
Abstract
Lipid droplets (LDs), cytosolic fat storage organelles, are emerging as major regulators of lipid metabolism, trafficking, and signaling in various cells and tissues. LDs are altered in cardiovascular and neuronal disorders, inflammation, obesity, and cancer. Flavonoids comprise different classes of molecules, characterized by a well-known antioxidant activity and a beneficial effect in several diseases. However, the cellular mechanism by which different classes of flavonoids improve health is poorly understood, in particular as far as LDs biogenesis is concerned. Here we used Drosophila melanogaster as a model system to investigate the effects of a selected group of flavonoids on larval tissues by examining LDs biogenesis. In our study, fruit flies were grown in xanthohumol-, isoquercetin-, and genistein-enriched food and larval tissues were analyzed using a LD marker. Total mRNA expression of two main enzymes (minotaur and midway) responsible for triacylglycerides synthesis was evaluated after treatments. Among the flavonoids analyzed, xanthohumol and isoquercetin resulted to be potent regulators of LDs biogenesis in a tissue-specific manner, inducing fat storage decrease in fat bodies and accumulation of LDs in nerves. Since LDs have been suggested to play a protective role against intracellular stress in nonadipocyte cells, our data support the hypothesis that some phytochemicals could act as strong modulators of LDs biogenesis in vivo. The knowledge of how different flavonoids act on lipid metabolism in different tissues can help to manage the use of phytochemicals with the aim of selectively ameliorating specific neuronal and metabolic diseases’ manifestations.
Flavonoids are the largest group of phenolic compounds in plants and are synthetized through phenyl propanoid pathway against microbial infection. 1 Numerous studies have shown that food and beverage containing flavonoids, such as fruits, vegetables, cereals, red wine, and green tea, can improve health and participate in the prevention of numerous diseases. 2,3 Moreover, emerging evidences suggest that flavonoids may have beneficial effects on cardiovascular diseases, neuronal disorders, inflammation, and obesity in animals as well as in humans. 4-8 This protective mechanism seems to be related to their free radicals scavenging activity, inhibition of enzymes activity (such as aldose reductase, xanthine oxidase, lipoxygenase, and cyclic nucleotide phosphodiesterases [PDEs]), and modulation of protein kinase (e.g., AMP kinase), lipid kinase signaling, and peroxisome proliferator-activated receptor pathways. 9,10 Several in vivo studies had revealed that flavonoids help to reduce body weight gain, food consumption, and fat accumulation. 11,12 In mammals, fat is stored in the adipose tissue, which is made principally of white and brown adipocytes. It is well known that selected flavonoids can induce lipolysis in adipose tissue, likely through inhibition of PDEs and antagonism of cyclic adenosine monophosphate degradation. 10
The lipids are stored as single or multiple droplets called lipid droplets (LDs). LDs are cellular specialized organelles that store neutral lipids in all living organisms composed by a core, containing mainly triacylglycerides (TAG) and sterol esters, enclosed by a phospholipid monolayer. 13 LDs are particularly important in tissues specialized for energy storage or lipid turnover, such as the adipose tissue, the liver, and the intestine, and accumulate in skeletal muscles and nervous system. 14-17 LDs not only provide substrates for energy metabolism and building blocks for membranes, but also play a pivotal role in various cellular pathways, such as protein trafficking, protein degradation, and modulation of nuclear receptors. 14 Moreover, LDs exhibit a protective function against oxidative damages caused by different stimuli leading to endoplasmic reticulum (ER) stress. 18-22 Simple animal models as Caenorhabditis elegans and Drosophila melanogaster have been used to study the effects of flavonoids-rich diet on longevity, locomotors activity, fertility, and lethality. However, only few studies were conducted to investigate the relationship between the antioxidant effect of flavonoids and LDs biogenesis at cellular and tissue levels. 23-25 How different classes of flavonoids modulate lipid metabolism and intracellular LDs in various tissues is poorly understood. On the basis of this considerations, the aim of this study was to evaluate the effects of flavonoids on cytosolic LDs content in Drosophila fat bodies and oenocytes (adipose- and liver-like larval tissues), and as well in tissues with high energy demands such as the central nervous system and muscles.
The effects of genistein (an isoflavone), xanthohumol (a prenilated flavonol), and isoquercetin (a flavonol) on LDs mobilization in fruit fly fat body were evaluated. Drosophila fat body is considered the analogous of white adipose tissue in humans. It is known that some flavonoids exhibit lipid-lowering activity and decrease LDs in adipocytes through lipolysis by different mechanisms reducing LDs’ number. 26-28
Fat tissues were dissected from control larvae and stained with a lipophilic dye (BODIPY 493/503) to detect neutral lipids (Figure 1a). In the fat body cells, all the flavonoids induced a decrease of the percentage of cytoplasm area occupied by LDs, from 35.15% of the control to 30.17%, 32.56%, 26.82%, and 29.04% in the genistein, xanthohumol, and isoquercetin exposure, respectively (Figure 1b). Genistein induced similar changes, but only the increase of the smaller LDs’ size (of about 6.4 µm) resulted statistically different (Figure 1c and d).

(a) Representative confocal images of larval fat bodies in control and treated animals. The compounds utilized were genistein (10 µM), xanthohumol (10 µM), and isoquercetin (10 µM). The LDs were visualized by using BODIPY 493/503. Scale bar = 50 µm. Percentage of fat body cell cytoplasm occupied by LDs was calculated (b). Quantification of LDs number (c) and diameter (d). (e) The results of the LDs’ size were plotted as box-and-whisker plot, showing the interquartile range, the median as horizontal bar, and the whiskers are the minimum and maximum values. Values are mean ± SD from at least 3 independent experiments. Significance was calculated using unpaired t-test (two-tailed) for LDs number and one-way analysis of variance with Tukey’s post hoc test for LDs size. P < 0.05* and P < 0.01** vs control. LD: lipid droplet.
The Drosophila specialized cells called oenocytes are considered the analogous of mammal hepatocytes and could be used as a model for lipid metabolism studies. 29 When LDs from fat bodies are mobilized in order to furnish energy, the LDs content in oenocytes increased. In addition, under starved conditions, stimulated lipolysis results in high levels of circulating lipids and subsequently causes strong LDs staining in oenocytes. To correlate the effect of lipid mobilization in the fat body after chronic flavonoids treatment, we analyzed LDs distribution changes in oenocytes (Figure 2a). As expected, LDs in control larvae were barely present, while their number increased upon treatments with almost all the flavonoids used, except genistein (Figure 2b). Indeed, xanthohumol and isoquercetin led to increase of LDs content and induced the formation of smaller LDs (Figure 2c and d). The enhancement of oenocyte LDs amount by xanthohumol (1.24 LDs number/µm2) and isoquercetin (0.44 LDs number/µm2) treatment suggests an active lipolysis and breakdown of LDs in the fat body, whereas genistein, at least at the concentrations used in this study, has no clear lipolytic effect.

(a) Representative confocal images of larval oenocytes in control and treated animals. The compounds utilized were genistein (10 µM), xanthohumol (10 µM), and isoquercetin (10 µM). The LDs were visualized by using BODIPY 493/503. Scale bar = 20 µm. Both number (b) and diameter (c) of LDs were measured. (d) The results of the LDs size were plotted as box-and-whisker plot, showing the interquartile range, the median as horizontal bar, and the whiskers are the minimum and maximum values. Significance was calculated using unpaired t-test (two-tailed) for LDs number and one-way analysis of variance with Tukey’s post hoc test for LDs size. P < 0.05* and P < 0.01** vs control. LD: lipid droplet.
In Drosophila, LDs are produced in glial cells and are demonstrated to be important modulators of neuronal function and maintenance. 30 In Drosophila wild-type larvae, all the treatments led to a statistically significant increase of LDs number (Figure 3a and b) without affecting (Figure 3c and d). We found that in control larvae were present 0.044 LDs/μm2, while in the larvae treated with genistein were observed 0.05 LDs/μm2. Moreover, the number of LDs upon xanthohumol and isoquercetin treatment was increased to 0.06 LDs/μm2. Our data show that glial cells are sensitive to flavonoids supplementation.
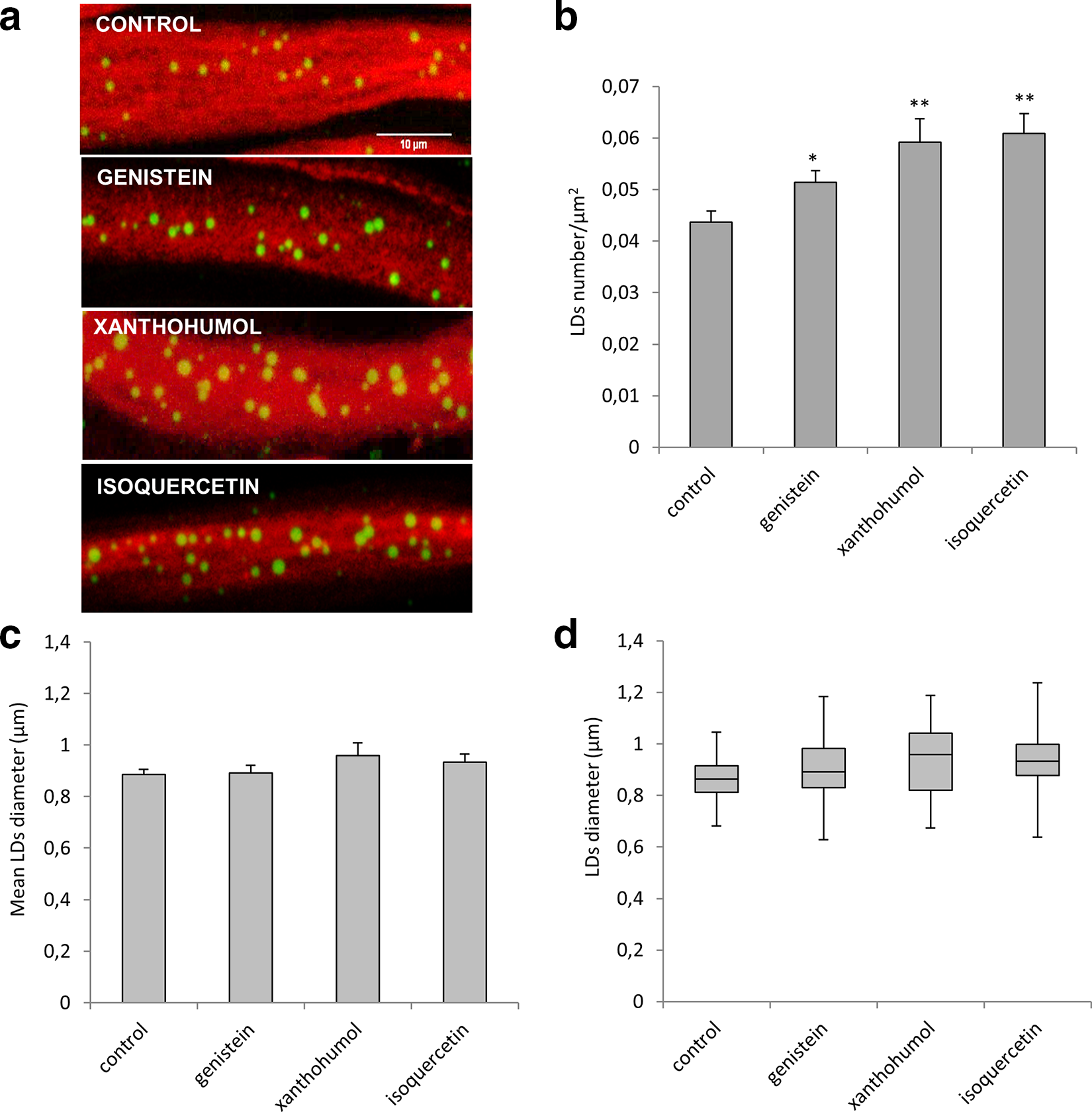
(a) Maximum-intensity projections of proximal ventral ganglion nerves from control or treated Drosophila third instar larvae. The compounds utilized were genistein (10 µM), xanthohumol (10 µM), and isoquercetin (10 µM). Nerves were labeled with anti-horseradish peroxidase antibody (red) to visualize neuronal membranes and BODIPY 493/503 dye (green) to determine the number and size of LDs. Scale bar = 10 µm. Both number (b) and size (c) of LDs were measured. (d) The results of the LDs size were plotted as box-and-whisker plot, showing the interquartile range, the median as horizontal bar, and the whiskers are the minimum and maximum values. Values are mean ± SD from at least 3 independent experiments. Significance was calculated using unpaired t-test (two-tailed) for LDs number and one-way analysis of variance with Tukey’s post hoc test for LDs size. P < 0.05* and P < 0.01** vs control. LD: lipid droplet.
Muscle fibers are full of LD in insect and mammals and are localized in close proximity to mitochondria in the subsarcolemmal region near the plasma membrane or in-between the myofibrils. The effects of flavonoids on LDs in the muscle tissues were analyzed (Figure 4a). Xanthohumol and isoquercetin led to a significant increase of LDs number: 0.114 LDs/μm2 and 0.165 LDs/μm2, respectively, compared to 0.091 LDs/μm2 of the control (Figure 4b). Intriguingly, the treatment with genistein was not able to modify the number or the mean size of muscle LDs, compared to control; however, it changed the LDs’ profile size, similar to the xanthohumol administration. In control larvae, we observed that LDs diameters range was between 0.4 and 1 µm, while in muscle larval tissues exposed to genistein and xanthohumol, the LDs diameter ranged from 0.6 to 0.8 µm (Figure 4d). Taken together, our results show that the flavonoids tested have also an undeniable effect on LDs biogenesis in muscles.

(a) Representative images of Drosophila larval muscles labeled with BODIPY 493/503. The compounds utilized were genistein (10 µM), xanthohumol (10 µM), and isoquercetin (10 µM). Scale bar = 20 µm. Quantification of LDs number (b) and diameter (c) of LDs. (d) The results of the LDs size were plotted as box-and-whisker plot, showing the interquartile range, the median as horizontal bar, and the whiskers are the minimum and maximum values. Values are mean ± SD from at least 3 independent experiments. Significance was calculated using unpaired t-test (two-tailed) for LDs number and one-way analysis of variance with Tukey’s post hoc test for LDs size. P < 0.05* and P < 0.01** vs control. LD: lipid droplet.
To assess the effect of different classes of flavonoids on LDs biogenesis, we evaluated the expression of the 2 key enzymes of Drosophila fatty acid metabolism, using a real-time reverse transcripatse polymerase chain reaction (RT-PCR) approach. The human glycerol-3-phosphate acyltransferase and diacylglycerol O-acyltransferase 1 (DGAT1) are encoded by Drosophila minotaur (mino) and midway (mdy) genes, respectively 30 (Figure 5a). We, thus, investigated whether the protective effects of flavonoids were linked to lipid droplets metabolism. Total RNA from larvae grown in standard and flavonoids-enriched food were extracted and subjected to quantitative real-time PCR procedure. As shown in Figure 5(b), isoquercetin and xanthohumol strongly enhanced both the transcripts expression of mino and mdy, whereas genistein administration had the minor effect.

(a) TAG de novo synthesis in mammals and Drosophila. (b) Relative mRNA levels of minotaur and midway after treatments. The compounds utilized were genistein (10 µM), xanthohumol (10 µM), and isoquercetin (10 µM). Values are mean ± SD from at least 3 different experiments. Significance was calculated using unpaired t-test (two-tailed). P < 0.05* vs control. GPAT: glycerol-3-phosphate acyltransferase; LPA: lysophosphatidic acid; AGPAT: 1-acylglycerol-3-phosphate acyltransferase; PA: phosphatidic acid; DAG: diacylglycerol; DGAT: diacylglycerol acyltransferase; TAG: triacylglycerol.
This is the first in vivo study conducted in Drosophila reporting the modulatory effects on LDs in different tissues due to phytochemical consumption. Our data show that different classes of flavonoids, at the concentrations here reported, can regulate lipid droplets biogenesis in Drosophila larvae and their effects are tissue specific. Almost all the flavonoids tested decreased LDs accumulation in fat bodies and increased LDs number and/or size in oenocytes and muscles, with the exceptions of genistein, which displayed a marginal effect on LDs in all the tissues analyzed. Furthermore, the effects of flavonoids on LDs seem to be correlated to the transcriptional levels of the metabolic enzymes mino (GAPT) and mdy (DGAT1), both involved in LDs biosynthesis. Accordingly, the transcriptional level of mino and mdy were barely upregulated after genistein administration. To our knowledge, the effects of flavonoids on LDs are not well studied and the research of LDs regulation and homeostasis has been principally focused on adipocyte cells. Flavonoids are considered antiobesity and lipid-lowering agents but they are also shown to display neuroprotective activity and prevent muscle atrophy. 31-34 Adipocyte specialized cells and their LDs have distinct properties compared to nonadipocyte cells. Increased LDs storage in adipocytes is considered a pathologic condition, while an increase of LDs in nervous system and muscles is considered a protective mechanism. 16,30,35-38
Our results confirm that flavonoids administrated to Drosophila, similar to mammalian systems, decreased fat storage in adipocyte-like tissues. LDs number, size, and total area occupied by LDs are decreased after flavonoids administration. This finding is in accordance with a previous study reporting that polyphenols as theaflavin, epitheaflagallin, and epigallocatechin suppressed LDs accumulation in larval fat body of a Drosophila model of obesity. The authors also reported that the 3 polyphenols prevented the changes of expression in lipid metabolism-related genes, induced by high-fat diet and thereby improved lipid metabolism. 39 Antidiabetic and dyslipidemia-correcting effects of polyphenols may be explained by different mechanisms that modulate the adipocyte number causing apoptosis and decreasing adipogenesis, or by modulating the lipid accumulation through activation of lipolysis and oxidation of fatty acids, or by decreasing lipid synthesis. 12,40,41 In our study, the increase of LDs in oenocytes suggests that fat body lipolysis is increased after flavonoids administration.
On the other hand, treatment with isoquercetin, genistein, and xanthohumol in Drosophila greatly modified LDs in nerves and muscles. Changes in the number of LDs have been associated to neurological disorders 42,43 and mutations of several LDs proteins are linked to motor neuron diseases. 44 Diets rich in polyphenolic compounds, like flavonoids, were associated with better cognitive functions and reduced risk of dementia and age-associated degenerative diseases. 8,45 In Drosophila, oxidative stress and hypoxia stimulate LDs biosynthesis in order to protect both neuroblasts and glia from peroxidation. LDs accumulation limits the reactive oxygen species (ROS) levels and inhibits the increase of polyunsaturated fatty acids. 46 Furthermore, studies performed in mice and Drosophila showed that LDs accumulation occurs in glial cells in a mitochondria dysfunction model as a protective mechanism. 47 This protection is attributed to the metabolic exchange between glia and neurons: glial lactate can supply neuronal lipogenesis in response to ROS production, and neuronal lipids are transported and stored in glia as lipid droplets. 10 Flavonoids can protect against cognitive deficits, neuronal injury, and oxidative stress induced in a rat model of Alzheimer’s disease, whereas genistein administration had the minor effect. 48 Similarly, isoquercetin and quercetin protect neurons from different cellular stresses. 33 It is also demonstrated that certain flavonoids protect muscle functionality, 34,49 and proved that LDs accumulation in muscles is a protective mechanism against the toxicity of lipid intermediates accumulation, such as lipid-derived DAGs and ceramides. 50
In this scenario, the isolation of certain lipids in specific cell compartments, such as LDs, is conceived as a lifeguard compensatory strategy. Considering that several studies demonstrate that some phytochemicals exert their beneficial effects through the activation of the adaptive stress response-signaling pathway, the effects of the flavonoids used could be due to the hormetic mechanism. The main pathways implicated in adaptive response include the activation of Nrf2/Keap1 and its interaction with other signaling such as Sirtuin-FOXO and NF-ĸB signaling. 51 More precisely, several studies suggest that phytochemicals act as a low-dose stressor activating adaptive response to prepare cell against stress condition. The accumulation of LDs could be a cell-type response to protect nonadipocytes from ER stress and lipotoxicity. Moreover, we found that LD changes in Drosophila tissues were associated with increased mdy and mino expression levels, suggesting the potential ability of certain flavonoids to promote LDs biogenesis. DGAT1 upregulation, which we observed after flavonoids treatment, has been demonstrated to protect cells from lipotoxicity and ER stress in both Drosophila and mammalian in vitro and in vivo models. DGAT1, in fact, functions in fatty acids re-esterification and formation of new small LDs after lipolysis to protect ER from lipotoxic stress and adipose tissue inflammation. 22 Moreover, DGAT1 protects mammalian cells during starvation-induced autophagy and increases LDs by activating the biogenesis to sequester FAs in TAG-rich LDs, preventing acylcarnitine accumulation and subsequent mitochondrial dysfunction. 21 In a Drosophila model of Huntington’s disease, the flies carrying a mutated Huntington protein (Htt) showed an alteration in the size and distribution of LDs. 43 In this light, the study of LDs biogenesis and turnover in cellular and animal models of neurological and muscle genetic disorders such as amyotrophic lateral sclerosis, myoclonic epilepsy, ataxia, and muscular dystrophy, 52-56 for which pharmacological treatments are still not available, could help to better clarify the biological role and importance of LDs in healthy and diseased states, as well as their pharmacological impact in dysfunctional conditions. In conclusion, the pharmacological modulation of lipid metabolism could be a potential novel molecular‐based approach for treating diseases due to dysfunctions of lipid storage. A major comprehension of phytochemicals mechanism of action and their biological targets can promote a more selective use of phytochemicals to repair pathological conditions. The knowledge of how different flavonoids act on lipid metabolism in different tissues can help to manage the use of phytochemicals with the aim of selectively ameliorating specific neuronal and metabolic diseases manifestations.
Experimental
Drosophila melanogaster Strain and Maintenance Conditions
A white mutant strain w1118 was obtained from the Blominghton stock center and used in all experiments. The flies were reared on a standard corn flour medium (4.4% w/v yeast granules, 4.4% w/v sucrose, 1.1% agar, and 2.6% w/v nipagin dissolved in ethanol 96%) at constant temperature of 25 ± 1°C and under a 12-hour dark/12-hour light cycle.
Antibodies, Chemicals, and Reagents
Antibodies and probe used for immunofluorescence analysis were anti-horseradish peroxidase antibody Cy3 or Cy5 conjugated (Jackson ImmunoResearch, Inc., West Grove, PA, USA), to bind Drosophila neuronal membranes, and BODIPY 493/503 dye (Thermo Fisher Scientific Inc., Waltham, MA, USA), to visualize and determine the LDs number and size. Trizol reagent was obtained from Thermo Fisher Scientific Inc (Waltham, MA, USA), and Direct-Zol TM RNA MiniPrep kit was purchased from Zymo Research (Irvine, CA, USA). Xanthohumol was purchased from HWI ANALYTIK GMBH (Rülzheim, Germania). All other chemicals used were purchased from Sigma-Aldrich (St Louis, MO, USA).
Flavonoids Treatments
Three different compounds (xanthohumol, isoquercetin, and genistein) were added to regular Drosophila food, all at the final physiological concentration of 10 µM as recommended in Kanazawa 57 . Flavonoids were dissolved in dimethyl sulfoxide. For control flies, the same amount of vehicle without chemicals was added to standard food. Briefly, adult flies from white1118 strain were allowed to lay eggs in flavonoids-enriched food, and in standard medium. Embryos were reared in these conditions until third instar wandering larval stage was concluded. The transcription levels of the main enzymes involved in LDs biogenesis and changes in LDs number and size in fat bodies, oenocytes, muscles, and nerves were then evaluated.
Immunohistochemistry and LDs Quantification
Third instar larvae were raised on regular and flavonoids-enriched food. After harvesting larvae, they were dissected in phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde in PBS for 10 minutes and washed in PBT 0.3% Triton X-00 (PBTX) as in Antonioli et al. 58 . To detect LDs in muscles, fat bodies, and oenocytes, the permeabilized larvae were incubated with BODIPY 493/503 (1:100 dilution in PBT 0.3%) for 1 hour at room temperature and mounted on glass slides with Mowiol 4-88. For LDs detection in proximal nerves the permeabilized larvae were stained with BODIPY 493/503 (1:100 dilution in PBT 0.3%) and with anti-horseradish peroxidase conjugated to Cy3 (1:500 dilution with PBT 0.3) for 1 hour at room temperature to mark neuronal membranes and mounted on glass slides with Mowiol 4-88. Images were acquired with a Nikon EZ-C1 confocal microscope (Tokyo, Japan) equipped with a Nikon Plan APO 60.0×/1.40 oil immersion objective as in Papadopoulos et al. 59 . Sixty Z-stacks with a step size of 1 µm were taken using identical settings. Each stack consisted of 10 to 12 plane images. Ten animals per treatment were analyzed. The area and number of LDs were calculated using Fiji (NIH) particle analyzer tool.
RNA Extraction and Real-Time Quantitative PCR
Total RNA was isolated from 5 third instar larvae per group and was extracted by Trizol reagent and purified using Direct-Zol TM RNA MiniPrep kit according to the manufacturer’s instructions. The RNA concentration was determined using a NanoDrop 2000c spectrophotometer (Thermo Fisher Scientific), and the purity of RNA was determined using the 260/280 nm absorbance ratio. A real-time quantitative PCR was performed on Eco Real-Time PCR System (Illumina Inc., San Diego, CA, USA), using One Step SYBR® Prime Script TM RT-PCR Kit II (Takara-Clontech, Kusatsu, Japan) as previously described. 60 The real-time PCR cycling conditions were reverse transcription 50°C for 15 minutes, polymerase activation 95°C for 2 minutes, followed by 40 cycles of 95°C for 15 seconds, 60°C for 1 minute; melting curve 95°C for 15 seconds, 55°C for 15 seconds, and 95°C for 15 seconds for all target genes. Primers used are listed below. Relative mRNA expression levels were adjusted with the housekeeping gene rp49 and were calculated by the threshold cycle (Cq) value of each PCR product and normalized using comparative 2- ΔΔCt method, which normalizes the expression of the target genes relative to a single reference gene and expressed relative to a reference sample. All experiments were performed in triplicate.
midway 5′-TGG TTA TGG CCT TGT TTC AA-3′
5′-CAC GTC CAT ATT GGA GAA CG-3′
minotaur 5′-CCC GAG AAG ATG GAA ACA TT-3′
5′-GGA GGA GGT GGA CTG ATT GT-3′
rp49 5′-AGG CCC AAG ATC GTG AAG AA-3′
5′-TCG ATA CCC TTG GGC TTG C-3′
Statistical Analysis
Statistics was performed with Graph Pad Prism 7.0 software. Significance was calculated using unpaired t-test (two-tailed) to assess the differences in the number and area of LDs, and one-way analysis of variance followed by post hoc Tukey’s honestly significant difference test to assess the differences in the size distribution of LDs. Differences were considered statistically significant at P < 0.05. For real-time PCR data were analyzed using unpaired t-test (two-tailed). Data are presented as mean ± SEM. At least 5 images of each sample were acquired and a minimum of 2 random regions of interest (ROI) of 35.5 µm2 were created from each image and the area of the LDs contained in the ROI was retrieved and analyzed.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by the Ricerca Corrente of Ministry of Italian Health; Fondazione Cariparo, Young Investigator Grant on Pediatric Research; PRID-J SID18-01 from Univeristy of Padova.