetic Diversity
Genetic Diversity Estimates for the Military Macaw by Sampling Localities and by Physiographic Regions.

N = Sample size (individuals); NT = total number of alleles; AR = allelic richness; PAR = private allelic richness; HE = expected heterozygosity; HO = observed heterozygosity; HS = expected heterozygosity for each physiographic region; HT = total expected heterozygosity; HW = Tests for Hardy–Weinberg equilibrium; FIS = inbreeding index.
aSignificance level p = .05.
HWE, Bonferroni correction, p < .0001.
The AR was high for each physiographic region (Sierra Madre Oriental = 18.7, Sierra Madre del Sur = 18.0, Sierra Madre Occidental = 16.8). The average number of private alleles (PAR) was 5.7 for Sierra Madre Oriental, 4.9 for Sierra Madre del Sur, and 7.1 for Sierra Madre Occidental. The AR was high for all sampling localities, with values of 16.1 to 19.3; the PAR ranged from 4.9 to 8.5 (Table 2).
The total expected heterozygosity (HT) was 0.62, and similar values were found for each physiographic region (HS) (Sierra Madre Oriental = 0.54, Sierra Madre del Sur = 0.60, and Sierra Madre Occidental = 0.69). The observed heterozygosity (HO) levels of sampling locality varied from 0.51 to 0.69 and the expected heterozygosity (HE) from 0.54 to 0.76 (Table 2). Values of the inbreeding coefficient (FIS) in sampling localities showed positive values from 0.03 to 0.15. Sinaloa and Nayarit in the Sierra Madre Occidental and Querétaro in the Sierra Madre Oriental presented significant deviations from the HWE (pBC < 0.0001), indicating a significant excess of homozygotes (Table 2).
The results of testing for a recent bottleneck in Jalisco, Oaxaca, Querétaro, and Tamaulipas were nonsignificant (p > .050) for the two models analyzed (IAM and SMM). The IAM suggests a bottleneck in Sonora (p = .025), particularly in the locus AgGT32. Sinaloa and Nayarit showed evidence of bottlenecks only with the SMM (p = .0001), particularly in the loci UnaCT21, UnaCT32, UnaCT41, and AgGT32.
Genetic Differentiation
Paired Genetic Differentiation (FST) in the Seven Sampling Localities of the Military Macaw.

Total number of samples analyzed for each locality is given in parentheses. FST = *p < .05; ns = nonsignificant at p > .05; SO = Sonora; SI = Sinaloa; NAY = Nayarit; JA = Jalisco; OA = Oaxaca; TA = Tamaulipas; QUE = Querétaro.
Genetic Distinctiveness
In the STRUCTURE analysis, the ΔK statistic revealed K = 2 to be the optimum value for the number of genetic groups in the data. The proportion of ancestry of each population and individuals in these two genetic groups is shown in green and red (Figure 1). The localities of the Sierra Madre del Sur are not differentiated from those of the Sierra Madre Occidental representing the same genetic group to the other genetic group is shown only in the populations of the Sierra Madre Oriental, where the proportion of individuals with high membership values is between 0.8 and 0.9 for each group (Figure 1). The analysis of genetic differentiation between populations confirmed that the ancestry ratio of the admixtures is geographically structured (Table 3). Hierarchical analysis of molecular variance indicated that most of the genetic variation occurred within localities (ΦST = 0.054, p = .001), followed by variation among groups (ΦCT = 0.035, p = .001), while the differentiation among localities within groups accounted for the remaining variation (ΦSC = 0.114, p = .001).
Results of STRUCTURE analysis with K = 2. Each individual is represented by a vertical line, with colored segments in proportion to their membership in a genetic group. Black lines separate the different populations: Sierra Madre Occidental/Sierra Madre del Sur (1 = Sonora, 2 = Sinaloa, 3 = Nayarit, 4 = Jalisco, and 5 = Oaxaca) and Sierra Madre Oriental (6 = Querétaro and 7 = Tamaulipas).
In the BARRIER analysis, the maximum difference algorithm of Monmonier identified six genetic breaks, two with strong bootstrap support (95–100%) that separate the localities of the Sierra Madre Oriental from those of the Sierra Madre Occidental and the Sierra Madre del Sur (Figure 2). The four other genetic breaks have a minimum support of 10–20% (Figure 2).
Distribution of Military Macaw sampling localities in Mexico (shaded regions) and barriers determined with BARRIER analysis. 1 = Sonora, 2 = Sinaloa, 3 = Nayarit, 4 = Jalisco, 5 = Oaxaca, 6 = Querétaro, and 7 = Tamaulipas. Roman numerals indicate divisions between sampling localities, with thin lines indicating weak divisions (20% support) and heavy lines indicating strong divisions (95% support).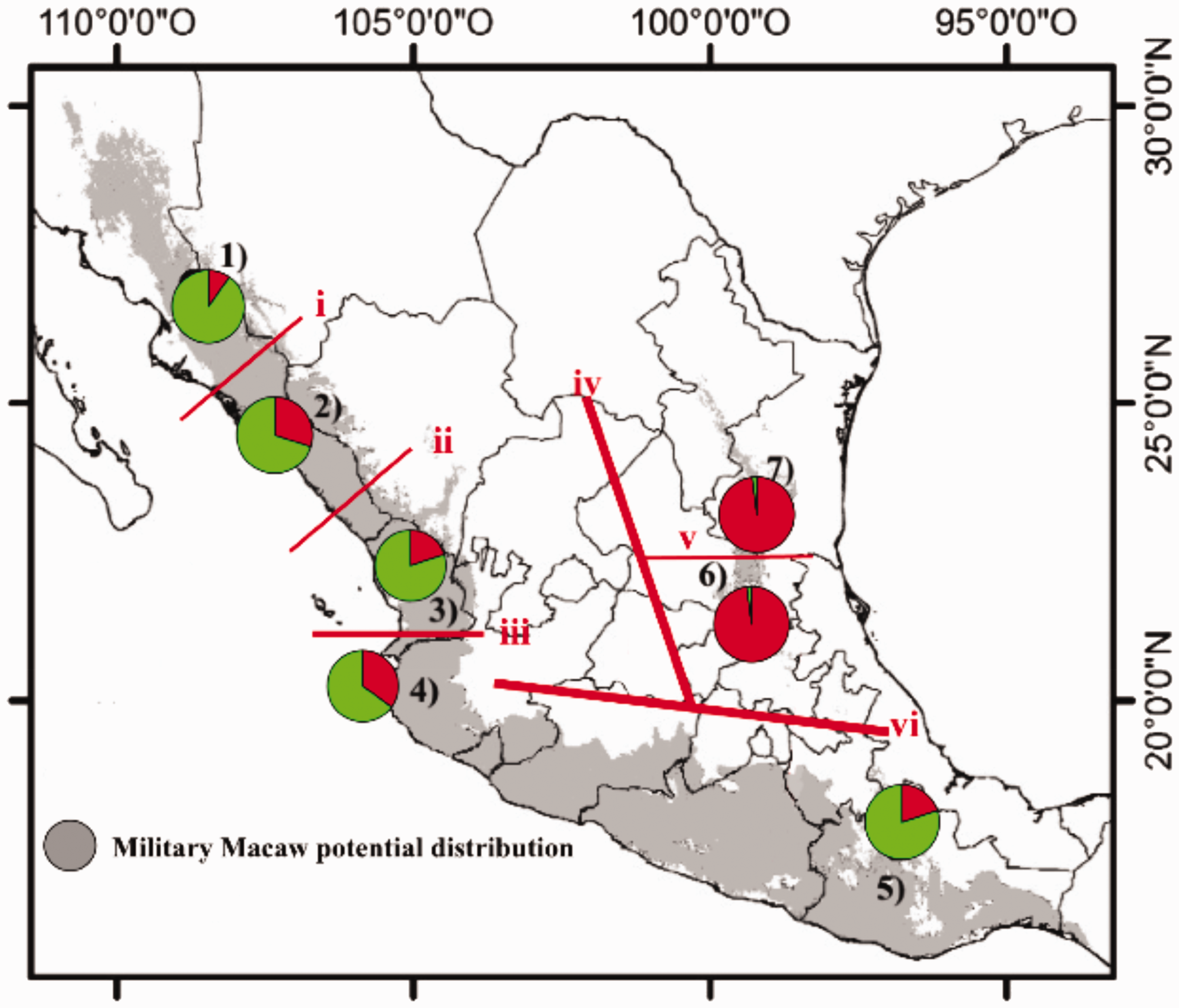
Discussion
The localities sampled in this study show moderate levels of allelic richness and heterozygosity for the Military Macaw that are similar to those of other Ara macaw species, although the diversity of the Military Macaw in Mexico was lower than that found for other Ara macaws in South America. As expected, this sample shows greater heterozygosity than that of the threatened endemic macaws of Brazil (Caparroz et al., 2003; Caparroz, Miyaki, Bampi, & Wajntal, 2001; Faria et al., 2008; Mock et al., 2002; Nader et al., 1999; Presti et al., 2011; Presti et al., 2013). These results may reflect a diversity in ancestral large populations that no longer occur (e.g., Nei, 1978), and the estimators may be affected by historical factors. It is also possible that population decreases are very recent, as has been seen in other macaw species (e.g., Caparroz et al., 2001). The Military Macaw has been severely overhunted in Mexico for the last 70 years to Cantú, Sánchez, Grosselet, and Silva (2007) estimate that illegal traffickers catch between 65,000 and 78,500 Mexican Psittacids annually, including more than 500 Military Macaws, with a mortality rate of 77% (Cantú et al., 2007). Another explanation for the genetic diversity found could lie in the birds’ movement over long distances. Average displacements of 20–30 km have been reported, with extensive movements in non-breeding season (Juárez et al., 2012).
Despite these indications of genetic diversity, we also find evidence of a bottleneck in three populations (Sonora, Sinaloa, and Nayarit), but these results should be taken with caution because we do not satisfy the program assumptions of 20 loci and 10 individuals per population. Our ability to detect significant bottlenecks based on levels of heterozygosity is limited by the small sample size (Bouzat, 2010; Williamson-Natesan, 2005).
We observed significant inbreeding coefficient values for two localities in the Sierra Madre Occidental (Sinaloa and Nayarit) and one locality in the Sierra Madre Oriental (Queretaro), these inbreeding values could portend problems for conservation. The relatively high homozygosity levels may indicate small effective sizes. In Querétaro, the fragmentation of habitat and the loss of connectivity between populations are severe problems, which can reduce populations (Gaucín, 2000; Reed & Frankham, 2003). For the populations of the Sierra Madre Occidental (Nayarit and Sinaloa), apart from the loss of habitat, hunting for illegal trade may increase extinction risks in the future (Carreón, 1997; Gómez-Garduño, 2004; Rubio et al., 2007). It has been observed in the coasts of Michoacán (Marín-Togo, 2012), where the species has been extirpated of the zone mainly for illegal trade. In general, the reduction in the effective population size of the Military Macaw is due to poaching (more than 100 individuals per year) (Birdlife International, 2013; Iñigo-Elías, 1999, 2000b) and loss of habitat (31% of its habitat has been reduced by the change in land use) (Marín-Togo et al., 2012; Ríos-Muñoz & Navarro-Sigüenza, 2009; Rivera-Ortíz et al., 2013) as has been reported in other macaws (Caparroz et al., 2001).
We found a clear genetic differentiation between sampling localities on the Gulf of Mexico slope (Sierra Madre Oriental) and those on the Pacific slope (Sierra Madre Occidental) and in central Mexico (Sierra Madre del Sur), which is consistent with a previous study by Eberhard et al. (2015) using mitochondrial sequences. That study found two clades in Mexico, one in the eastern populations and the other in the northeast. Our data suggest that the Military Macaw sampling localities along the Sierra Madre Oriental are closely related, as are those along the Sierra Madre Occidental and the Sierra Madre del Sur. These results indicate that this species is able to fly long distances (≈30 km); however, the effective dispersal seems to be constrained by the selective use of habitat and resources (Contreras-González, Rivera-Ortiz, Soberanes-González, Valiente-Banuet, & Arizmendi, 2009; Evans, Ashley, & Marsden, 2005; Myers & Vaughan, 2004). It would be necessary to have telemetry and banding data to fully understand the mobility of this species. Such data are not yet available for the Military Macaw mainly due to the difficulty of capturing and marking adult macaws (Nader et al., 1999). It would also be useful to analyze additional molecular markers, such as mitochondrial DNA (Evans et al., 2005).
Geographic isolation is apparently responsible for evolutionary isolation in the Military Macaw (Eberhard et al., 2015). Similar phenomena have been seen in other Mexican bird species, such as the Ferruginous Pygmy Owl (Proudfoot et al., 2006) and the Wild Turkey (Mock et al., 2002), where the genetic differences are due to the presence of geographic barriers such as mountain ranges (Trans-Mexican Volcanic Belt, Sierra Madre Occidental, and Sierra Madre Oriental) and the Central Mexican Plateau (Mock et al., 2002; Pritchard, Stephens, & Donnelly, 2000). We thus propose protection of the two genetic groups found in the three physiographic regions that have acted as evolutionary reservoirs (Sierra Madre Occidental/Sierra Madre del Sur and Sierra Madre Oriental), and that they be considered independent priority conservation units, namely management units (Moritz, 1994a, 1994b).
Implications for Conservation
Military Macaw populations have experienced both habitat fragmentation and high levels of illegal trafficking. These threats are a concern from a conservation perspective because they could potentially reduce gene flow and genetic diversity and thus produce increased inbreeding (Fahrig, 2003). Our results suggest that these threats have a strong impact mainly on the populations of Sinaloa, Nayarit, and Querétaro, where we saw high inbreeding values. These populations suffer from a high level of anthropogenic threats, mainly loss of habitat (Williamson-Natesan, 2005), so it is important to focus available resources on reproductive and foraging habitat restoration in order to reestablish demographic stability and maintain the genetic health of these populations.
Our study of the genetic structure of Military Macaw in Mexico has implications for conservation, since most of the localities we studied represent breeding areas, and therefore, they need effective protection actions at the regional level to preserve the habitat. It is necessary to decrease or eliminate anthropogenic threats in breeding areas in order to reinforce the demographic stability and maintain the genetic integrity of the remaining populations and preserve the overall genetic variability of the species. The sampled localities in the Sierra Madre Oriental require particular attention, because these habitats have been heavily affected by changes in land use (agriculture and ranching) and present only moderate genetic diversity, increasing the risk of a decline in populations and local extinction in the region (Rivera-Ortíz et al., 2013). As a first step, populations that are not being permanently monitored (Palmito, Sinaloa; Mirador del Aguila, Nayarit; Tuito, Jalisco; Papalutla, Guerrero; and Jaumave, Tamaulipas) urgently require protection, since they are possible sources for the illegal trade. It is also necessary to work with local communities to protect the natural habitat (mainly in the Sierra Madre Oriental) and reduce poaching.
A second strategy is to avoid the genetic isolation among the populations within each large physiographic area. This can be accomplished by restoring lost habitat and perhaps by establishing natural corridors that maintain connectivity. In addition, the specific effects of illegal trafficking should be studied, to determine the populations most affected and propose solutions. A well-supported program of ecotourism could be one of the best options for management and conservation, as in Santa Maria Tecomavaca, Oaxaca and the Bahia de Banderas region in Jalisco, where scientific research combined with community work and environmental education have been effective strategies to combat illegal trafficking and habitat loss.
Footnotes
Acknowledgments
We thank the various authorities for the facilities provided for the completion of fieldwork. A. L. Albarrán-Lara assisted with statistical analyses. V. Rocha and D. L. Aquino provided technical assistance in the laboratory. We also thank the anonymous reviewers for their suggestions for improving this manuscript. Many colleagues participated in field research and data collection, and the we are particularly grateful for the dedication shown by V. Garcia, A. M. Contreras-González, E. Berrones, C. Villar, H. Verdugo, and Y. Rubio.
Author Contributions
Francisco A. Rivera-Ortiz collected, analyzed the data and molecular markers, and also wrote the article; María del C. Arizmendi and Patricia Dávila-Aranda reviewed and corrected the article; Ken Oyama and Sofía Solórzano performed and analyzed the data, reviewed, and also wrote the article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author (s) disclosed the receipt of the following financial support for research, authorship and / or publication of this article: The authors wanted to thank the financial support was also provided by CONACyT Projects 60270 (S. Solórzano) and DT006 (M. C. Arizmendi). Logistic support was provided by the SDEI-PTID-02-UNAM (P. Dávila) project. F. A. Rivera-Ortiz is grateful for the support of a postdoctoral fellowship from the Dirección General de Asuntos del Personal Académico (DGAPA) of the Facultad de Estudios Superiores Iztacala (FES-I), the Graduate Program in Biological Sciences at the Universidad Nacional Autónoma de México (UNAM), and the Consejo Nacional de Ciencia y Tecnología (CONACyT).