
Abstract
Objectives
Pteris vittata L. is one of the hyperaccumulator plants that can grow in heavy metal-contaminated soil. This plant may tolerate metal stress through different mechanisms, eg, by producing a wide range of secondary metabolites. These compounds play an essential role in physiological function and can also serve as rich sources of chemicals for health benefit purposes. This study aims to determine the phytochemicals of plant extracts and the relationship between these profiles with pharmacological properties to predict active agents and their interaction.
Methods
The total phenolic and total flavonoid contents were measured by colorimetric assays, while the widely targeted metabolomics was applied to reveal leaf and root P. vittata metabolome. In parallel, the antioxidant and antibacterial effects of these extracts were determined by ABTS scavenging and micro-broth dilution methods. Multivariate analysis, ie, Partial Least Square Regression, calculated the correlation between metabolites and bioactivities.
Results
The results showed the significant content of phenolics and flavonoids in plants collected from mining soils of Thai Nguyen province, Vietnam, and their potential ABTS scavenging activity. Additionally, the plant's metabolome characterized 82 compounds, mainly secondary metabolites. The multivariate analysis indicated the positive correlation among quercetin derivatives and bioactivity, suggesting the contribution of their combination, not individually, to the antibacterial effect of P. vittata.
Conclusion
Hyperaccumulators in metalliferous soils are potential reservoirs for exploring the antibacterial agents against multi-drug resistant strains, and this effect may be exerted by additive or synergistic mechanisms of action of multiple compounds.
Keywords
Introduction
Heavy metals are toxic to plants in many ways, depending on their different states. At the cellular and molecular levels, these agents can impact various plant physiological processes, for example, disrupting the normal function of macromolecules such as enzymes and proteins, making chelates with functional groups of metabolically critical small molecules, and modifying the conformational of membrane integrity. 1 As a consequence, plant metabolism could change, leading to variation in the metabolite profile of plant samples.2–5 In addition, heavy metals can disrupt the redox-oxidative balance of plant cells by inducing the formation of free radicals and reactive oxygen species (ROS). 6 However, several plants could survive in the metalloid areas and still grow and mature normally. There are two main ways to resist heavy metal stress: avoidance or tolerance. The former modality induces the restriction of metal uptake, and the latter activates many procedures that allow plants to adapt in the presence of heavy metal inside. Indeed, plants can produce a range of metabolites or binding ligands that are intracellularly chelated with heavy metals, upregulating the antioxidant defense against the negative impact of ROS. 7 Phenolics, known as the most important secondary metabolites, play an essential role in the oxidative defense system of the plant kingdom. 8 Increased phenylpropanoid metabolism and phenolics level can be observed in some species under heavy metal contamination. 9 This group, especially flavonoids, can be considered the chemical barrier for oxidative stress caused by elements.
Besides the effect on plants, heavy metals also put critical pressure on bacterial communities in metalliferous soils. 5 In long-term exposure to these agents, bacterial strains become more and more resistant to available biocides. This phenomenon was explained partly by horizontal gene transfer of heavy metal resistance genes and antibiotic resistance genes induced by high levels of metals in soils. 10 Also, efflux pumps are found to have a higher frequency in pollution ecosystems because of their effectiveness in the detoxification of exogenous elements.11–17 Indeed, this is an effective mechanism for microorganisms to extrude various heavy metals and compounds with diverse structures, not limited to antibiotics used in clinical activities, thus representing multidrug-resistant (MDR) phenotypes. A wide range of bacterial species was reported to express efflux systems, and exposure to multiple heavy metals induced their over-expression more effectively. 18 Stenotrophomonas maltophilla is one of these species. The strain is often associated in plant rhizosphere, and is commonly used in bioremediation because of its antifungal properties. 19 Furthermore, a wide range of isolates of S. maltophilla was detected in both environmental samples and clinical samples, suggesting that these MDR phenotypes spread and transmitted across different matrices in the ecosystems.20–24
Interestingly, plant metabolites may be responsible for the expression or the dispersion of MDR phenotypes, thus, possibly involved in bacterial growth. Indeed, i) plant antimicrobial compounds, eg, berberine, are sometimes substrates of MDR pumps, 19 and many pieces of evidences indicate the structural similarity between plant compounds and substrates of bacterial MDR pumps (eg, polyphenols and polycyclic aromatic hydrocarbons)25–27; ii) plant produce compounds able to induce the expression of MDR pumps in rhizosphere bacteria 28 ; and iii) plant produce compounds able to inhibit bacterial MDR pumps (Efflux Pump Inhibitors- EPIs), allowing to reduce the MIC of antimicrobial compounds that are the substrate of these pumps.29–31 For all the above-mentioned reasons, plants growing in metalliferous soil have been considered a potential source for screening the antioxidant and antibacterial effects because of the deep interaction with heavy metals and MDR phenotypes in the ecosystem.
This research focuses on the most dominant fern, Pteris vittata L., which was collected in lead and zinc mine at Hich village, Tan Long, Dong Hy, Thai Nguyen. Many studies have been conducted to reveal the metal tolerance mechanism of this hyperaccumulator. 32 The metabolome profiles of P. vittata have been characterized, providing a solid database for dereplicating metabolites within these extracts and indicating a reservoir of compounds for human benefit purposes. 33 Some pharmacological effects of P. vittata have been shown in previous investigations34–37; however, no studies on the antibacterial effects and the efflux pumps inhibition potential of P. vittata have been reported. For the first time, we explore the chemicals inside this hyperaccumulator, such as total phenolic content (TPC), total flavonoid content (TFC), the metabolite profile and their potential to inhibit ROS and MDR bacterial growth. In addition, multivariate analysis revealed the relationship between these samples’ chemical profiles and bioactivities, particularly against S. maltophilia, which suggested a new approach to identifying bioactive compounds against MDR phenotypes. The transversal interaction of plants, bacteria and heavy metals was significantly highlighted within metalloid ecosystems in our study.
Material and Methods
Chemicals
For TPC, gallic acid, Na2CO3 solution, Folin–Ciocalteu reagent were purchased from Merck (Darmstadt, Germany). To determine TFC, NaNO2, AlCl3, NaOH, and ethanol were obtained from Aladdin (Shanghai, China); quercetin was purchased from Vietnam National Institute of Drug Quality Control (Hanoi, Vietnam). In the ABTS assay, Trolox, methanol, and K2S2O8 originate from Merck (Darmstadt, Germany). Distilled and ultrapure water were prepared using a Millipore Milli-Q purification system (Millipore GmbH, Schwalbach, Germany).
Plant Materials
35 plant individuals of Pteris vittata L. were gathered at different places in Hich village, Tan Long ward, Dong Hy district, Thai Nguyen province, Vietnam (N: 21o43′612″ E: 105o51′380″) under dry weather conditions. They were named in order from A1 to A35 for aerial parts and S1 to S35 for belowground parts. Plant identification was conducted by Dr Nguyen The Cuong, Institute of Ecology and Biological Resources, Vietnam Academy of Science and Technology. The voucher specimens were stored in the Department of Life Science, University of Science and Technology of Hanoi.
Bacterial Materials
The tested bacteria were six different Stenotrophomonas maltophilia strains: 502, 0366, K279a, PierC1, BurA1, and BurE1. Among them, 502, 0366, and K279a are resistant bacteria isolated from clinical samples, while PierC1, BurA1, and BurE1 originated from environmental materials. These isolates were distinguished by their antibiotic resistance profiles, as mentioned in the article of Youenou et al, 2015. 20 The bacterial strain was grown in Tryptone soya broth (TSB) medium (HIMEDIA, India) and incubated at 37 °C with shaking for 16-18 h in the incubator. The optical density value of approximately 0.4 was measured by spectrophotometry (equivalent to 109 CFU/mL) in TSB. These suspensions were diluted in TSB to get stock solutions containing 2.106 CFU/mL as bacterial density.
Methods
Sample Preparation
For plant treatment, the whole plant was divided into aerial and belowground parts, washed to remove contamination, and then rinsed in distilled water. All tissues were cut down into small parts and dried in the oven at 55 °C for 48 h until their weight did not change. Dry tissues were then ground into powder. Subsequently, 100 mg of each sample were extracted with 80% methanol at room temperature in a sonic bath for 30 min. 38 The extracts were concentrated in the vacuum concentrator to obtain dry extracts. These extracts were subjected to all chemical tests in further experiments.
Total Phenolic Content
The TPC was measured using the Folin-Ciocalteu assay with gallic acid as a standard. 39 The extracts were weighed and dissolved in methanol at different concentrations. Subsequently, 40 μL of the diluted sample was mixed with 480 μL of Folin-Ciocalteu reagent and 480 μL 6% Na2CO3. The mixture was then incubated at 40 °C for 15 min and measured the optical absorption at the wavelength λ = 765 nm. For the standard curve, a series of gallic acid (C7H6O5) solutions from 100 to 1000 μg/mL was prepared, and the same reaction with the tested sample was followed. The TPC was measured as mg gallic acid equivalent per g extract (mg GAE/g).
Total Flavonoid Content
For the TFC measurement, a colorimetric assay followed the procedure of Nurcholis et al 40 The samples were similarly weighed and dissolved in methanol at different concentrations. A mixture including 240 μL of the initial solution and 40 μL 5% NaNO2 was incubated at 25 °C for 6 min. Then, 40 μL 10% AlCl3 was added to the mixture and put at the same temperature and duration as the previous step. Finally, 400 μL NaOH 1 M and 280 μL 30% ethanol were mixed with, incubating for 15 min at 25 °C. The solution was measured at the wavelength λ = 510 nm. A calibration curve was built by a series of standard solutions of quercetin (C15H10O7) from 0 to 200 μg/mL. The TFC was calculated as mg quercetin equivalent per g extract (mg QE/g).
Metabolite Profiling
In our previous investigation, UPLC-QqQ-MS was used for the rapid dereplication of a wide range of phytochemicals in leaf and root extracts of P. vittata individuals. 33 In this study, this approach was applied to 70 crude extracts. The same parameters for separation in liquid chromatography and detection in mass spectrometry were set up following our published articles. For data processing, metabolites were selected according to their relative standard deviation (RSD) compared to those in quality control samples and the signal-to-noise ratio (S/N). Selected peaks should ensure two criteria, including the RSD not greater than 30% and S/N not smaller than 3. The intensity of these peaks was normalized by internal standards before being integrated for multivariate analysis.
2,2-Azinobis 3-Ethylbenzothiazoline-6-Sulfonic Acid (ABTS) Scavenging Assay
ABTS assay is commonly used to determine the antioxidant capability of samples. 41 The crude extracts were dissolved in methanol at the stock concentration of 10 mg/mL and then diluted in 0.25 and 0.5 mg/mL as two tested concentrations. To prepare the standard curve, the stock of 5 mg/mL Trolox in methanol was used to make a series of 1/64, 1/96, 1/128, 1/192, and 1/256 as dilution factors. The absorption of these solutions is measured in 96-well plates at 734 nm by a microplate reader. The ABTS scavenging activity was calculated according to the formula: % scavenging capacity = (Ac-As)/Ac×100, where Ac and As are the absorbance of the standard and the tested sample, respectively.
Antibacterial Activity
The inhibition potentials of P. vittata L. crude extracts against different bacterial strains 502, 0366, K279A, PierC1, BurA1, and BurE1 were determined by micro-dilution assay with some modification. 42 Stock solutions (20.48 mg/mL) were prepared in dimethyl-sulfoxide (DMSO) from all the dry extracts; then, twentyfold dilution was made with TSB to reach a concentration of 1.024 mg/mL. Subsequently, 100 μL of each extract was first aliquoted into a 96-well plate. Equal volumes of bacterial suspension prepared in TSB mentioned above were introduced to the wells. The final concentration of extract in wells was 512 μg/mL, and that of DMSO was 2.5%. The bacterial growth was determined by the optical density of each well, measured at the wavelength of 600 nm. Each experiment was performed in triplicate, and the reported value was the average. Gentamicin at the concentration of 32 μg/mL was used as positive control. The well containing sterile TSB was blank, while DMSO at 2.5% in TSB was used as negative control.
Statistical and Multivariate Analysis
For statistical analysis, t-tests and one-way ANOVA were applied to determine whether the difference between the two variables is significant using Graphpad Prism 9.2.0. The univariate analysis, ie Pearson correlation and the multivariate analysis was done by using RStudio version 3.5.3. (http://www.R-project.org/). Peak intensity was normalized by taking their logarithm base 10. The correlation between the whole metabolome and bioactivities was calculated by the plsr function in the mixOmics package. 43 Figures were built using ggplot2 R-package. 44
Results and Discussion
The results were presented in three main parts. The metabolite profiles reported the TPC, TFC and metabolomes of 35 leaf and 35 root methanolic extracts. In parallel, the bioassay part demonstrates the antioxidant and antibacterial characteristics of these 70 extracts. Finally, univariate and multivariate analysis integrated chemical and biological profiles to reveal the relationship between metabolites and effects.
Total Phenolic Content and Total Flavonoid Content
Figure 1 shows the distribution of TFC and TPC in the methanolic extract of 35 aerials and 35 belowgrounds. It was indicated that TPC varied a lot in both tissues, ranging from 5.78 to 29.53 and from 7.37 to 33.17 mg GAE/g dry weight for those leaf and root extracts, respectively. All the samples showed remarkable TPC, suggesting that phenolics represented as major components in the extracts. The TFC of P. vittata ranged from 2.59 to 7.28 mg QE/g extract in leaves and from 2.38 to 7.67 mg QE/g extract in roots. These data indicated that flavonoids occupied a high proportion of the phenolics profile. The t-test and one-way ANOVA calculations show no significant statistical difference between aerial and belowground parts in TFC and TPC (p-value > 0.05).

TPC (mg GAE/g dry extract) and TFC (mg QE/g dry extract) of 70 samples: 35 aerial parts (from A1 to A35, in orange color) and 35 belowground parts (from S1 to S35, in blue color).
Metabolome Profiling
By the widely targeted metabolomics analysis, 70 extracts were profiled and a total of 82 metabolites were selected according to the criteria mentioned in the method section (Table 1). It is indicated that amino acids and carboxylic acids dominate the metabolome of both aerial and belowground parts of P. vittata, with 32 representatives belonging to this group. Other primary metabolites, sugars, occupied significantly the list. These primary metabolites are vital for many physiological functions of cells and organisms, including being precursors for synthesizing secondary metabolites. On the other hand, the secondary metabolites profile is more diverse in terms of structure that contribute to the adaptation of plant against environmental conditions. Indeed, flavonoids are main secondary metabolites detected in the profile of P. vittata. Twenty-five compounds belong to this group with flavonols (quercetin, kaempferol, isorhamnetin), flavons (apigenin, luteolin, vitexin, peltatoside), anthocyanidins (cyanidin), flavan-3-ols (catechin, procyanidin C1), flavanones (eriodictyol, hesperetin, naringenin, rhoifolin) as aglycone skeletons. The glycoside unit attached in the structure also varied greatly in these flavonoids. Additionally, several simple phenylpropanoic acids and coumarins, together with flavonoids, make the phenolics profile of these samples.
Metabolite Profiling of P. vittata Methanolic Extracts by Widely Targeted Metabolomics.

Each metabolite has its code (ID), annotation, precursor and product, retention time and the group that it belongs to.
Naturally, flavonoids have a significant role in plant defense and adaptation, ie, regulating the development, pigmentation, flavoring, and UV protection. 45 These functions are supposed to be emphasized in metalliferous soils. 46 As an overdose of metals can generate ROS; plants produce more phenolics and their subgroup, flavonoids, the most important components in antioxidative systems. Additionally, these compounds possess a wide range of health benefits such as anticancer, antimicrobial, antiviral, and neuroprotective activity thanks to their diverse structures.47–50
Quantitatively, the intensity of selected peaks could be valuable data. The table gathers the peaks’ intensity normalized by internal standards, which can be seen in the Supporting Information, Table S1. Indeed, the metabolome is the most time-sensitive parameter in system biology because metabolites’ levels can change rapidly during the life span of an organism. Even though plant individuals grow in the same region, their metabolome varies, leading to their differences in bioactivities.51,52 Therefore, we recorded the variation in the intensity of selected peaks among 35 leaf and 35 root extracts to correlate them to the bioactivities data.
The Capability to Scavenge the ABTS Radical
The scavenging capability of ABTS radical of 70 extracts of P. vittata at the concentration of 0.25 and 0.50 mg/mL was illustrated in Figure 2 and Supporting Information Table S2. There was a significant variation in the antioxidant capacity of tested samples. At the concentration of 0.25 mg/mL, the values ranged from 4.07 to 52.74% and from 10.59 to 79.42% for aerial and belowground parts, respectively. When testing at a bigger concentration, the figure for two parts increased, from 18.07 to 90.16% and from 23.08 to 90.25% in the given order. The t-test and one-way ANOVA revealed that the antioxidant activity of belowground parts is significantly greater than the antioxidant activity of aerial parts (p-value < 0.05).

The inhibition percentage (%) of ABTS scavenging activity of 70 samples: 35 aerial parts (from A1 to A35, in orange color) and 35 belowground parts (from S1 to S35, in blue color).
Antibacterial Activity
To identify the most sensitive isolates of S. maltophilia with P. vitatta extracts, we screened at the first stage the response of these samples on six strains, including BurA1, BurE1, PierC1, 0366, K279a and 502. Three leaf samples and three root samples were prepared by mixing equally powders from 35 aerial and 35 belowground parts to ensure their representativeness. The mixed materials were then extracted with MeOH 80%, as indicated in the method section. Overall, six combined extracts, including three aerial and three belowground parts, demonstrate a wide range of inhibition rates from 32% to 75% at the concentration of 512 μg/mL against all six S. maltophilia strains (Figure 3). The growth curve of bacteria under the treatment of extracts was shown in Supporting Information Figure S1. Among bacteria isolated from the environment, BurA1 was the most sensitive isolate, followed by BurE1 and PierC1, while the order for clinical strains is 0366, K279a and 502. These trends were simultaneously found in all mixtures. Regarding genotypic features, K279a, 0366, and 502 were the multi-drug resistant S. maltophilia strains isolated from a bloodstream infection, which was shown a wide array of antibiotic resistance genes, ie, efflux pumps. BurA1, BurE1, and PierC1 originated from soils and represented different susceptibility profiles against antibiotics. 20 The study of Youenou et al 2015 also showed that K279a, BurA1, and BurE1 demonstrated low antibiotic susceptibility, whereas PierC1 was sensitive. In addition, BurA1 and BurE1 showed increased resistances compared with strain K279a. Looking at the activity of extracts on these isolates, the susceptible/resistant characteristics were correlated with the inhibition percentage: plant extracts exhibited a better effect against the more sensitive bacteria. As a consequence, these extracts, especially root tissues, present the most remarkable antimicrobial activity against 0366 strains.

Inhibition percentage of six mixed extracts (pool 1,2,3 - aerial parts, pool 4,5,6 - belowground parts) on different strains of S. maltophila PierC1, BurE1, BurA1, 502, 0366 and K279a.
For further experiments, we examined the activity of 70 crude extracts on these isolates to find out the relationship between the whole metabolome profile and antimicrobial activity. Supporting Information Table S2 illustrates the inhibition percentage of 70 samples against 0366 isolates, as an input for multivariate analysis.
Correlation among Chemical Profile and Bioactivities of P. vittata Extracts
Univariate analysis does not show any significant correlation between TPC, TFC, and the scavenging activity ABTS and inhibition percentage against S. maltophilia 0366. This result suggested that the variation in the total phenolic content and total flavonoid content did not reflect the variation in bioactivities (Supporting Information Figure S2). Additionally, no single compound represented a significant correlation with antioxidant and antimicrobial activities (Supporting Information Table S3).
Multivariate analysis, ie, partial least square regression (PLSR), were then conducted to reveal correlations between the whole profile and antioxidant/antimicrobial effects. This model was generally illustrated by a circle plot (Figure 4). These metabolites are located inside a 2-components space spanned by a circle of radius 1 (correlation coefficient 1) and a radius 0.5 (correlation coefficient 0.5). The relationship between the two variables is measured by the angle created by the endpoint of the variable vectors. The positive correlation is found if the angle is sharp, negative in the case of an obtuse angle, and null (having no meaning) if the angle is right, 90°. Therefore, following the cross-arrows quadrant separation, metabolites that fall into quadrant II can be predicted as having a positive correlation with antibacterial activity (Y). In contrast, quadrant IV predicts a negative relationship with Y, while quadrants I and III present no correlation at all.
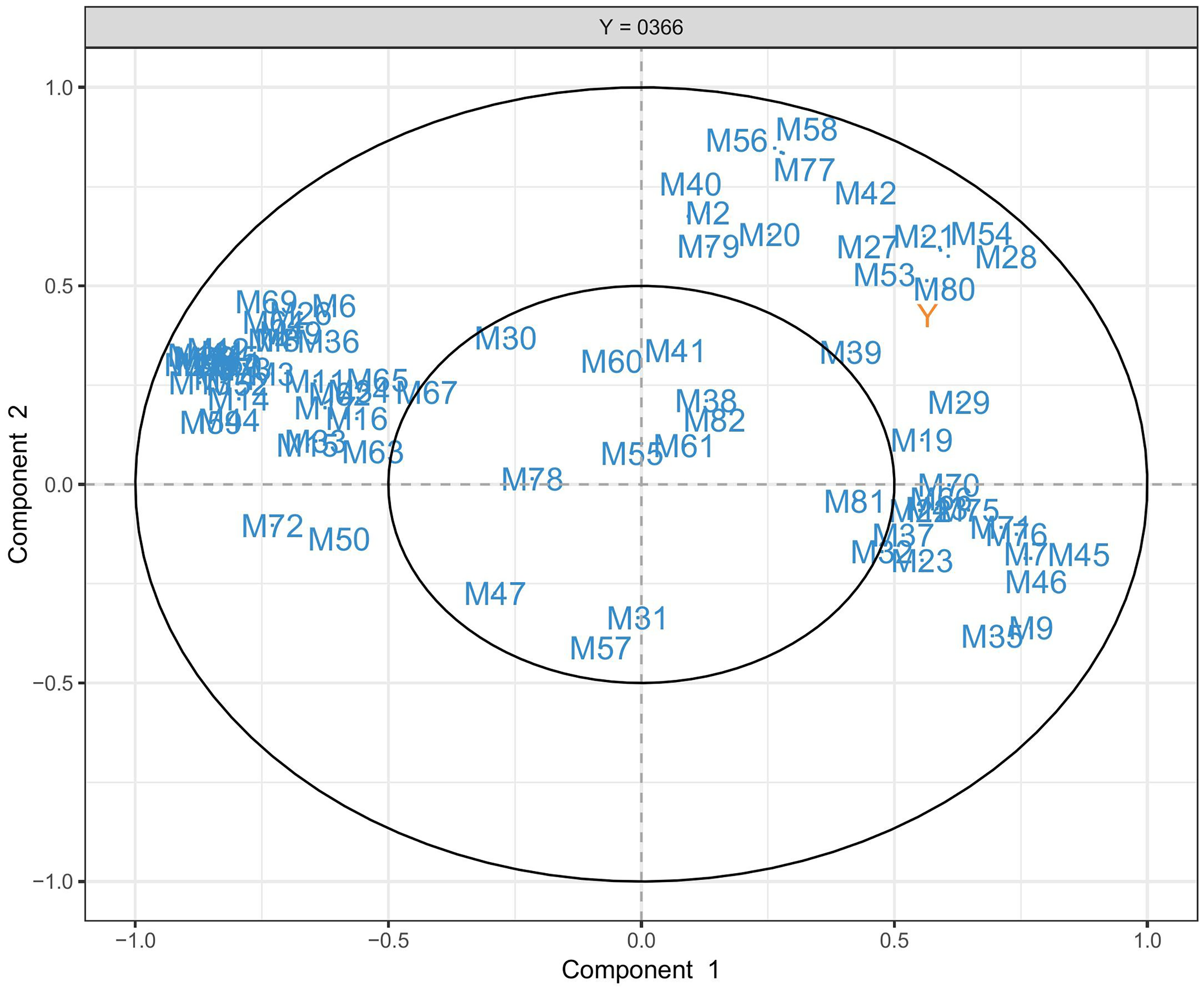
PLSR-based loading plot illustrates the correlation between the whole profile and the antimicrobial effect of leaf extracts.
Figure 4 illustrates the circle plot of the PLSR model between 82 metabolites and the inhibition percentage of 35 leaf extracts of P. vittata. The quadrants I and III were dominated mainly by primary metabolites, indicating that these agents did not figure out any interaction with antibacterial activity. Other compounds located in quadrant IV, ie, M72 (betaine) and M50 (L-carnitine), negatively correlated to the vector Y, thus may support the growth of bacteria. Remarkably, seven flavonoids, mostly quercetin aglycone with different sugar units, including M21 (hyperoside), M27 (hesperidin), M28 (neohesperidin), M42 (quercetin-3-arabinoside or quercetin-3-D-xyloside), M53 (quercitrin or quercetin-7-O-rhamnoside), M54 (rutin or quercetin-3-O-b-glucopyranosyl-7-O-a-rhamnopyranoside) and M80 (homoorietin) position into a cluster of the quadrant II. They group into three smaller clusters, a group of M19 (rhoifolin), M29 (vitexin), M39 (isorhamnetin-dihexose), a group of M56 (luteolin-4'-O-glucoside) M58 (maritimein or luteolin-7-O-glucoside) and M77 (kaempferol-3-O-glucoside), and another group of M2 (kaempferol-3-O-alpha-L-arabinoside), M20 (peltatoside), M40 (kaempferol dihexose), M79 (apigenin-7-O-glucoside). Generally, this distribution on the correlation circle plot suggested that these compounds belong to the same biosynthetic pathway. The similarity in the structures of compounds in the same subgroup proves this hypothesis.
In addition, the cluster of flavonoids is located near vector Y, highlighting that these compounds correlated significantly to bioactivity and are thus responsible for the effect. Note that no single correlation was found through univariate analysis, while the multivariate can reveal strong coefficients, reaching around 0.9 (nearly the cycle of radius 1) among flavonoids and bioactivity. Therefore, these compounds may work additionally or synergistically to contribute to the inhibition percentage of plant extracts against S. maltophilia 0366. In natural product-based drug discovery, the “single compound – single target” has shifted to “multiple compounds – multiple targets” recently. As natural products are produced by complicated biosynthetic pathways, these complex pools can interact with different proteins. They can exhibit different mechanisms of action on various targets inside an organism, ie, bacterial cells, to give bioactivity. 53 Previous literature has shown that mass spectrometry profiling with assays can reveal the additives and synergists of plant compounds to enhance the antimicrobial effect of berberine. 54 Our study has demonstrated that the mixture improves the inhibition percentage of individual compounds by either antibacterial or resistant modifying characteristics. To investigate the mode of action of the combination, further experiments should be conducted to unravel the interaction of molecules and different biological targets.
In this study, the prediction power for the inhibition percentage of the metabolome profile has provided a good example of how to identify bioactive compounds in combination. At the same time, conventional strategies, such as bio-guided isolation, cannot detect this mechanism. However, our study still has limitations. The extracts obtained are crude extracts that provide moderate effects on a single bacterial strain. While our findings provide initial insights on the anti-S. maltophilia effect, testing on other species would offer a more comprehensive assessment of the antibacterial power of selected flavonoids. Otherwise, further experiments can be performed on fractions and pure compounds isolated from these extracts, solely or in the mixture, to confirm the model's accuracy. From a more applied point of view, the pipeline developed by this research can be considered metabolomics-guided isolation as the chromatogram of extracts proposes an indication for the isolation of pure substances through their retention time value and separation conditions. This approach is more efficient, more economical, and more targeted than conventional natural products drug discovery like bio-guided isolation.
Conclusion
In a nutshell, this study demonstrates for the first time the potential of P. vittata as a hyperaccumulator with significant secondary metabolites, particularly phenolics and flavonoids, that exhibit both ATBS scavenging and antibacterial properties against several types of multi-drug-resistant S. maltophilia. The positive correlation between quercetin derivatives and the inhibition percentage of 35 leaf methanolic extracts suggests that the antibacterial effect may arise from an additive or synergistic interaction of multiple compounds, not a single action. This finding not only highlighted the ecological function and health benefit of P. vittata but also underscored its potential as a source for compounds against multi-drug resistant bacterial strains. For perspectives, the in vitro experiment of isolated compounds should be performed to confirm the hypothesis. Otherwise, further experiments may be set up to reveal the mechanism of action of additive/synergistic combination at the molecular level.
Supplemental Material
sj-docx-1-npx-10.1177_1934578X251322868 - Supplemental material for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds
Supplemental material, sj-docx-1-npx-10.1177_1934578X251322868 for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds by Phuong Nhung Nguyen, Ngoc Lien Nguyen, Hai Nam Pham, Phuong Khanh Nguyen and Kieu Oanh Nguyen Thi in Natural Product Communications
Supplemental Material
sj-xlsx-2-npx-10.1177_1934578X251322868 - Supplemental material for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds
Supplemental material, sj-xlsx-2-npx-10.1177_1934578X251322868 for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds by Phuong Nhung Nguyen, Ngoc Lien Nguyen, Hai Nam Pham, Phuong Khanh Nguyen and Kieu Oanh Nguyen Thi in Natural Product Communications
Supplemental Material
sj-xlsx-3-npx-10.1177_1934578X251322868 - Supplemental material for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds
Supplemental material, sj-xlsx-3-npx-10.1177_1934578X251322868 for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds by Phuong Nhung Nguyen, Ngoc Lien Nguyen, Hai Nam Pham, Phuong Khanh Nguyen and Kieu Oanh Nguyen Thi in Natural Product Communications
Supplemental Material
sj-xlsx-4-npx-10.1177_1934578X251322868 - Supplemental material for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds
Supplemental material, sj-xlsx-4-npx-10.1177_1934578X251322868 for Antioxidant and Antibacterial Activities of Pteris vittata L. Extracts from Metalliferous Soils: Correlations with Phenolic Compounds by Phuong Nhung Nguyen, Ngoc Lien Nguyen, Hai Nam Pham, Phuong Khanh Nguyen and Kieu Oanh Nguyen Thi in Natural Product Communications
Footnotes
Acknowledgements
The authors would like to thank the UMR 5557 Ecologie Microbienne (Eq Bacterial Efflux & Environmental Resistance), Lyon, France and the International Joint Lab on “Drug Resistance in Southeast Asia” (LMI-DRISA) to donate & store the bacterial strains for this research.
Author Contributions
NLN and PNN are co-first authors of this article. TKON and PNN conceived of the presented idea. TKON conducted the widely targeted analysis. NLN, HNP, and PKN carried out the experiments related to TPC, TFC, ABTS and antibacterial activities. PNN and TKON conducted the multivariate analysis. All authors discussed the results and contributed to the final manuscript.
Data Availability
All data was included in the article or supporting information.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the University of Science and Technology of Hanoi [grant number USTH.LS.01/22-24].
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.