
Abstract
Fucoidans are widely used as an ingredient of dietary supplements. We investigated the immune stimulatory activities of Undaria pinnatifida (Alariaceae) fucoidan-rich extract (UPF-RE) in vitro as well as in vivo. In vitro, the extract stimulated Raw 264.7 cells to produce significant nitric oxide (NO) metabolites and cytokines (TNF-α, IL-1α, IL-1β, and IL-6). It also induced the proliferation of primary mouse splenocytes and the secretion of IL-4, which correlated with the phosphorylation of Extracellular-signal-regulated kinase (ERK) protein. In in vivo experiments, first, 50 mg/kg of 3 different types of UPF-RE, DSU02, DSU02L (low molecular weight, <3 kDa), and DSU02H (high molecular weight, >10 kDa), were orally administered to C57BL/6 mice. After 14 days, the frequencies of CD3+, CD4+, and CD8+ T cells and NK cells from each group were analyzed. Plasma concentrations of TNF-α and IFN-γ were determined. The frequencies of CD3+ and CD4+ showed a statistically significant increase in splenocytes isolated from the DSU02 and DSU02H groups. Also, there was significant production of TNF-α and IFN-γ from the DSU02 group. Second, 3 different concentrations of DSU02 (50, 100, and 150 mg/kg) were orally administered. After 14 days, the proliferative capacity of CD3+, CD4+, and CD8+ T cells was investigated, and the plasma concentrations of IgM and total IgG were determined. Plasma concentration of IgM from the DSU02 150 mg/kg group was statistically significantly higher compared with that from the other groups. We suggest that UPF-RE could be a good candidate for a natural immune stimulator to induce innate as well as adaptive immune responses.
Fucoidans (sulfated polysaccharides) are known to exist mostly in marine brown algae. A large part of the structure of fucoidans consists of fucose and sulfated polysaccharides with additional sugar constituents such as mannose, galactose, glucose, xylose, and uronic acids. 1 These products have been reported to possess a variety of biological properties such as immune stimulatory, anti-inflammatory, antiviral, and antitumor activities. 2 -4 Especially, fucoidan-containing polysaccharide structures have been known to serve as a potent immune stimulator. 5 However, the immune modulatory activities of fucoidans are quite different based on the type of extract, algal species from which the fucoidans are derived, molecular weight, the content of fucoidans, and the content of other components. 6,7 In a previous study, high- and low-molecular weight fucoidans presented clearly opposite effects on the induction of immune responses. 8 Therefore, the biological activity of all fucoidan-related products should be evaluated and standardized before being made commercially available as dietary supplements because different components of fucoidans have caused confusion in understanding their exact mechanism in terms of immunomodulation.
In this study, we extracted fucoidan-rich fractions from Korean Undaria pinnatifida collected from Jeju-island. Undaria pinnatifida derived fucoidans were reported to have a high sulfate and l-fucose content 9 and showed a wider range of biological activities than other fucoidans. Our study identified the immune stimulatory effect of an U. pinnatifida fucoidan-rich extract and compared the activities of different types of fucoidan-rich extract.
First, we utilized either Raw 264.7 cells or primary splenocytes to investigate the effect of U. pinnatifida fucoidan-rich extract (UPF-RE) in terms of innate and adaptive immunity. The production of NO metabolites and pro-inflammatory cytokines was measured in Raw 264.7 cells stimulated with different concentrations of UPF-RE; there was significant production of nitrite/nitrate in Raw 264.7 cells treated with the extract (50 and 125 µg/mL) and Lipopolysaccharide (LPS) (1 µg/mL) compared with that of an untreated control (Figure 1(a)). Also, there was a dramatic increase of Tumor necrosis factor- α (TNF-α) and Interleukin-6 (IL-6) production in Raw 264.7 cells treated with all concentrations of the fucoidan-rich extract (Figure 1(b), top panel). In addition, there was a significantly greater secretion of both IL-1α and IL-1β in Raw 264.7 cells treated with UPF-RE (50 and 125 µg/mL for IL-1α; 125 and 250 µg/mL for IL-1β) compared with that in the untreated control. Several studies have examined the immunomodulatory effect of either fucoidan or fucoidan-rich extract by using Raw 264.7 cells. Teruya et al reported that 2 different fucoidans isolated, respectively, from Cladosiphon okamuranus and Laminaria angustata var. longissima stimulated the production of NO, TNF-α, and IL-6 in Raw 264.7 cells. They also mentioned that both sulfate and acetyl groups of fucoidan are involved in the activation of the Raw 264.7 cells. 10,11 Jin et al reported that fucoidan from U. pinnatifilda induced the production of IL-6, IL-8, and TNF-α in human neutrophils through a PI3/AKT signaling pathway. 12 Meanwhile, Sanjeewa’s group reported the anti-inflammatory effect of sulfated polysaccharide (fucodians) isolated from Sargassum horneri. 13 Also, Lee’s group showed that Ecklonia cava fucoidan significantly inhibited NO production in LPS-induced Raw 264.7 macrophage cells by inhibiting the expression of iNOS, COX-2, and pro-inflammatory cytokines. 14 Sanjeewa’s group used a crude polysaccharide-rich extract, but the precise composition of the extract was not specified. Lee et al demonstrated that their fractionated fucoidans contained large amounts of carbohydrates (47.1%-57.1%), but they did not mention the level of fucosterol contamination in their fractions. Fucosterol is a steroidal compound reported to have a strong anti-inflammatory activity. 15 Therefore, it is important to analyze the exact composition of the fucoidan-rich extract. The U. pinnatifida fucoidan-rich extract used in this study had little contamination with fucosterol (0.25% ± 0.01%), as mentioned in material and methods. Therefore, we propose that U. pinnatifida fucoidan-rich extract induces an innate immune activation in Raw 264.7 cells.

Effect of Undaria pinnatifida fucoidan-rich extract on the activation of Raw 264.7 cells. Raw 264.7 cells were incubated with different concentrations of either Undaria pinnatifida fucoidan-rich extract (0, 50, 125, and 250 µg/mL) or Lipopolysaccharide (LPS) (1 µg/mL) for 24 hours and culture supernatants were harvested. (a) Production of nitrite/nitrate. (b) Production of TNF-α, IL-6, IL-1α, and IL-1β, measured by enzyme-linked immunosorbent assay.
The proliferation, IL-4 secretion, and expression of immune activation-linked molecules (Nuclear factor kappa-light-chain-enhancer of activated B cells (NK-κB), pNK-κB, Extracellular-signal-regulated kinase (ERK), and pERK) were examined in primary mouse splenocytes treated with UPF-RE. This induced significant proliferation of splenocytes in a dose-dependent manner (Figure 2(a)), which clearly correlated with increased expressions of pERK (Figure 2(c) and (d)). Also, IL-4 production in primary splenocytes was significantly increased by all 3 concentrations of the extract (Figure 2(b)), whereas neither IL-2 nor IL-13 was detected (data not shown). While the expression of pERK increased in a dose-dependent manner along with the activation of splenocytes, there was little change in the expression of NK-κB, pNK-kB, and ERK (Figure 2(c)). Fucoidans can be prepared from a variety of brown seaweeds, including Ascophyllum nodosum, Macrocystis pyrifera, and U. pinnatifida. The biological effects of fucoidan extracts seem to be differentiated in accordance with the origin of the extract, fucoidan content, and other molecules that are contained. 6,7 In particular, fucoidan from U. pinnatifida showed a strong activation of spleen T cells in comparison with fucoidans from either A. nodosum or M. pyrifera. 6 Vetvicka reported that only pure fucoidan showed a significant proliferation effect on either T cells or B cells compared with other fucoidan-rich extracts in a complex with other substances, which clearly indicates that the U. pinnatifida fucoidan-rich extract used in this study has been prepared with a high fucoidan content. 7 Recently, U. pinnatifida sporophyll-derived fucoidans showed an anticancer effect by inhibiting the phosphorylation of Mitogen-activated protein kinase (MAPK)-related molecules, including pERK in mouse hepatocarcinoma. 16,17 Meanwhile, other studies reported an enhancing effect of either fucoidans or fucoidan-rich extracts on the MAPK pathway, which is closely associated with activation of immune cells or normal cells. 18,19 In this study, we provide the first report that the stimulation of splenocytes by U. pinnatifida fucoidan-rich extract is mediated by the induction of pERK expression (Figure 2(c) and (d)).
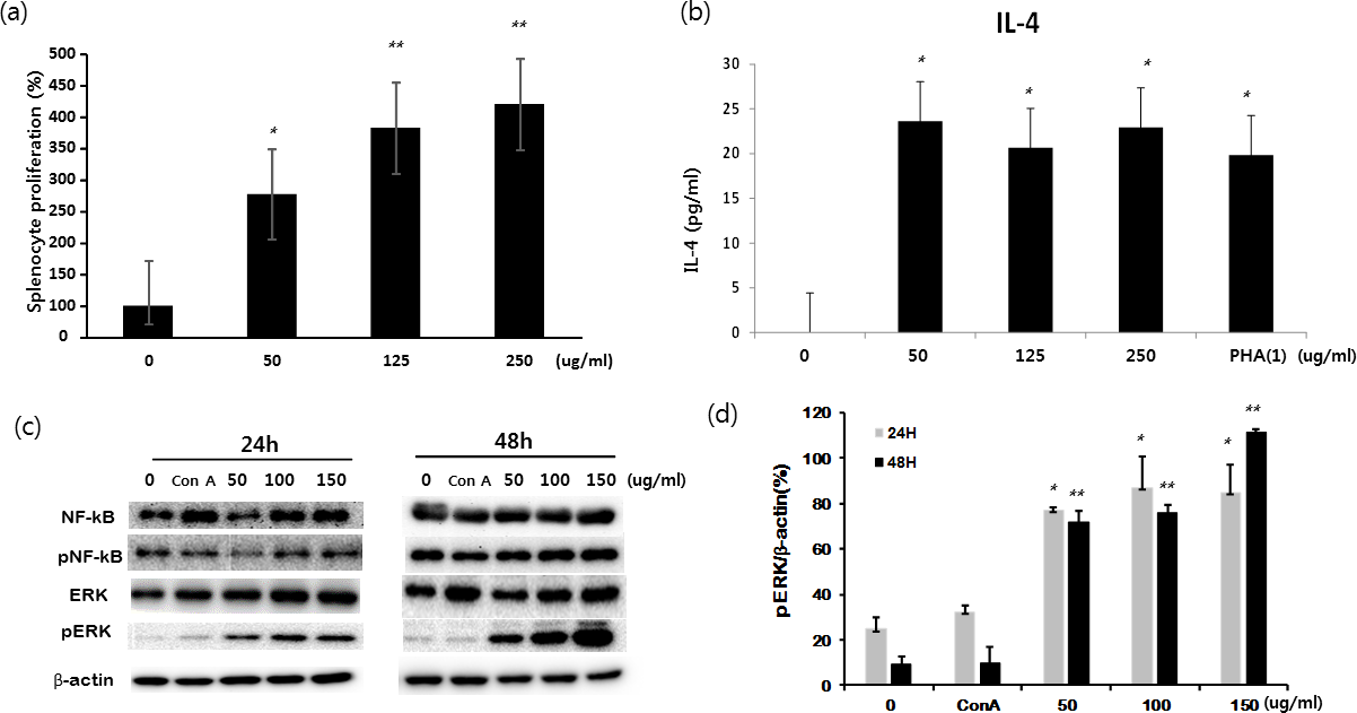
Effect of Undaria pinnatifida fucoidan-rich extract on the activation of splenocytes. Primary splenocytes were isolated from C57BL/6 mice and cultured with different concentrations of either Undaria pinnatifida fucoidan-rich extract (0, 50, 125, and 250 µg/mL) or phytohemagglutinin (1 µg/mL). (a) Proliferation of splenocytes measured by CCK-8 assay at 72 hours after incubation. (b) Production of IL-4 measured by enzyme-linked immunosorbent assay at 24 hours after incubation. (c) Splenocytes were cultured with different concentrations of either Undaria pinnatifida fucoidan-rich extract (0, 50, 100, and 150 µg/mL) or concanavalin A (1 µg/mL). Protein expressions of pERK and NFκB were analyzed by Western blot analysis at 24 hours after incubation.
Second, 50 mg/kg of either 3 different types of U. pinnatifida fucoidan-rich extract, DSU02, DSU02L (low molecular weight, <3 kDa), and DSU02H (high molecular weight, >10 kDa), or 3 different concentrations of DSU02 (50, 100, and 150 mg/kg), were orally administered in different experimental sets. After 14 days, the frequencies of CD3+, CD4+, and CD8+ T cells and NK cells and the proliferation of each subpopulation were analyzed by flow cytometry. In addition, the plasma concentrations of TNF-α, IFN-γ, IgM, and total IgG were determined. There was little change in either whole body weight or liver or spleen weight among 5 different treatment groups after 14 days (Figure 3(a) and (b)). The percentages of CD3+ T cells significantly increased in the DSU02H- and DSU02-treated groups compared with the other groups (Figure 4(a)). Only the DSU02H- and red ginseng (RG) extract-treated groups upregulated the percentages of CD8+ T cells, while the DSU02H-, DSU02-, and RG extract-treated groups showed a significant increase in the percentage of CD4+ T cells compared with the other groups (Figure 4(b)). However, there were no differences in the frequency of NK cells among the groups (Figure 4(c)). In fact, other studies have previously reported that the percentages of CD3+ and CD4+ T cells increased in an animal group treated with a high-molecular weight U. pinnatifida fucoidan-rich extract, but not in a group treated with a low-molecular weight U. pinnatifida fucoidan-rich extract, which partly agrees with our data. 8,20 Whole blood was drawn on day 14 after treatment, and the amounts of TNF-α and Interferon-γ (IFN-γ) in plasma were measured using enzyme-linked immunosorbent assay (ELISA). Both TNF-α and IFN-γ, the typical immune activating cytokines, were significantly induced from animals treated with either DSU02 or RG extract (Figure 5(a) and (b)). However, neither DSU02L nor DSU02H was able to stimulate the production of either TNF-α or IFN-γ (Figure 5(a) and (b)). Our study shows that only in the DSU02-administered group, but not in the DSU02H-administered group, was a significant amount of TNF-α and IFN-γ secreted (Figure 5(a) and (b)), which agrees with a report by Maruyama’s group, in which Mekabu fucoidan derived from sporophylls of U. pinnatifida increased IFN-γ and suppressed Th2 responses. 21 We speculate from the results of both Mekabu fucoidan extract and our DSU02 that the crude UPF-RE may have additional components that have a potent immunodominant property toward the Th1 response. Therefore, we also examined whether DSU02 had either other immune enhancing effects or a dose-dependent response in vivo.

In vivo effect of Undaria pinnatifida fucoidan-rich extract on body and organ weight changes. Mice were orally administered drinking water and either one of three different types of Undaria pinnatifida fucoidan-rich extract (DSU02L 50 mg/kg, DSU02H 50 mg/kg, or DSU02 50 mg/kg) or red ginseng extract (50 mg/kg). (a) Whole body weight changes were monitored daily until the experimental end point. (b) Liver or spleen weight changes.

In vivo effect of Undaria pinnatifida fucoidan-rich extract on the ratio of T cell population. Mice were orally administered drinking water and either one of three different types of Undaria pinnatifida fucoidan-rich extract (DSU02L 50 mg/kg, DSU02H 50 mg/kg, or DSU02 50 mg/kg) or red ginseng extract (50 mg/kg). Spleen was isolated from each animal on day 14 after treatment. A single cell suspension of splenocytes was prepared and the percentages of (a) CD3+ T, (b) CD8+ T and CD4+ T, and (c) NK+ were analyzed using flow cytometry.
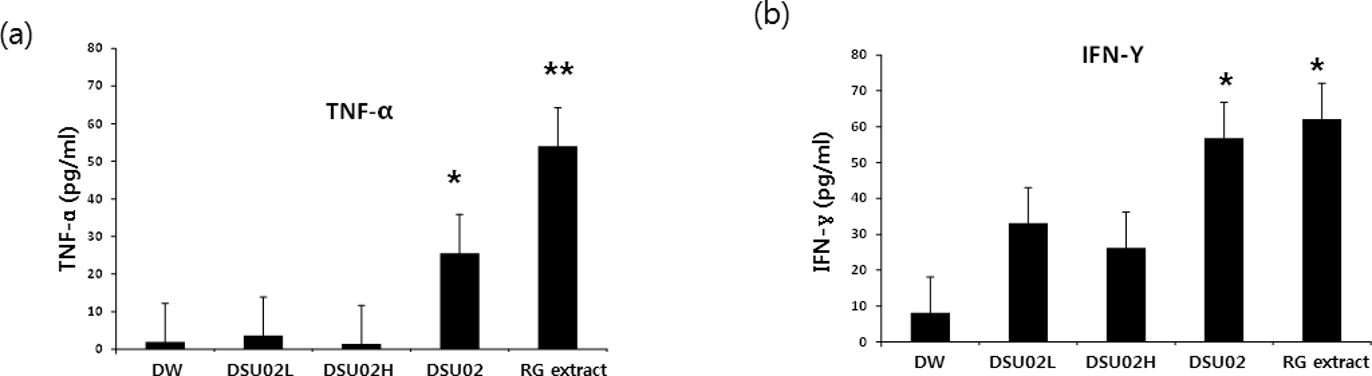
In vivo effect of Undaria pinnatifida fucoidan-rich extract on the production of TNF-α and IFN-γ. Mice were orally administered drinking water and either one of three different types of Undaria pinnatifida fucoidan-rich extract (DSU02L 50 mg/kg, DSU02H 50 mg/kg, or DSU02 50 mg/kg) or red ginseng extract (50 mg/kg). Whole blood was drawn on day 14 after treatment, and production of TNF-α (a) and IFN-γ (b) was measured using enzyme-linked immunosorbent assay.
Finally, the proliferation of CD3+, CD4+, and CD8+ T cells was measured through the analysis of splenocytes by carboxyfluorescein succinimidyl ester (CFSE) staining, and also the production of IgM and total IgG was identified. The CD4+ T cells from either the DSU02- or RG extract-administered group tended to proliferate more than those from the DW group, whereas the proliferative capacity of CD8+ T cells did not differ among the groups (Figure 6(a) and (b)). Plasma IgM showed a significant increase with oral administration of either DSU02 (150) or RG extract (Figure 7(a)). Also, total IgG in blood from animals administered with either DSU02 (100) or DSU02 (150) was higher than that from either the DW group or RG extract group, but the result was not statistically significant (Figure 7(b)). Our data for IgM and total IgG partly agree with those from the Takai study, but there are some variables that are not comparable between the in vitro and in vivo experiments. 22

In vivo effect of Undaria pinnatifida fucoidan-rich extract on the proliferation of CD3+, CD4+, and CD8+T cells. Mice were orally administered drinking water and either one of three different types of Undaria pinnatifida fucoidan-rich extract (DSU02 50 mg/kg, DSU02 100 mg/kg, or DSU02 150 mg/kg) or red ginseng extract (50 mg/kg). Spleen was isolated from each animal on day 14 after treatment. A single cell suspension of splenocytes was stained with carboxyfluorescein succinimidyl ester and cultured for another 7 days, and proliferations of CD3+ T, CD4+ T, and CD8+ T cells were analyzed using flow cytometry. Representative Flow cytometer (FACS) plots show CD3+, CD4+, and CD8+ expression (a) and specific percentages of CD3+ T, CD4+ T, and CD8+ T cells (b).

In vivo effect of Undaria pinnatifida fucoidan-rich extract on the production of IgM or total IgG mice were orally administered drinking water and either one of three different types of Undaria pinnatifida fucoidan-rich extract (DSU02 50 mg/kg, DSU02 100 mg/kg, or DSU02 150 mg/kg) or red ginseng extract (50 mg/kg). Whole blood was drawn on day 14 after treatment, and production of IgM (a) and total IgG (b) was measured using enzyme-linked immunosorbent assay.
In summary, our study provides a comprehensive understanding of the immune-stimulatory effect of an U. pinnatifida fucoidan-rich extract in vitro as well as in vivo. Especially, we report that the activation of splenocytes by this extract is strongly associated with the induction of pERK expression.
Experimental
Preparation of Undaria pinnatifida Fucoidan-Rich Extract
Undaria pinnatifida was collected in April 2017 from Gonae-ri, Jeju-island, Republic of Korea. The seaweed was washed repeatedly 3 times with purified water to remove salt and then air-dried for 12 hours, powdered and extracted with 0.1 N HCl (v/v) for 12 hours at 45°C for acid hydrolysis. The hydrolysate was filtered using a decanter and the supernatant collected. The same volume of 95% (v/v) ethanol was added to the supernatant and soaked for 12 hours. The extract was filtered with a continuous-centrifuge to collect a pellet. The lyophilized pellet was obtained as an U. pinnatifida fucoidan-rich powder (yield, 5.1% (w/w)); the average molecular weight of fucoidan in this was 200 kDa. Fucosterol (reference from Ecklonia cava extract) was estimated to form 0.25% ± 0.01% (w/w). The extracted intact fucoidans mostly consisted of carbohydrates (78.8%) with protein (1.2%), salt 1.4%, water 5.3%, and others 13.3%. Carbohydrates included fucose 23.38%, glucose 1.93%, and others 3.76 %, such as rhamnose, galactose, and xylose (provided by Seojinbiotech Co., Ltd.). The lyophilized pellet of DSU02 was further processed for microorganism fermentation, and the resulting extract was divided into low molecular weight (<3 kDa) and high molecular weight (>10 kDa) fractions by ultrafiltration. The lyophilized pellets of DSU02, DSU02L, and DSU02H were dissolved in dimethyl sulfoxide (Sigma-Aldrich, St. Louis, MO, United States), filtered at 0.22 µm, and stored at −20°C until use. All samples were diluted with distilled water for oral administration in the mouse experiments.
Cell Cultures
Raw 264.7 cells were cultured in Dulbecco’s modified Eagle’s medium (Gibco, United States) supplemented with 10% heat-inactivated fetal bovine serum (FBS, Hyclone, United States). Primary splenocytes isolated from C57BL/6 mice were cultured in Roswell Park Memorial Institute (RPMI) 1640 with l-glutamine (Corning, United States). All cells were cultured in the presence of 100 U/mL penicillin and streptomycin (Gibco, Grand Island, NY, United States) at 37°C in a humidified atmosphere with 5% CO2.
Nitric Oxide Metabolites Detection
Cell free supernatants were harvested to measure the production of nitric oxide metabolites (nitrite/nitrate) using a Nitrite/Nitrate Assay kit (Sigma-Aldrich, MO, United States). Absorbance was measured at 540 nm using a microplate reader (BMG Labtech, Ortenberg, Germany).
Cell Proliferation Assay
The cell proliferative effect of UPF-RE extract on primary mouse splenocytes was assessed by using a Cell Counting Kit-8 (CCK-8, Dojindo, Japan). Cells were seeded at a density of 1 × 104 cells per well in a 96-well flat bottom plate and different concentrations of U. pinnatifida fucoidan-rich extract (0, 50, 125, and 250 µg/mL) were added for 48 hours. Either phytohemagglutinin (1 µg/mL) or concanavalin A (1 µg/mL) was used as positive control. Then, CCK-8 solution was added to each well, and the cells incubated in accordance with the reactive time of the solution. The absorbance was measured using a microplate reader (BMG Labtech, Ortenberg, Germany) at 450 nm.
Enzyme-Linked Immunosorbent Assay
Cell free supernatants were harvested to measure the production of TNF-α, IL-6, IL-1α, IL-1β, IL-4, and IFN-γ using a mouse ELISA kit (BD Biosciences, NJ, United States). Absorbance was measured at 450 nm using a microplate reader (BMG Labtech, Ortenberg, Germany).
Western Blot
Primary mouse splenocytes were cultured with different concentrations of UPF-RE (0, 50, 100, and 150 µg/mL) for 24 hours. Concanavalin A was used as a positive control. Cells were lysed by protein extraction buffer (Intron Biotechnology, Korea). Proteins in cell lysates were quantified by the Bradford assay, separated by electrophoresis, and transferred to nitrocellulose membranes, which were then blotted with first and second antibodies. Anti-NFκB (#3114, MillIpore, United States), anti-p NFκB (#3036s, Cell Signaling, United States), anti-ERK (#9102s, Cell Signaling, United States), and anti-pERK (#4370s, Cell Signaling, United States) were used for first antibodies. The blots were visualized by an enhanced chemiluminescent detection solution (Intron Biotechnology, South Korea), as previously described. 14
Oral Administration of Undaria pinnatifida Fucoidan-Rich Extract
All the animal experiments were conducted in accordance with the recommendations in the National Research Council’s Guide for the Care and Use of Laboratory Animals. The experimental protocol was approved by the Animal Experiments Committee of Duksung Women’s University (permit number: 2018-003-008). Two sets of animal experiments were organized. The first set was designed to investigate the immune-stimulatory effect of DSU02L 50 mg/kg, DSU02H 50 mg/kg, and DSU02 50 mg/kg. C57BL/6 mice (female, 5-6 weeks old) were adapted for 1 week before the experiment. Mice were orally administered drinking water (DW) and either DSU02L 50 mg/kg, DSU02H 50 mg/kg, or DSU02 50 mg/kg, or RG extract (50 mg/kg). The second set of experiments was designed to examine the dose response of DSU02. Mice were divided into 5 groups (DW, DSU02 50 mg/kg, DSU02 100 mg/kg, DSU02 150 mg/kg, and RG extract 50 mg/kg; orally administered). Body weight changes were monitored during treatment, and liver and spleen were isolated from each animal on day 14 after treatment. Whole blood was drawn by cardiac puncture, and either plasma cytokines (TNF-α and IFN-γ) or antibodies (IgM and total IgG) were measured. Also, the percentages of NK cells and subpopulations (CD3+, CD4+, or CD8+) of T cells from spleen were measured.
Fluorescent Antibody and Cell Surface Antigen Staining
Splenocytes were stained with rat anti-mouse CD8-APC (BD Biosciences, NJ, United States), rat anti-mouse CD4 -PE (BD Biosciences, NJ, United States), rat anti-mouse CD3-PE (BD Biosciences, NJ, United States), and mouse anti-mouse NK-1.1-APC (BD Biosciences, NJ, United States) according to the manufacturer’s instructions. After staining, cells were analyzed by flow cytometry (Novocyte Flow Cytometer, ACEA Biosciences, United States). The positivity of CD8, CD4, and CD3 was determined by comparison with the defined cutoff values obtained with unstained control cells, as previously described. 15
Proliferative Response of CD3, CD4, and CD8 T Cells by Carboxyfluorescein Succinimidyl Ester
T cell proliferation was assessed using a CellTrace CFSE Cell Proliferation Kit (ThermoFisher, MA, United States). Briefly, lymphocyte cells were isolated from spleen of C57BL/6 mice, and then the cells were stained with CFSE (5 µM) staining solution diluted in DPBS for 20 minutes in a 37°C water bath. After addition of RPMI1640 (Corning, NY, United States) media with 10% heat-inactivated FBS (Yong-In, Republic of Korea) and 100 U/mL penicillin and streptomycin (Gibco, NY, United States), the CFSE-stained cells were incubated for 5 minutes. Then, the cells were centrifuged at 300×g for 5 minutes, the supernatants aspirated, and the CFSE-stained cells were plated with a density of 1 × 107 cells per well in a 6-well bottom plate and incubated for 5 days in 5% CO2 at 37°C. After 5 days, the cells were stained with rat anti-mouse CD3-PE (BD Biosciences, NJ, United States), rat anti-mouse CD4-PE (BD Biosciences, NJ, United States), and rat anti-mouse CD8-APC (BD Biosciences, NJ, United States). The stained cells were analyzed by flow cytometry (Novocyte Flow Cytometer, ACEA Biosciences, United States).
Statistical Analysis
Data were processed using Microsoft Excel, and the results are presented as mean ± standard deviation. Comparisons of several means were performed with one-way analysis of variance followed by Fisher’s Least Significant Difference as a post hoc test. Differences among groups were considered significant at a value of P < 0.05.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was part of the project titled "Development of Global Senior-friendly Health Functional Food Materials from Marine Resources", funded by the Ministry of Oceans and Fisheries, Korea.