
Abstract
The chemical composition of the essential oils of Passiflora sexocellata and Passiflora trifasciata (Passifloraceae, subgenus Decaloba) were studied for the first time. Essential oils were obtained by steam distillation of fresh leaves and flowers. The chemical composition was assessed by using GC/FID and GC/MS. For P. sexocellata leaves, the optimized analytical procedure allowed the identification of 33 compounds (75% of the total oil composition) and 29 (74% of the total oil composition) in flowers. Regarding P. trifasciata, 35 compounds (76% of the total oil composition) were detected in leaves and 32 (71% of the total oil composition) in flowers. Terpenes and mono unsaturated hydrocarbons were quantified as major constituents of the volatile fraction in flowers (17.0 to 52.6%) and (13.7 to 20.0%). Organic acids were detected in both leaves and flowers with a percentage ranging from 3.3% to 32.0%. Aldehydes were also detected in leaves (12.6 to 41.4%) and in flowers (1.4 to 5.1%). The GC/MS analyzes allowed alcohols to be detected in leaves (20.6 to 42.9%) and in flowers (8.2 to 18.1%). These compounds represent the most important feature of the large Passiflora family. Moreover, a critical role in the coevolved mechanisms of pollinators' interaction has been investigated.
Passiflora trifasciata Lem., and Passiflora sexocellata Schltdl. are two tropical vines of the subgenus Decaloba (DC.) Rchb., family Passifloracee, with tendrils to aid in climbing and they differ from the most common subgenus Passiflora L. because of relatively small flowers, two to 3 series of coronal filaments and a chromosome number of 2n = 12. 1 Passiflora trifasciata (Figure 1(A)) is included in the supersection Decaloba 2 and it is an endemic species in Ecuador, Perù and Brasil. The tri-color passionvine produces single or coupled white fragrant flowers with green pistils and stamens that standout against the beautifully marked foliage. This species is cultivated as ornamental. Passiflora sexocellata (Figure 1(B)) is included in the supersection Cieca (Medic.) J. M. a monophyletic group primarily distributed in the southern United States, México, Central and South America, and the Caribbean. P. sexocellata is found from southern Mexico to Nicaragua. It possesses peltate bilobed leaves bearing 4‐13 laminar nectaries often with 2‐4 of the nectaries occurring proximal to the lateral leaf veins. Flowers are often produced in long inflorescences and the fruits are spherical and possess more than 40 arillate seeds. Passiflora. sexocellata is pollinated by small to medium guild bees (Colletes sp.) as reported for the majority of the species in the supersection Cieca (e.g., P. coriacea, P. mcvaughiana, P. megacoriacea, P. trinifolia). The flower coronal filaments act as a landing platform for pollinators and have conspicuous areas of yellow and/or purplish coloration to attract them. Moreover, the more advanced species of supersection Cieca (P. xiikzodz, P. juliana, P. viridiflora, P. coriacea, P. sexocellata, P. megacoriacea) are mainly visited by tropical butterflies. In particular Dryas julia and Heliconius erato are P. sexocellata herbivory in México and Central America. 3 The close association of species in the Heliconiinae and Passifloraceae is commonly reported as an example of plant-insect coevolution. Scent plays an important role in foraging, sometimes acting synergistically with color. The importance of visual and olfactory signals most likely varies with species, with each species having a unique favored combination of color cues and chemical compounds. 4 GC/FID and GC-MS coupled with steam distillation has been employed to analyze fresh flowers and leaves of P. mucronata (subgenus Passiflora) in order to assess the role of floral volatiles in plant pollination. 5 Long chain methyl and ethyl esters, accounting for 50.8% of the essential oil from flowers, seems to be key of attraction for bats such as Carollia perspicillata and Glossophaga soricina, driven by the olfactory stimuli of these compounds when visiting P. mucronata for foraging. A high content of odd linear long-chain hydrocarbons was detected in fresh tissues of three bee-pollinated Passiflora hybrids, acting as allelochemicals in plant herbivore interactions. 6 In this work we have studied, for the first time, the chemical composition of essential oil obtained by steam distillation of both fresh leaves and flowers of P. sexocellata and P. trifasciata in order to assess their role in plant pollination and plant association with “passion flower butterfly”.

Passiflora trifasciata Lem. (
Results and Discussion
The yields of P. sexocellata and P. trifasciata essential oil obtained by steam distillation from fresh leaves and flowers were quantified and reported as a percentage of 0.04% and 0.06%, 0.07% and 0.11% (weight/fresh weight basis), respectively. Table 1 shows the results of qualitative and quantitative oil analyzes on the Elite-5MS column. The compounds are listed in order of their elution and are reported as percentages of the total essential oil, determined using octyl octanoate as internal standard. For P. sexocellata leaves, the total number of peaks was 44 with a number of 33 identified peaks (75% identification), for the flowers was 39 with a number of 29 identified peaks (74% identification). For P. trifasciata leaves, the total number of peaks was 46 with a number of 35 identified peaks (76% identification) and, for flowers was 45 with a number 32 identified peaks (71% identification). Differences in the qualitative and quantitative composition of the obtained essential oils have been observed.
Composition of Essential Oils From Leaves and Flowers of P. Sexocellata and P. Trifasciata.
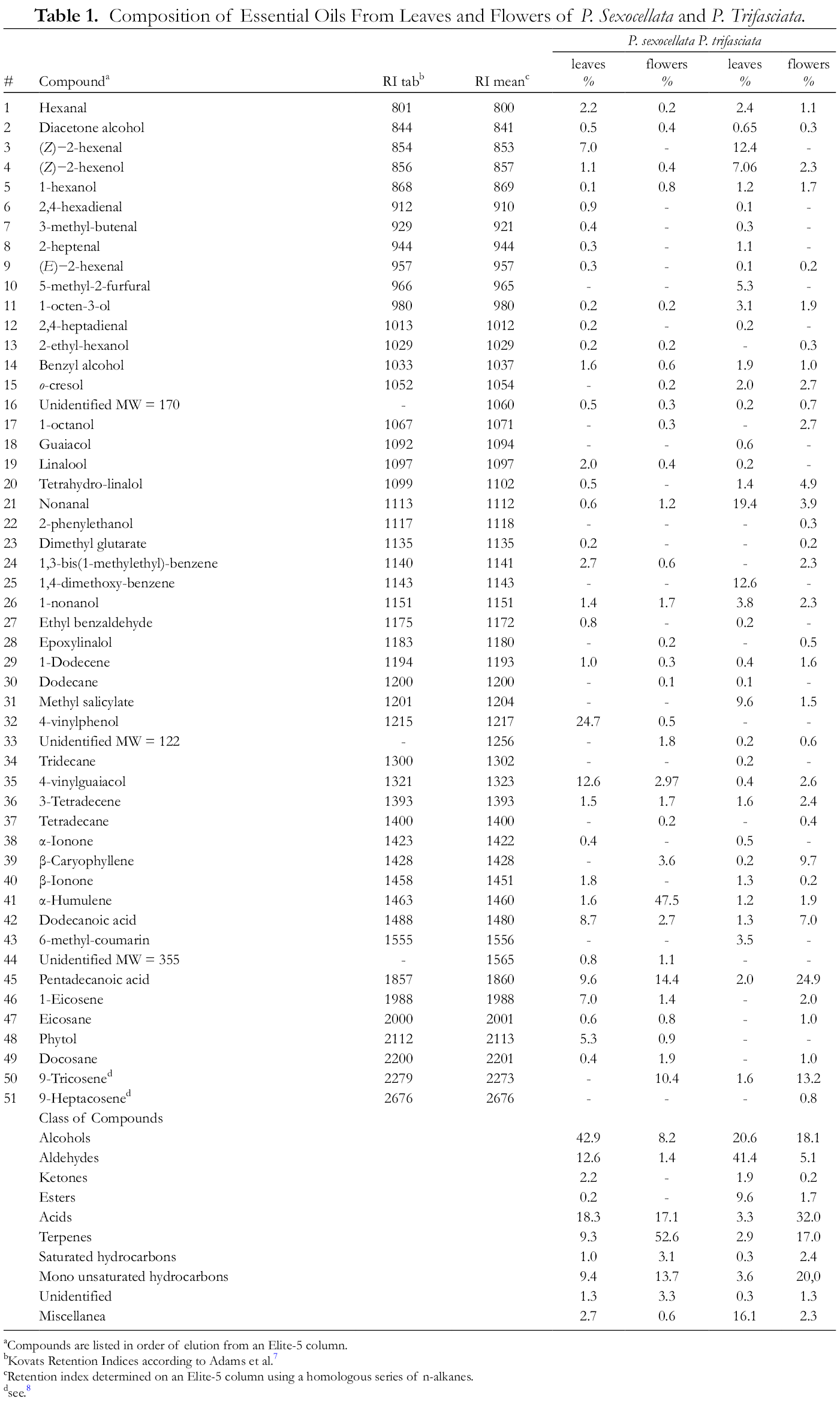
aCompounds are listed in order of elution from an Elite-5 column.
bKovats Retention Indices according to Adams et al. 7
cRetention index determined on an Elite-5 column using a homologous series of n-alkanes.
dsee. 8
Passiflora Sexocellata
Leaves essential oil
The main bulk of constituents of the volatile fractions were found to be alcohols (42.9%), detected in the range of 0.2 to 24.7%, from which 4-vinylphenol (24.7%), 4-vinylguaiacol (12.6%) and benzyl alcohol (1.6%) are the most abundant compounds. Organic acids accounting for 18.3% of the total essential oil represent the second largest class. This class is dominated by pentadecanoic acid (9.6%) and dodecanoic acid (8.7%). Aldehydes are present at 12.6%, from which (Z)-2-hexenal (7.0%) and hexanal (2.2%) are the most representative compounds. Mono unsaturated hydrocarbons accounting for 9.4% of the total essential oil are represented by 1-eicosene (7.0%) followed by 3-tetradecene (1.5%).
Flowers essential oil
The main bulk of constituents of the volatile fractions were found to be terpenes (52.6%), detected in the range of 0.2 to 47.5%, and dominated by α-humulene (47.5%) followed by β-caryophyllene (3.6%). Organic acids (17.1%) characterize the second largest class where pentadecanoic acid (14.4%) and dodecanoic acid are the most abundant compounds. Mono unsaturated hydrocarbons (13.7%) are represented by 9-tricosene (10.4%) and 3-tetradecene (1.7%). Alcohols are represented by 4-vinylguaiacol (3.0%) and 1-nonanol (1.7%).
Passiflora Trifasciata
Leaves essential oil
The main bulk of constituents of the volatile fractions were found to be aldehydes (41.4%), detected in the range of 0.1 to 19.4%, from which nonanal (19.4%), (Z)-2-hexenal (12.4%) and 5-methyl-2-furfural (5.3%) are the most abundant constituents. Alcohols (20.6%) represent the second largest class and (Z)-2-hexenol (7.0%), 1-nonanol (3.8%), 1-octen-3-ol (3.1%) and o-cresol (2.0%) are the most representative compounds. Esters class is dominated by methyl salicylate (9.6%). Mono unsaturated hydrocarbons (3.6%) are represented by 9-tricosene (1.6%) and 3-tetradecene (1.6%). Organic acids, accounting for 3.3% of the total essential oil are represented by pentadecanoic acid (2.0%) and dodecanoic acid (1.3%). 1,4-dimethoxy-benzene (12.6%) and 6-methyl-coumarin (3.5%) were also detected.
Flowers essential oil
The main bulk of constituents of the volatile fractions were found to be organic acids (32.0%), represented by pentadecanoic acid (24.9%) and dodecanoic acid (7.0%), respectively. The second largest class is represented by mono unsaturated hydrocarbons (20.0%), from which 9-tricosene (13.2%), 3-tetradecene (2.4%), 1-eicosene (2.0%) and 1-dodecene (1.6%) are the most abundant compounds. Alcohols account for 18.1% of the total essential oil are represented by 1-octanol (2.7%), o-cresol (2.7%), 4-vinylguaiacol (2.6%), 1-nonanol (2.3%), (Z)-2-hexenol (2.3%), 1-octen-3-ol (1.9%), 1-hexanol (1.7%) and benzyl alcohol (1.0%). Terpenes (17.0%) are featured by β-caryophyllene (9.7%), followed by tetrahydro-linalol (4.9%) and α-humulene (1.9%). Aldehydes are represented by nonanal (3.9%).
The highest intraspecific differentiation level of subgenus Decaloba was studied by Pérez et al., (2017) by using qualitative descriptors of 61 Passiflora L. species. This investigation showed morphological variations when subgenus Decaloba was compared to the Passiflora and Astrophea subgenus. 9 In addition, a comparison between cytogenetic and morphological features with pollinator syndrome underlined different evolutionary time scales. To this regard, the supersection Decaloba of the subgenus Decaloba showed the highest divergence among plant-pollination adaptation mechanisms. Although the whole subgenus is characterized by a reduced flowers size, the pollination by small insects (i.e., wasps and small bees) is predominant. For other species, such as P. trinervia and P. penduliflora that are pollinated by hummingbirds and bats, different and extreme mutualistic specializations between flower and pollinator are reported. 10 Regarding the floral structure, other mechanisms (e.g., floral scent chemistry) play an important role both in controlling flower visiting and in the foraging behavior of insects. This process is considered as the major specialization that provides the mutualistic relationship plant-pollinator.
The significant increase in organic acids (31.9%) and mono unsaturated hydrocarbons (20.0%) detected in flower of P. trifasciata could attest a different insect attraction mechanism, in comparison with P. sexocellata that highlighted lower percentages - 17.1% and 13.7% respectively - for the examined compounds. Simultaneously, the opposite trend observed by comparing the leaf extracts of the two plant species (higher amount of both organic acids (18.3%) and mono unsaturated hydrocarbons (9.4%) in leaves of P. sexocellata) suggested an involvement of these compounds in the reproductive mechanism commonly due to the flower traits.
Despite the absence of any physiological evidences concerning the nectar dynamics and reproductive biology of P. trifasciata, our findings support the hypothesis of a divergence in the scent production related to a different pollinator syndrome.
Regarding Passiflora family and its nectar production dynamic, high rates of nectar like the storage of starch in P. actinia (subgenus Passiflora) have been shown to be associated with structural characteristics of nectaries and pollinator behavior and breeding strategies. 11 For P. biflora (supersection Decaloba) starch storage was also described by Durkee et al. (1981). 12 The high content in total acid and alcohols recorded in P. trifasciata, could be considered as an additive mechanism playing a role in the selective pressures exerted on the pollinator species, towards larger and long-distance flight capable bees. On the other hand, P. sexocellata belonging to a different taxonomical supersection (Cieca) co-evolved a close association with a limited number of herbivores - only one or two known- and limited its pollination to small or medium guild bees. 3 Concerning our findings, a conspicuous content in terpenes (52.6%) was recorded in the flower extracts of P. sexocellata, associated to a reduced amount of total acids and alcohols (17.1% and 8.2%, respectively), comparing to P. trifasciata and to a complete absence of detectable ketones and esters. On this regard, Porter-Utley (2014) reports the absence of flavonoid compounds as a distinctive feature of most species of supersection Cieca, analyzing dried leaf samples. 3 Accordingly, the absence of ketones and esters in the volatiles of the flower fraction should be further investigated as a distinctive trait of P. sexocellata, including additional species within the same supersection. Terpenes are a large class of compounds usually detected in the volatile constituents of essential oils of different plant species. 13 Several studies investigating the complex pollination mechanisms of orchids have largely highlighted the involvement of terpenes in the pollination by sexual deception. 14 This latter is a highly specialized pollination strategy that mainly mimic the sex pheromone of females of their pollinator species to attract male insects that then pollinate the flowers. 15 Although considerable progress, many gaps remain in the understanding of the terpene’s biosynthesis, but evidence was provided on the attraction exercised by short-chain terpenes on pollinators, that seem to be attracted from distance to their mates. 14 Different kinds of chemicals, other than terpenes are involved in this mechanism (cyanogenic glucosides, alkaloids, flavonoids, saponins and tannins). On the contrary, chemical mimicry is widely known in different Passiflora species as a coevolutive process occurring between passion flower and heliconiines, in terms of a protection system evolved by the plant against its predators. 16 In particular, the high terpenes content detected in the volatile fraction of flower extracts of P. sexocellata was ascribable to a high content in α-humulene. On this respect, high level of (E)-caryophyllene and α-humulene, were reported by Da Silva et al. (2015) in Commiphora leptophloeos leaf oil, and the oviposition deterrent activity of the two terpenes were investigated, suggesting an insecticides role for those chemicals. 17 However, interactions with environment and insects in plants, promoted by terpenes and terpenoids, involve a myriad of mechanisms, underlining their important role as chemical mediators. 18 In the contest of the present paper, the high amount of terpenes detected in P. sexocellata suggests a mechanism aimed to control the flower visitors, both repelling the presence of undesired predators and as a selective approach promoting specific coevolved pollinators.
Conclusions
The present paper provides the first qualitative and quantitative characterization of the essential oil volatile fraction isolated from fresh leaves and flowers of P. trifasciata and P. sexocellata. Terpenes, organic acids and unsaturated hydrocarbons represent the major constituents of flowers by acting as semiochemicals to attract insect pollinators. These compounds are deemed to be crucial for P. sexocellata and P. trifasciata reproduction. Further investigations of the involvement of these compounds in pollinators’ interaction mechanism might be of great interest.
Experimental
Plant Material
Passiflora trifasciata Lem. and Passiflora sexocellata Schltdl. are part of a greenhouse-grown plant collection including wild species, varieties and hybrids, belonging to the subgenera Passiflora and Decaloba, was established in Sanremo (NW-Italy) UTM WGS84, Zone 32T North: Long 400142.51 Lat 4852298.79. The plants were grown in 30 cm pots supplied with plastic structures to allow climbing of the tendrils and they were heated during winter and shadowed during summer time, with temperatures ranging from 10 °C (min) to 32 °C (max). At the flowering stage, flowers and young leaves produced in the current year of the same phenological stage and without any defect were collected at about 9.30 am, put into Pyrex 250 ml sealed bottles and stored at −80 °C until analyzed. A voucher specimen was deposited at CREA-OF (Sanremo, Italy) with code PASS017 and PASS027, respectively. The specimen was fully characterized by morphological characters and by the ISSR markers. 19
Isolation of Essential Oil
samples of flowers and leaves of P. sexocellata (7.12 g and 18.67, respectively) and P. trifasciata (8.55 g and 20.52, respectively) to which octyl octanoate was added as internal standard, were steam distilled for 3 hours. Steam distillation was performed in the steam distillation system. Briefly, a body for the distillation was made of heat-resistant glass, including a plate of stainless steel, and the steam passing through the plants was water-cooled, thereby producing the essential oil and plant water simultaneously. The distillate was extracted three times with 100 ml of methylene chloride, dried over anhydrous Na2SO4 concentrated with a rotary evaporator and finally using a gentle stream of N2, stored at −20 °C until analyzed.
Fractionation and Alkylthiolation of Alkenes
after analyzes of the whole essential oil, a portion from each sample was subjected to selective purification process 8 and alkylthiolation reaction. 20 The dimethyl disulfide adducts were identified, and the positions of the methyl sulfide substituents were deduced from the fragmentation pattern.
GC-FID Analysis
The analyzes were carried out using a Hewlett Packard model 5980 GC, equipped with Elite-5MS (5% phenyl methyl polysiloxane) capillary column of (30 m × 0.32 mm i.d.) and film 0.32 µm thick. The carrier gas was He at a flow of 1 ml/min. An aliquot (1 µL) of essential oil were manually injected in “split” mode (30:1). The oven temperature program included an initial isotherm of 40 °C for 5 minutes, followed by a temperature ramp to 260 °C at 4 °C/min, and a final isotherm at this temperature for 10 minutes. Injector and detector temperatures were set at 250°C and 280 °C, respectively. The relative amount of each component was calculated based on the corresponding FID peak area without response factor correction.
GC-MS Analysis
The analyzes were carried out using a GC Model 6890 n, coupled to a bench top MS Agilent 5973 Network, equipped with the same capillary column and following the same chromatographic conditions used for the GC/FID analyzes. The carrier gas was He at constant flow of 1.0 mL/min. The essential oils were diluted prior to analysis and 1.0 µl was manually injected into the GC system with a split ratio of 30:1. The ion source temperature was set at 200 °C, while the transfer line was at 300 °C. The acquisition range was 40‐500 amu in electron-impact (EI) positive ionization mode using an ionization voltage of 70 eV.
Identification of the Components of the Volatile Fractions
The components of the essential oils were identified by comparing their mass spectra with NIST 98 and Wiley 5 MS libraries, as well as by comparing their Kovats retention indices, relative to a C8-C23 homologous series of n-alkanes and calculated according to Van Den Dool and Kratz, 21 with literature values. 7
Footnotes
Acknowledgments
Research partially funded by the project “Implementation of the FAO International Treaty on Plant Genetic Resources for Food and Agriculture” of the Ministry of Agriculture, Alimentation and Forestry Policies, aimed at research and experimentation supporting the collection, characterization and evaluation of plant genetic resources. The experimental work for this study has been performed within the framework of the master’s degree thesis of Mr. Filippo Testa (Bachelor of Science in Pharmacy, University of Pavia).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.