
Abstract
Some tropical countries, such as Colombia, are venturing into the international carbon market by means of avoided deforestation programs, i.e., they seek carbon credits by not cutting down forests. Consequently, basic information about carbon storage in different forest types is urgently needed. We describe the tree community dynamics of forest fragments in cattle ranches of the western Orinoco basin, and their relationship to the forests’ ability to accumulate carbon. We re-sampled three 1-ha vegetation plots after six and seven years of establishment. We found that these forests have different population dynamics; smaller fragments have negative net population change, but the dynamics allow for an annual carbon accumulation rate of around 0.57 Mg ha−1. This rate is similar to the rate reported for some mature Amazon forests and may be attributed to high soil fertility.
Introduction
Carbon storage estimation in different ecosystems has become a central subject in conservation science, because natural ecosystems can mitigate climate change effects by acting as carbon sinks, limiting the availability of this greenhouse gas in the atmosphere (Ashton, Tyrrell, Spalding, & Gentry, 2012; Pan, Birdsey, Phillips, & Jackson, 2013). In tropical forests, which account for 55% of the carbon stored in the world’s forests, 65% of the carbon is stored as biomass in the roots, trunks, and leaves of woody plants (Pan et al., 2011). In Colombia, a tropical country, avoided deforestation initiatives have emerged during the last decade from a variety of stakeholders such as the agriculture and livestock industries, which hope to mitigate the damage of their own productive activities while offering the service of carbon storage to other productive sectors (Fedegan, 2006; Fedepalma, 2013). However, the impact of these initiatives is still unknown, because the carbon stocks in Colombia have been estimated only for primary forests (Alvarez et al., 2012; Phillips et al., 2016; Sierra et al., 2007).
High deforestation rates in Colombia have left vast areas of fragmented and secondary forests (Phillips et al., 2016). These forests, if conserved and restored, could be useful for carbon trade (Saatchi et al., 2011). Research on land-use changes and ecosystem services is still scarce (Etter, McAlpine, Wilson, Phinn, & Possingham, 2006; Rodríguez Eraso, Armenteras-Pascual, & Retana Alumbreros, 2012; Sánchez-Cuervo, Aide, Clark, & Etter, 2012) and is needed for planning conservation strategies in geographic areas dedicated to agriculture and livestock.
Estimations of carbon accumulated in tropical forests depend on climate, soil type (since in cold climates and anoxic conditions carbon usually accumulates in soils), and the structure of plant communities and their productivity (Carvalhais et al., 2014; Lähteenoja, Ruokolainen, Schulman, & Oinonen, 2009). The amount of carbon in plants is divided in two components, above and below ground, which are estimated based on allometric equations (Gibbs, Brown, Niles, & Foley, 2007). However, these equations are more common for aboveground biomass (AGB), using information on the diameter of trees at breast height (DBH) and wood density (Alvarez et al., 2012; Chave et al., 2005). In some cases, equations can also include tree height (Feldpausch et al., 2012; Goodman et al., 2013). The ecosystem service related to carbon storage from the atmosphere may also depend on forest dynamics, because a forest in which the recruitment of new trees is higher than mortality would show high accumulation rates (if variables such as wood density and tree height are kept constant) (Johnson et al., 2016).
Cattle ranching is a productive activity involving deforestation, the use of forest resources, fragmentation, and edge effects associated with high mortality of large trees (Laurance, Delamónica, Laurance, Vasconcelos, & Lovejoy, 2000), which are the main drivers of AGB stocks (Slik et al., 2013). Fragments, especially small or narrow forest fragments, have a strong relationship between perimeter and area, and thus are susceptible to large edge effects (Nascimento & Laurance, 2004). Forest fragments show drastic changes in plant species composition, because there is high recruitment of pioneer species near edges (Laurance et al., 2006). As pioneer trees are characterized by low wood density and usually short stature (Muller-Landau, 2004), it is expected that forest fragments should have greater abundance of low wood density species. In addition, the mortality of large trees caused by more wind exposure and changes in abiotic conditions should exacerbate the pattern (Laurance et al., 2011).
Our main objective is to describe the tree community dynamics of forest fragments in cattle ranches of the Orinoco Basin, located in the Andean piedmont, and to determine their relationship to carbon accumulation. Specifically, we want to: 1. Determine growth, mortality, and recruitment rates of woody plant species in these forest fragments; 2. Quantify the variation in wood density of tree species in these fragments; and 3. Estimate the carbon stocks and carbon accumulation rates of the forest fragments. Considering the predictable changes in the tree community, we expected to find a low estimate of carbon storage in the forest fragments and a negative balance of carbon accumulation over time.
Methods
Field study
The municipality of San Martín (Meta, Colombia) is one of the oldest cities in the region, founded in 1585. The surrounding region has since been used for extense cattle ranching in large areas. The original landscape was mainly continuous forest from the foothills of the Andes, with some natural savannas to the southeast. Land-use change estimations from the Colonia (Etter, 2015; Etter, Sarmiento, & Romero, 2010) show that fragmentation has been occurring since the late 1600s, resulting in a landscape of riparian forest fragments (Appendix 1).
The remaining forest in the area has been categorized as tropical moist forest (Romero Ruíz, García, García, i Pascual, & Ruíz, 2004). The mean temperature is 27.5°C, ranging from 19 to 35°C (Stevenson, 2011). Annual rainfall in the region is usually between 2,600 and 3,000 mm and is seasonal, with two dry months with less than 100 mm (usually January and February) (Stevenson, Castellanos, Cortés, & Link, 2008). During the last three decades cattle ranches have been increasingly replaced with oil palm plantations, making the Meta Department the most extensive producer in Colombia (Wagner, Castro, & Stevenson, 2007). It has been observed that when grasslands are free from human intervention, a secondary forest is established (Stevenson, 2011).
We re-sampled three forest plots (1 ha) in forest fragments in three cattle ranches in San Martín (Appendix 1). We established the plots during the first semester of 2005 (Stevenson & Aldana, 2008) and the second semester of 2009. Fragment 1 has an approximate area of 28 ha, and the 1-ha plot (3°36′30.40″N, 73°38′20.72″W) was re-sampled six years after establishment (second semester of 2011); Fragment 2 has an approximate area of 23 ha, and the 1-ha plot (3°37′42.02″N, 73°38′19.86″W) was re-sampled seven years after establishment (second semester of 2012); Fragment 3 is the biggest remaining fragment in the municipality, 1,000 ha in area; the 1-ha plot (3°34′54.9″N, 73°27′14.7″W) was re-sampled six years after establishment (second semester of 2015).
During the re-sampling of the plots we measured the increased DBH of trees greater than 10 cm, recording the number of dead individuals and new recruits. We measured height of 76% of the individuals in our database, using a laser clinometer. We collected wood cores to determine wood density, here measured as Wood Specific Gravity (WSG sensu, Williamson & Wiemann, 2010), of at least one individual from the most abundant species, following the protocol proposed by Chave (2005). Green volume was calculated using the dimensional method, and wood cores were oven dried at 103°C for 72 hours.
Data analysis
We compared the census data from 2005 (Stevenson & Aldana, 2008) and 2009 with census data from 2011 – 2015 to determine annual growth, mortality, and recruitment rates for each plot using the formulae presented by Sherman and collaborators (Sherman, Fahey, Martin, & Battles, 2012).
We calculated above ground biomass (AGB) for each sampling year, using the allometric equations for Tropical Moist Forest of the type I, which take into account DBH, height and wood density. We used the best predictive model for neotropical moist forests (Type I.3) (Alvarez et al., 2012). Biomass calculations for other woody life forms were done using different allometric equations to minimize errors; for palms we calculated the biomass using the equations proposed by Goodman et al. (2013), and for lianas we calculated the biomass using the equation proposed by Schnitzer, DeWalt, and Chave (2006).
For cases where the WSG was not measured by us (33% of species), we used information on species measured by our research group, following the same protocols (Casas et al., 2016), and values reported in the Dryad global database (Chave et al., 2009; Zanne et al., 2009). For species with no reported WSG values in either of the databases, we used the mean for the genus or family (using a combined database with the three sources mentioned previously), assuming that this trait is phylogenetically conserved (Chave et al., 2006). To evaluate whether mean WSG was different among plots we performed an ANOVA and a Tukey test, using the information from the year of re-sampling.
The net change in AGB was calculated as the difference between the AGB in 2011 – 2015 and the AGB in 2005–2009, divided by the number of years between each sampling. We compared the results among plots and between sampling years with a t-test.
Results
Population Dynamics
Community dynamics of forest fragments: Recruitment, mortality, net population change and average relative growth rates of the tree communities of three forest plots (1ha) in fragments located in cattle ranches of the Andean piedmont in Meta, Colombia.

Wood Specific Gravity
The distribution and mean values of Wood Specific Gravity for woody plant species differ among forest fragments (Figure 1). The analysis showed differences among fragments (ANOVA, F = 0.26, P < 0.01) and a Tukey test showed that mean WSG is lower in fragment 1.
Histogram of the distribution of wood specific gravity values for tree species in three forest fragments of the Andean piedmont in Meta, Colombia. Values were assigned to each individual stem from a species average calculated based on the measurements made by us in the present study and complemented with information from unpublished data (Casas et al., 2016). For species missing in our database (10%) we used information from Global Data Bases (Zanne et al., 2009). Dotted lines represent the mean values for each fragment.
Above Ground Biomass Change
The estimations of the rate of biomass change were positive for fragments 1 and 3 and negative for fragment 2 (Figure 2). These forests fragments, on average, are accumulating 0.57 Mg of AGB per hectare per year. Sierra et al. (2007) calculated that carbon biomass content in moist tropical forests is around 45%, from which we estimate that, on average, each hectare of these forest fragments is accumulating 0.26 Mg of carbon every year.
Above Ground Biomass change in three forest plots (1-ha) in fragments located in cattle ranches of the Andean piedmont in Meta, Colombia, after 6 and 7 years of establishment.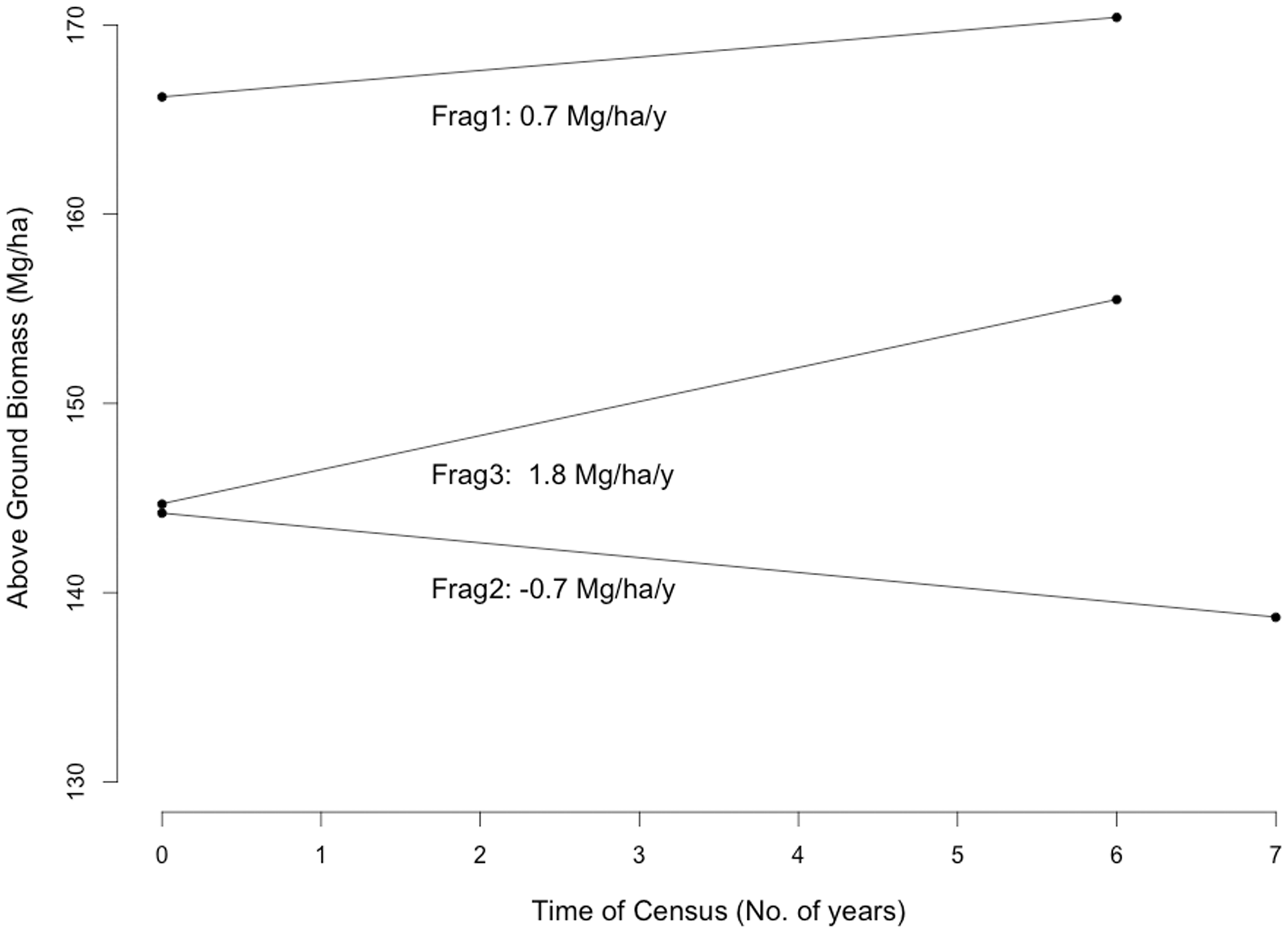
Growth rate and Wood Specific Gravity
Given the surprising result of positive AGB change and negative stem density change in fragments 1, we explored the possibility of the mortality rate being compensated by the rapid growth of some tree species. A negative relationship between growth rates and wood density is expected in contexts of r and K strategies. Although we did not find this relationship, we found high WSG values (WSG > 0.6) for some species with high relative growth rate (RGR) (RGR > 2%). These results are summarized in Figure 3, which was drawn with species that had more than two individuals with observations of RGR in our database.
Relationship between Relative Growth Rate and Wood Specific Gravity for 63 tree species (species in our data set with more than 2 observations), found in forest fragments located in cattle ranches of the Andean piedmont in Meta, Colombia. Species colored in red may be of interest for native wood production and ecosystem restoration (WSG>0.6, RGR>2%). Species names have been shortened for easier reading, species names from top to bottom are: Eschweilera cabrerana (cabo de hacha), Hieronyma oblonga (candelo), Hirtella americana (moradito), Garcinia madruno (madroño), Eugenia sp., Alchornea discolor, Licania subarachnophylla.
Discussion
Population Dynamics
We found that the smaller fragments have a negative change in stem densities, due to a mortality rate higher than the recruitment rate. A higher mortality rate can be attributed to selective logging that has been occurring in these fragments in past years. Similarly, a lower recruitment rate may be due to intensive use by the cattle that go daily into the forest to drink water from the streams within these fragments. Although we do not have measurements of the effect of cattle in these forests, cattle presence in the forest may cause high seedling mortality and soil compaction (Hernández-Vargas, Sánchez-Velásquez, Carmona-Valdovinos, & Cuevas-Guzmán, 2000).
This idea is supported by a positive change in stem densities for fragment 3, which is isolated from cattle presence. Also, the negative effect of selective logging may be attenuated by the size of the fragment.
Wood Specific Gravity
We found that fragment 1 has a lower WSG mean value, which is possibly an effect of selective logging and low species richness (67 species per hectare). Fragment 2 is not richer in species (69), but some species are not shared, which allows for a higher frequency of species with higher WSG values (Figure 1). Fragment 3 is the most diverse in species (127) and it is also the fragment with the greatest frequency of species with high WSG values. This result is consistent with the reduction in fragment size. However, in our study system selective logging may have eliminated individuals of some old-growth species from the fragments, specially fragment 1 (Stevenson & Aldana, 2008).
Biomass Stocks and Above Ground Biomass Change
Comparing our results with previous estimations, based on extrapolations (Phillips et al., 2011), we found that these forests have lower biomass stocks than previously estimated.
We found that fragment 2 had a negative change in AGB which maybe a consequence of a negative change in stem density; opposite to this pattern, fragment 1 had a negative stem density change, but a positive rate of AGB accumulation, which may be due to a high RGR (see below). Although fragment 3 is more conserved than the other two fragments, it has not reached a neutral carbon balance as have primary forests of the Amazon (Phillips et al., 1998) and it accumulated more AGB per year than the other fragments. In the tropics, the Amazon region has been subject of many studies of AGB accumulation, which show that Amazon primary forests tend to be on an equilibrium state in terms of carbon storage (Hernández-Vargas et al., 2000). Therefore, this result can be explained by past removal of large trees for domestic wood use, which allows for growth and recruitment of competing trees. Fragments 1 and 2 also have a higher RGR than fragment 3, which means that some tree species in these fragments may have high growth rates and high WSG values (see below).
Because these three forest plots may not be representative of all the forest fragments in the Orinoco Basin, the results of this study should be compared with other studies from the same region, in order to reach sound conclusions about the carbon dynamics of forests fragments of the region.
Growth rate and Wood Specific Gravity: Promising Tree Species
The tree species we highlight in Figure 3: Eschweilera cabrerana (cabo de hacha), Hieronyma oblonga (candelo), Hirtella americana (moradito), Garcinia madruno (madroño), Eugenia sp., Alchornea discolor, Licania subarachnophylla could be considered promising for restoration and reforestation programs in the Andean piedmont, as these are native species that grow fast and could enhance the ecosystem services of the forest by accumulating more carbon in their trunks (given their wood density).
Implications for conservation
Although AGB stocks in these forest fragments are lower than in Amazonian primary forests (Baraloto et al., 2013), we found that these forests are capable of accumulating at least 0.26 Mg of carbon every year, very similar to many primary forests in the Neotropics (Phillips, Lewis, Baker, Chao, & Higuchi, 2008). We believe this accumulation rate is due to fertile soils and could be enhanced by reducing selective logging and cattle use of the forest. If well managed and protected, these fragments could be used by cattle ranchers, oil palm producers, and other productive sectors of the region either to compensate for the impacts of their activities or to receive payment for ecosystem services that enhance forest cover and connectivity between forest fragments. Connectivity among these forest fragments would not only benefit tree species populations, but also populations of big mammals like Panthera onca and other native fauna. We found some native tree species that have high values of WSG and are faster growing than other native species; these species should be studied for their potential for timber production, and if successful, may be used to replace Eucalyptus pellita (eucalipto – Red Mahogany) and Pinus caribea (pino - Caribbean Pine). Plantations of the latter species are replacing natural forest and savanna ecosystems in the region.
Footnotes
Appendix 1
Detail of the study area in the municipality of San Martín (Meta, Colombia) in the western Orinoco Basin. Yellow squares show the location of the three 1-ha vegetation plots in the forest fragments. Areas shaded in orange are oil palm plantations, which may be confused with forest fragments. Light green areas are pastures for cattle ranching. Images were created using GoogleEarthPro in June 2016.
Acknowledgements
The Sánchez-Rey family and the Enciso family were very kind to let us work on their premises; they also helped with fieldwork logistics. Sasha Cárdenas, Marcela Córdoba, Indira León, Camila Monje, Ángela Perilla, Erika Rodríguez, Vanessa Rubio, Luis Francisco Henao, Eduardo Pinel, Diana Acosta, Edna Beltrán, Efraín Rincón, Camilo Quiroga, Maria Juliana Pardo, Felipe Aramburo and Ángela Sánchez helped with the re-sampling of the plots and wood specific gravity measurements. We thank an anonymous reviewer who helped us improve the quality of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors received finantial support from Fondo de Investigaciones of the Facultad de Ciencias at Universidad de Los Andes. AMA received support from the Lóréal – Unesco grant For Women in Science.