
Abstract
Background:
Porphyromonas gingivalis (P.g) is unique among pathogens due to its ability to generate citrullinated proteins in an inflammatory milieu, potentially mediating the loss of immune tolerance, the production of anticitrullinated protein antibodies (ACPAs), and subsequently the development of rheumatoid arthritis (RA). Based on this hypothesis, we set out to investigate whether P.g is linked to ACPAs in a well-characterized German population.
Participants and methods:
A total of 600 participants (292 women and 308 men with a mean age of 67 years) of the European Prospective Investigation into Cancer and Nutrition-Potsdam study were selected in 2013, and paired saliva and serum samples were collected. Salivary P.g DNA and serum anticyclic citrullinated peptide (anti-CCP2) levels were quantified by real-time polymerase chain reaction and anti-CCP2 enzyme-linked immunosorbent assay, respectively. In selected participants, additional ACPA fine-specificities were also analysed on a custom-made multiplex peptide array.
Results:
Among participants with C-reactive protein greater than 3.0 mg/l, a one-unit increase in P.g DNA was associated with an almost twofold increase in anti-CCP2 levels. Moreover, participants with high P.g DNA had on average approximately 2.8-times higher anti-CCP2 levels when compared with participants with low P.g DNA, (Holm-adjusted p value = 0.01). Furthermore, citrullinated epitopes on α-enolase and vimentin were common ACPA reactivities among participants who also had high P.g DNA and elevated C-reactive protein.
Conclusions:
Our study suggests that in specific subgroups of individuals with systemic inflammation, higher salivary P.g DNA is associated with elevated serum ACPA. These data support a role for P.g in the development of anticitrulline immunity.
Introduction
Chronic periodontitis is arguably the most prevalent inflammatory disease affecting between 20% and 50% of adults worldwide.1–4 Chronic periodontitis is the result of an aberrant host response to the oral microbial challenge in susceptible individuals. 5 Porphyromonas gingivalis (P.g) is considered a key periodontal pathogen 6 and P.g infection is highly prevalent in patients with chronic periodontitis. 7 Chronic periodontitis and P.g infection have been hypothesized to contribute to the development of rheumatoid arthritis (RA). 8 In fact, chronic periodontitis and P.g infection were shown to be associated with the prevalence of RA in a systematic review. 9
P.g has been suggested as an aetiological link due to its unique feature among prokaryotes to express an arginine-specific proteinase, referred to as peptidyl arginine deiminase enzyme (PPAD).10,11 The aetiological hypothesis is that P.g, through the actions of PPAD and gingipains, directly initiates protein citrullination (deamination of arginine residues) of both host and bacterial proteins, generating neoepitopes at mucosal surfaces.11,12 Continual spreading of neoepitopes and somatic hypermutation in genetically susceptible individuals eventually results in a breakdown of immune tolerance, upregulation of inflammatory responses, development of autoimmunity and the subsequent production of high-affinity antibodies to citrullinated proteins (ACPAs), a hallmark of RA.13,14
Local and systemic proinflammatory stimuli are essential for citrulline autoimmunity. 15 Indeed, citrullinated proteins and ACPAs are present in inflamed gingival tissue, suggesting local ACPA production, which may prime an individual for robust systemic ACPA production. 16 Observational epidemiological studies are commensurate with this hypothesis, in that higher ACPA titres were reported in the periodontium and serum of patients with periodontitis when compared with healthy controls.17,18 A small clinical study also suggests that ACPA titres in patients with periodontitis may be specifically associated with oral exposure to P.g. 19 Previously, we have shown elevated anti-P.g antibody levels in patients with RA who are positive for ACPAs, compared with controls, and that these antibodies precede the clinical onset of RA.20,21 This was confirmed in a meta-analysis. 22 Moreover, increased prevalence of periodontitis and P.g was shown recently in individuals at risk of being positive for ACPAs and unlike P.g, the abundance of another periodontal pathogen, Aggregatibacter actinomycetemcomitans was not increased in these individuals. 23 In addition, oral exposure of rodents to P.g was found to trigger seropositive arthritis. 24
Most of the evidence linking P.g to ACPA is from patient groups and clinical cohorts, and very few are based on the general population. Therefore, we set out to evaluate the association between the presence of P.g in saliva and serum ACPA levels in a well-characterized population-based cohort of apparently healthy individuals.
Participants and methods
Study design and population
Our study population was participants of the European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam study. Details of the study design, recruitment, and follow up of the EPIC-Potsdam participants have been described previously.25,26 In brief, the EPIC-Potsdam participants were a cohort of 27,548 individuals, aged 35–64 years, who were recruited between 1994 and 1998 from the city of Potsdam, Germany and neighbouring communities. During this period, participants underwent baseline examinations, blood sampling, interviews, and completion of questionnaires about lifestyle characteristics and medical history. Every 2–3 years, questionnaires were sent to participants in order to obtain additional lifestyle and health data. All medical conditions at baseline and during follow up were medically verified. The Ethics Committee of the State of Brandenburg, Germany, approved the study and all participants provided informed consent.
Between 2010 and 2012, 1472 active participants were selected using a rectangular sampling scheme to ensure an approximately uniform sample spread across sex and three age groups (35–44, 45–54, and 55–64 years). All participants were invited to attend the study centre for specific clinical assessments. Of these participants, 815 (55%) responded to the invitation and consented to participate. These 815 participants were re-invited in 2013, and 728 (89%) consented. Paired blood and saliva samples were provided by 704 (97%) out of the 728 participants.
Unstimulated saliva was collected at the study centre in the morning using the PSP® (Pre-analytical Sample Processing) SalivaGene System kit (STRATEC Molecular GmbH, Berlin, and Germany) and processed. The sample collection, post-collection handling, processing, and storage of the samples were standardized. However, no presampling instructions were given to the participants. 30 ml of blood was also collected and processed. Both samples were stored at −80°C until laboratory analysis. A schematic description of the EPIC-Potsdam study in phases leading to the current study population is shown in Figure 1.
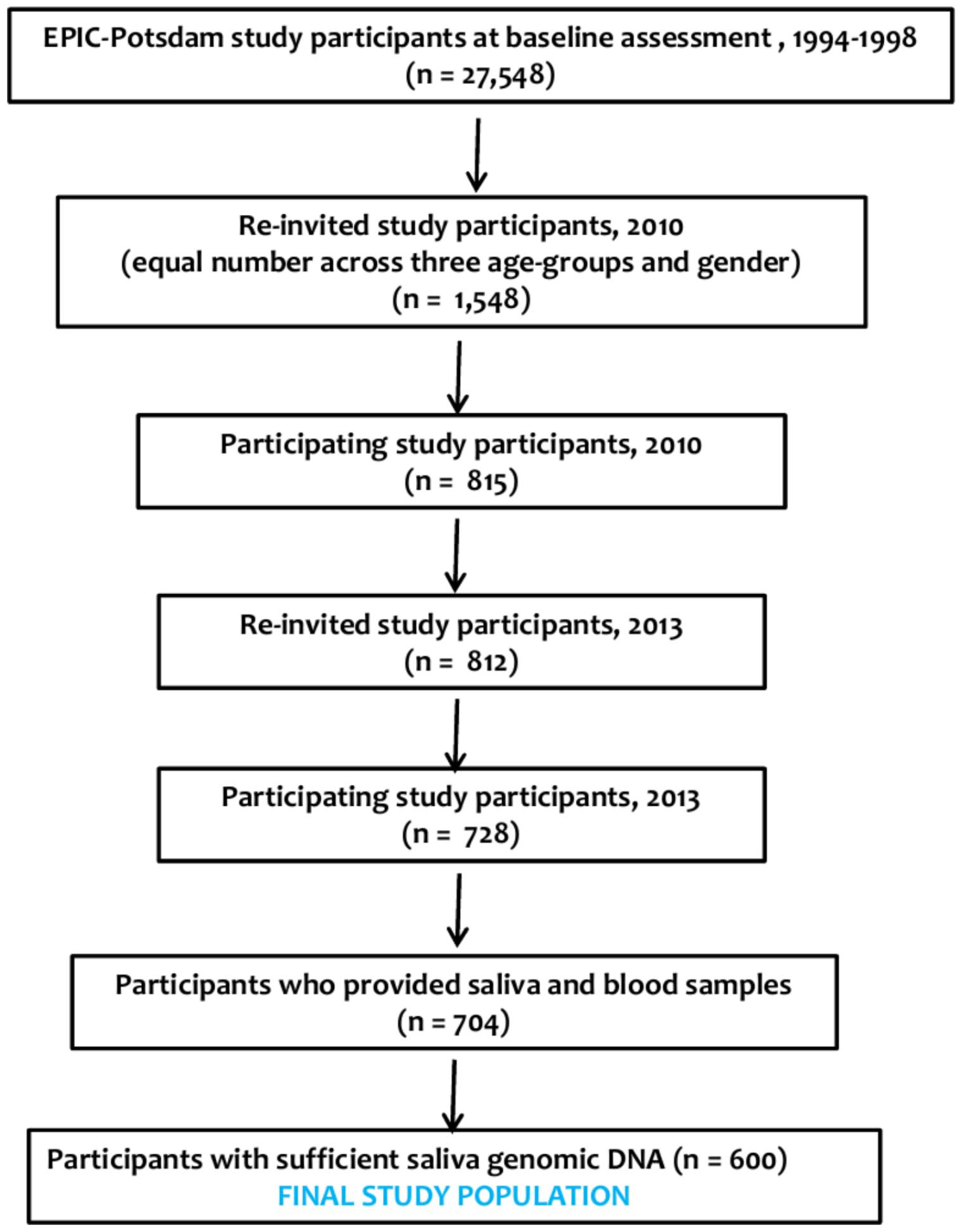
Flow chart of the current study population.
Isolation of genomic DNA from saliva
Genomic DNA (gDNA) was isolated from saliva using the QIAamp DNA mini kit (QIAGEN AB, Sollentuna, Sweden) according to the manufacturer’s protocol. In brief, a 300-µl aliquot of saliva was centrifuged at 10,000 r.p.m. for 15 min to pellet the sample. Then, 180 µl of buffer ATL and 20 µl of QIAGEN proteinase K were consecutively added to the pelleted sample. The mixture was vortexed and incubated at 56°C for 30 min, and purified using ethanol-containing buffers. The gDNA was eluted in nuclease-free water, followed by quantification and quality assessment (A260) with a NanoVue spectrophotometer (GE Healthcare Bio-Sciences, Uppsala, Sweden) and stored at −20°C until analysis.
Detection of P.g with quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qPCR) was performed on the gDNA using the universal primer 16S to detect the presence of nonspecific bacterial DNA and with P.g-specific primer to detect the presence of P.g. Duplicate samples were assayed in a total volume of 20 µl, containing 40 ng of the template gDNA solution, TaqMan Universal PCR Master Mix 2× (Applied Biosystems, Carlsbad, USA), and the specific set of primers (final concentration 18 µM) and probe (final concentration 5 µM; Cybergene AB, Sweden). P.g primers were: forward –GCG CTC AAC GTT CAG CC; reverse: CAC GAA TTC CGC CTG C; and probe: 6FAM CAC TGA ACT CAA GCC CGG CAG TTT CAA–TAMRA, and the 16s rDNA (universal) were forward: TGG AGC ATG TGG TTT AAT TCG A; reverse –TGC GGG ACT TAA CCC AAC A; and probe: 6FAM CAC GAG CTG ACG ACA RCC ATG CA–TAMRA. 27
The 7500 Fast Real-Time PCR System (Applied Biosystems) was used for amplification and detection of P.g. After an initial incubation step of 2 min at 50°C and denaturation for 10 min at 95°C, 40 PCR cycles (95°C for 15 s, 60°C for 1 min) were performed. The relative quantity of P.g was calculated as the fold difference in mean cycle threshold value of the P.g DNA relative to the mean cycle threshold value of the total bacterial DNA. DNA isolated from P.g strain ATCC 33277 was used as the positive control.
Detection and quantification of ACPAs
Serum samples were screened for the presence of ACPA immunoglobulin (Ig)G, using the Immunoscan CCPlus® enzyme-linked immunosorbent assay, from Euro-Diagnostica, Malmö, Sweden, according to the manufacturer’s instructions. In addition, sera from participants with anti-CCP2 antibody levels ⩾25 arbitrary units (AU)/ml (i.e. considered as anti-CCP2 antibody positive, according to the kit instructions) and an equal number of randomly selected participants with anti-CCP2 antibody levels <25 AU/ml (i.e. anti-CCP2 negative) were further analysed for the presence of ACPA fine-specificities using a custom-made microarray based on the ImmunoCAP ISAC microarray system (ThermoFisher Scientific, Phadia AB, Uppsala, Sweden), as previously described. 28 We investigated 10 citrullinated peptides derived from four different human proteins (fibrinogen, vimentin, filaggrin, and alpha-enolase) and 1 additional citrullinated peptide derived from PPAD, their arginine-containing control peptides, and 17 other autoantibody reactivities. The cut-off values for positivity (for each ACPA fine-specificity) were based on the 98th percentile cut-off among healthy controls. Supplemental Tables 1 and 2 provide details of the citrullinated and noncitrullinated antigens respectively.
Measurement of serum markers of systemic inflammation
Circulating levels of two markers of inflammation, C-reactive protein (CRP) and white blood cell (WBC; leukocyte) count, was measured. CRP was measured using a high-sensitivity immunoturbidimetric assay and expressed in mg/l. WBCs were measured by fluorescence flow cytometric method and expressed in Gpt/l.
Assessments of covariates
Information on sociodemographic characteristics, anthropometry, lifestyle, current medications (within 7 days), including antibiotics, nonsteroidal anti-inflammatory drugs (NSAIDs), and synthetic or biological disease-modifying antirheumatic drugs (DMARDs) were also reported. These covariates were ascertained with the use of questionnaire at baseline or during follow up. Importantly, participants were asked if they had ever been diagnosed with periodontitis and rheumatic diseases. These data are the status of the participants as of 2013. Details of covariates are available in supplemental text.
Statistical analyses
From the initial 704 individuals, we excluded 104 because of low gDNA levels that were insufficient for P.g quantification. There was no significant difference between these individuals and the 600 participants with sufficient gDNA (Supplemental Table 3). Using tertiles, 600 participants were categorized into three groups of low, moderate, and high P.g according to the relative quantity of P.g. For normally distributed continuous variables, non-normally distributed continuous variables, and categorical variables, one-way analysis of variance, Kruskal–Wallis nonparametric test, and Chi-square tests were used as statistical tests for differences across P.g groups, respectively.
We determined the relation between P.g and anti-CCP2 IgG by fitting Tweedie generalized linear models (details in supplemental text). The Tweedie generalized linear model is appropriate for modelling semicontinuous dependent variables 29 (anti-CCP2 IgG in the current study). Our independent variable, P.g fold difference was modelled in two ways: continuously and categorized into tertiles. At first, we investigated whether the association between P.g and anti-CCP2 varied across subgroups of age, sex, body mass index, smoking status, periodontitis status, rheumatic diseases, CRP, and WBC count (details of subgroups in supplemental text) by including a product term between P.g and each variable in univariate analysis. Afterwards, we controlled for confounding by adjusting for covariates derived using directed acyclic graphs. Overall, we had four models: model 1 with P.g only, and three multivariable models (models 2–4; adjustment sets in supplemental text). The beta estimates were exponentiated as relative risk (RR). The RR of the continuously modelled P.g was interpreted unit change in anti-CCP2 associated with one-unit change in the fold difference of P.g. The categorically modelled P.g was interpreted as the ratio of the mean of anti-CCP2 between participants with either high or moderate P.g compared with the reference category, low P.g. These P.g group comparisons were adjusted for multiple comparisons using the Holm method. We also assessed the robustness of these analyses by excluding participants with myocardial infarction, stroke, and antibiotics, NSAID and DMARD use. Additionally, we investigated the nonlinear association between P.g and anti-CCP2 using restricted cubic spline regression adjusted for covariates in model 4. Anti-CCP2 was transformed using the Box–Cox method (details in supplemental text).
Finally, for each individual within the subsets of participants that were positive for anti-CCP2 and equal numbers of randomly selected participants who were negative for anti-CCP2, we reported the presence (and number) or absence of additional antibody reactivities, the number of ACPA/non-ACPA antibody reactivities, the quantity of P.g, CRP level, rheumatic disease, and the use of DMARDs or NSAIDs.
All statistical analyses were performed using SAS Enterprise Guide, version 6.1 (SAS Institute Inc., Cary, NC, USA). For all analysis, p ⩽ 0.05 was considered statistically significant.
Results
Characteristics of the study population
The age range of the final study population (n = 600, comprising 292 women and 308 men) was 47–82 years (mean age is 67 years with a standard deviation of 8.32). As shown in Table 1, across groups of increasing P.g, there was increasing sport time, higher alcohol consumption, a higher prevalence of type 2 diabetes mellitus (T2DM), and a higher leucocyte count. Conversely, there was a lower proportion of higher-grade professionals. No difference was observed regarding other characteristics. Overall, 11 participants, corresponding to 2% of the study population, were positive for the anti-CCP2 antibody. The majority of the individuals who were positive for the anti-CCP2 antibody (n = 6) were found in the subset with a high P.g (Table 1).
Characteristics of the study population (N = 600).

Anti-CCP2, second generation anticyclic citrullinated peptides antibody; AU, arbitrary units; DMARD, disease-modifying antirheumatic drug; ever-smokers, former smokers and current smokers; IgG, immunoglobulin G; M, mean; NS, nonsignificant at p value < 0.05; NSAID, nonsteroidal anti-inflammatory drug; P.g, Porphyromonas gingivalis; SD, standard deviation.
Number of cigarettes of current smokers.
Anti-CCP2 IgG ⩾25 AU/ml.
Detection of P.g DNA was performed using real-time PCR analysis. The relative quantity of P.g was calculated as the fold difference in mean cycle threshold value of the P.g DNA relative to the mean cycle threshold value of the total bacterial DNA. P.g quantity (fold difference) was categorized into low, moderate and high, according to tertiles.
Multivariable analyses of the association between quantity of P.g and anti-CCP2 IgG levels
The interaction term of P.g (both as continuous and categorical variable) and CRP group on the continuously modelled dependent variable, anti-CCP2 IgG, was significant (p for interaction = <0.01). This significant interaction term indicates that the association between P.g and anti-CCP2 IgG depends on CRP group. None of the interaction terms of P.g with other covariates were significant. Therefore, we fitted regression models separately for each CRP category and effect estimates, and RRs were presented as such. For each CRP category CCP2 model, the goodness of fit, Pearson’s Chi-square statistic divided by the degrees of freedom was close to one (1.25 for CRP >3.0 mg/l, 1.62 for CRP 1.0–3.0 mg/l, and 1.86 for CRP <1.0 mg/l) showing that all specified CCP2 models fitted the data well.
Among participants with CRP >3.0 mg/l, in the fully adjusted model, a one-unit increase in the fold difference of P.g was associated with an almost twofold increase in anti-CCP2 IgG levels. Moreover, when compared with participants with low P.g, participants with high P.g had on average approximately 2.8 times higher anti-CCP2 IgG levels [RR = 2.82; 95% confidence interval (CI): 1.60–9.11, Holm-adjusted p value = 0.01; Table 2]. Other covariates significantly associated with levels of anti-CCP2 IgG were smoking and education. Specifically, there was a threefold risk of elevated anti-CCP2 IgG levels among ever-smokers when compared with nonsmokers (RR = 3.24; 95% CI: 1.72–4.26, p = 0.01), and a threefold risk of elevated anti-CCP2 IgG levels among those with technical school educational attainment when compared with those with a university degree (RR = 2.61; 95% CI: 1.23–3.72, p = 0.02).
Multivariable analyses of the association between quantity of P.g DNA and anti-CCP2 antibody levels.
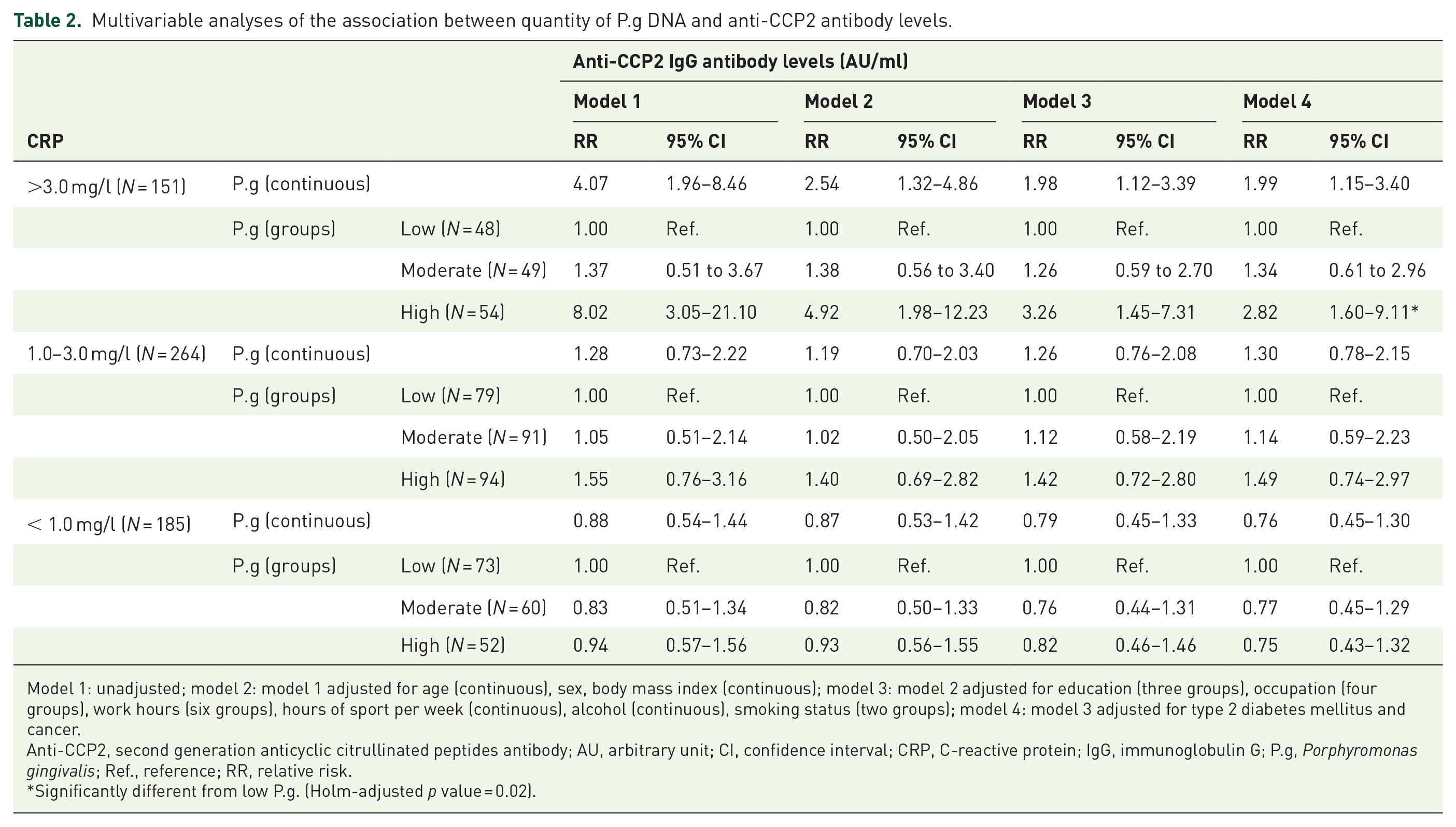
Model 1: unadjusted; model 2: model 1 adjusted for age (continuous), sex, body mass index (continuous); model 3: model 2 adjusted for education (three groups), occupation (four groups), work hours (six groups), hours of sport per week (continuous), alcohol (continuous), smoking status (two groups); model 4: model 3 adjusted for type 2 diabetes mellitus and cancer.
Anti-CCP2, second generation anticyclic citrullinated peptides antibody; AU, arbitrary unit; CI, confidence interval; CRP, C-reactive protein; IgG, immunoglobulin G; P.g, Porphyromonas gingivalis; Ref., reference; RR, relative risk.
Significantly different from low P.g. (Holm-adjusted p value = 0.02).
On the contrary, in groups of CRP 1.0–3.0 mg/l and CRP < 1.0 mg/l, there was no association between continuously modelled P.g and anti-CCP2 antibody levels. Similarly, there was no difference between the moderate and high P.g when compared with low P.g (Table 2). In addition, no covariates were significantly associated with levels of anti-CCP2 IgG in both groups. Further, there was no evidence of nonlinear associations between P.g and anti-CCP2 in both groups (Figures 2 and 3).

Restricted cubic spline regression graph to model nonlinear association among participants with CRP between 1.0 and 3.0 mg/l.
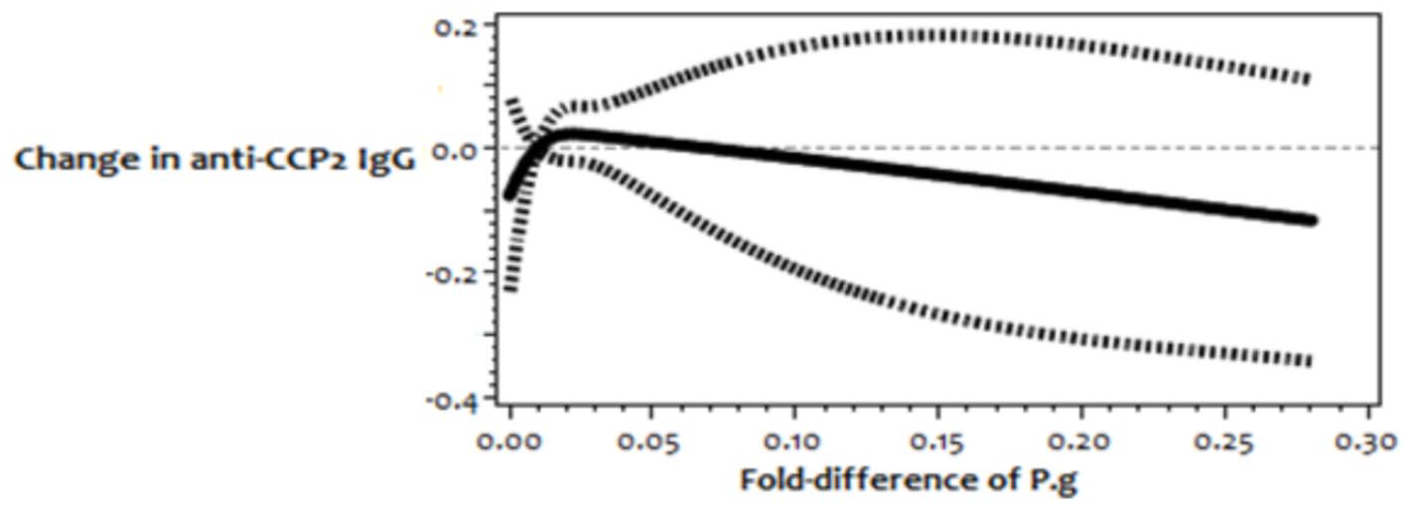
Restricted cubic spline regression graph to model nonlinear association among participants with CRP < 1.0 mg/l.
In sensitivity analyses, excluding participants with myocardial infarction, stroke and those who reported intake of antibiotics did not affect the results (data not shown). Moreover, excluding participants with CRP ⩾ 10.0 mg/l (as a marker of acute inflammation) did not affect the results either. Interestingly, when we excluded all 11 individuals who were positive for anti-CCP2, as well as those using NSAIDs and DMARDs; there still were higher anti-CCP2 IgG levels among participants with high P.g when compared with those with low P.g, within the group with CRP >3.0 mg/l.
Comparison of individuals positive and negative for anti-CCP2 with respect to additional autoantibody reactivities
Overall, 5 out of the 10 individuals who were positive for anti-CCP2 (1 was excluded due to technical reasons) had at least one additional autoantibody reactivity, while 4 individuals out of the 11 randomly selected individuals who were negative for anti-CCP2 had at least one additional autoantibody reactivity (Table 3). Antibody reactivities corresponding to different ACPA fine-specificities were only detected in individuals who were positive for anti-CCP2. Antibodies against citrullinated α-enolase (anti-CEP-15–21 IgG) and citrullinated vimentin (anti-Cit-Vim60–75 IgG) constituted the most commonly detected ACPAs, and were present in four out of five individuals who were double positive for anti-CCP2/ACPA fine-specificity. Moreover, these four individuals also had a high quantity of P.g and high CRP levels (>3.0 mg/l). In addition, three out of the five individuals who were double positive for anti-CCP2/ACPA fine-specificity used DMARDs or NSAIDs, and three reported a rheumatic disease. One of these individuals was also positive for non-ACPA autoantibodies, directed against double-stranded DNA and U1 RNP-C complex protein. The individuals who were positive for anti-CCP2 and negative for the ACPA fine-specificities (CCP2+ACPA−) had relatively lower anti-CCP2 IgG levels compared with individuals who were positive for both anti-CCP2 and ACPA fine-specificities (CCP2+ACPA+). Moreover, individuals who were positive for anti-CCP2 ‘P.g high’ when compared with individuals who were positive for anti-CCP2 ‘P.g low’ generally had rheumatic disease, high CRP, and used DMARDs or NSAIDs (Table 3).
Comparison of individuals positive for anti-CCP2 and randomly selected individuals negative for anti-CCP2.

AU, arbitrary unit; CCP, cyclic citrullinated peptide; CEP, citrullinated α-enolase peptide; Cit-Fib, citrullinated fibrinogen; Cit-Fil, citrullinated filaggrin; Cit-Vim, citrullinated vimentin; CRP, C-reactive protein; DMARD, disease-modifying antirheumatic drug; dsDNA, double-stranded DNA; NSAID, nonsteroidal anti-inflammatory drug; P.g, Porphyromonas gingivalis; Ro60/SSA, 60 kD RNA-associated protein; SmBB, small nuclear ribonucleoprotein-associated protein BB ribonucleoprotein; U1 RNP-C, human recombinant RNP-C protein.
No ACPAs were detected among 11 randomly selected individuals who were negative for anti-CCP2. Antibody reactivities in this subset included double-stranded DNA, 60 kD RNA-associated protein, and small nuclear ribonucleoprotein-associated protein BB. In addition, none of these individuals reported a rheumatic disease or DMARD use (Table 3).
Discussion
In this study of 600 individuals from a general population-based study cohort, we found a positive association between increasing quantity of salivary P.g DNA and serum anti-CCP2 IgG levels, after adjusting for various covariates. Specifically, this association was confined to individuals with systemic inflammation, CRP >3 mg/l. We also found that those individuals who were positive for a number of ACPA fine-specificities also demonstrated a higher quantity of salivary P.g and higher CRP levels when compared with individuals who were negative for ACPAs.
Our data demonstrating an association between P.g DNA and ACPAs support the aetiological hypothesis first proposed by Rosenstein and colleagues 13 that P.g infection plays a causative role in the development of ACPAs. This hypothesis was also supported by a number of other reports pinpointing a link between P.g infection and ACPA-positive RA. Some of these studies reported an association between the presence and abundance of P.g and anti-CCP2 antibodies, 18 while other studies,30–32 including our recent publications20,21 and a recent meta-analysis 22 reported a link between anti-P.g antibody levels and ACPAs, and yet others reported an association between clinical periodontitis and ACPAs.33–35 Mikuls and colleagues 30 additionally reported an association between anti-P.g antibody levels and CRP, suggesting that P.g infection influence both innate and adaptive immune responses. However, in other studies no relationship between anti-CCP2 and P.g abundance,36,37 anti-P.g antibody levels,38,39 and clinical periodontitis alone 40 was reported.
In relation to this ambiguity, elevated anti-CCP2 antibody levels were observed in patients with periodontitis and dental plaque P.g, as compared with patients with periodontitis without dental plaque P.g and healthy controls, 18 suggesting that the actual presence of P.g, rather than the severity of the periodontal lesion, is crucial in the relationship with ACPAs and ACPA-positive RA. This notion was also brought forward in our recent studies, where we were unable to identify a link between periodontitis and ACPA-positive RA, 41 but in the same RA cohort, we could demonstrate elevated anti-P.g antibody levels in ACPA-positive RA compared with both controls and ACPA-negative RA. 20 Based on data from these studies, we concluded that the previously reported link between periodontitis and RA could be accounted for by P.g infection. Moreover, it has been suggested that P.g infection contributes to the overall increase in ACPA seropositivity in individuals without RA.24,42,43
Importantly, the association between the presence of P.g DNA and higher serum anti-CCP2 antibody levels among individuals with high CRP in our study is noteworthy, given the evidence that P.g contributes to a systemic inflammatory milieu, 44 and that citrullination of proteins is known to occur in response to inflammation; 15 this may foster the generation of ACPAs in genetically susceptible individuals. 14 Our findings, with respect to the interplay between P.g and CRP on anti-CCP2 antibody, supports a recent review reporting the role of P.g and the proinflammatory profile among other factors in the association between periodontitis and RA. 45
This provides insight into the mechanisms for the development of ACPAs. The low ACPA levels in one individual with high a quantity of salivary P.g DNA and high CRP levels might suggest a successful clearing of citrullinated proteins, and potentially the lack of genetic risk alleles (i.e. the HLA-DRB1 shared epitope), known to associate strongly with ACPA production.
Intriguingly, CEP-1 was one of the two most common ACPA fine-specificities among participants with high P.g DNA. This supports previous findings that PPAD can citrullinate human α-enolase, 12 and our previous report that anti-CEP-1 antibodies purified from RA patients bind citrullinated P.g enolase, forming the basis of a molecular mimicry hypothesis. 11 This hypothesis links P.g with RA via citrullination of enolase and ACPA production. Our CEP-1 data also support recent findings from another German population, that periodontitis patients with P.g infection have elevated anti-CEP-1 antibody levels. 46 Moreover, patients with RA with moderate to severe periodontitis and the presence of P.g have elevated anti-CEP-1 antibody levels compared with patients with RA without periodontitis or P.g. 40
The proportion of individuals who were ACPA-positive in our study (1.8%) is similar to that reported from a large Swedish population-representative cohort (2.8%). 47 With a specificity and sensitivity of the anti-CCP2 assay of approximately 97% and 70%, respectively, and a Germany RA prevalence of 1.38%, 48 about 25% (4) of our study participants who were positive for anti-CCP2 antibody are expected to have RA, perhaps all four individuals who reported rheumatic disease. Specifically, the individuals who were double positive for anti-CCP2 and ACPA fine-specificity were likely suffering from RA, since a combination of ACPAs, use of DMARDs or NSAIDs, and high CRP are indicative of an RA diagnosis, 49 or ‘pre-RA’, since elevated anti-CCP2 are known to precede the development of RA. 50
Furthermore, smoking is the main environmental risk factor for ACPA-positive RA, 51 and the association between smoking and elevated anti-CCP2 antibody levels as seen in our study is also a well-known phenomenon in ‘pre-RA’. 52 Studies that reported a relation between lower educational level and anti-CCP2 antibodies is scarce. However, lower socioeconomic status because of lower educational attainment has been postulated to influence the risk of RA. 53 Consistent with our findings on factors related to the quantity of P.g, others have documented that individuals who have a higher alcohol intake harbour more P.g, 54 individuals with T2DM have a high quantity of salivary P.g, 55 and P.g infection induces local and systemic accumulation of leucocytes. 56 Aside from poor oral health that is generally observed in athletes 57 and the relation between educational level and P.g, 58 the association of P.g with sport activity and occupation has not been previously reported. Possible explanation for these determinants of P.g are alcohol consumption resulting in dysbiosis, 54 dehydration and local drying of the mouth during sporting activity reducing antimicrobial activity in the mouth 57 and systemic diseases weakening resistance to infection and altering inflammatory response. 59
A previous study 60 did not observe difference in individuals with P.g with RA compared with controls. One possible explanation for the contrasting results of Zhang and colleagues 60 is the discrepant detection of P.g that is related to age.58,61 Our study population (average age of 67 years) is older than that of Zhang and colleagues (the highest age was 65 years and the average age was not reported) 60 and since saliva P.g increases with age;58,61 it is likely that the more detectable P.g in our study population partly results in association with anti-CCP2/RA.
The strengths of our study were that participants were selected with special consideration for sex and age strata and were thus representative of a well-characterized population of apparently healthy individuals. In addition, both exposure and outcome were measured objectively, that is, in biological samples. Moreover, compared with P.g detection from dental plaque or another oral niche, salivary P.g captures an aggregate of overall P.g exposure, because P.g and other oral bacteria are continually shed into the saliva from all ecological sites. This study adjusted for several factors such as age, sex, smoking, health conditions, and medications, which might have confounded our results.
Nevertheless, some points have to be taken into account when interpreting our findings. The participants’ physicians verified most chronic diseases in the EPIC-Potsdam study, but periodontitis and rheumatic diseases were not verified because the EPIC-Potsdam study was not primarily designed to investigate dental health and rheumatic diseases. Hence, detailed information, such as dental hygiene practices, periodontal care, specific types of rheumatic disease and their duration were not available. Nevertheless, most of the covariates and confounders that were considered in the analysis were verified. In addition, the absence of an association between the quantity of P.g and periodontitis could be due the fact that periodontitis was self-reported and the poor validity of self-reported periodontitis, which is well documented, 62 and could also be explained in part by the fact that periodontitis is a polymicrobial infection and that P.g might be absent in some individuals with periodontitis. When sampling saliva, participants are often instructed not to eat, drink, smoke, or chew gum or perform oral hygiene for 30 min prior to sampling. These instructions were not given to our participants and these factors might have affected yields of gDNA and hence the quantity of P.g. However, the fact that gDNA was not significantly associated with quantity of P.g in the current study, and considering that these factors are unlikely to affect CCP2 IgG levels suggest that they cannot confound the association between P.g and ACPA. Nonetheless, future studies should endeavour to standardize saliva presampling, sampling, and postsampling procedures. Another drawback is that we did not specifically ask participants for RA diagnosis, but ‘rheumatic diseases’, which comprise an array of diseases. Rheumatoid factor, another important RA biomarker was not measured in this study. Additional reactivity to fine-specific ACPAs could have been determined in all (or larger number of) samples, but this was not feasible due to logistic reasons.
A further limitation of our study was its cross-sectional design. One would expect time lags between the colonization of P.g, onset of P.g infection, and the development of antibodies to citrullinated proteins, 31 therefore elevated ACPAs may reflect the action of past P.g infection only or in consort with present infections. Surely, serum IgG to virulence factors in P.g such as arginine-specific gingipain B or outer membrane antigen may be a more suitable marker of both present and past P.g infection. The homogenous German study population may limit generalizability of this current study to other nationalities or ethnic groups, although our findings are similar to reports in other populations. 30 Furthermore, we cannot rule out residual and unmeasured confounders of the association between P.g and ACPAs, such as levels of total IgG. In addition, we did not control for confounding between CRP and ACPAs since our primary interest was to assess the relation of P.g DNA with ACPA levels. Moreover, reverse causation due to RA was unlikely in the relation of P.g with CCP2 IgG levels since we excluded participants using DMARDs as proxy for RA in sensitivity analysis. The focus of this study was to investigate the association between salivary P.g and serum ACPA levels; hence, we did not perform measurements of anti-P.g antibodies in our participants. The measurement of anti-P.g antibodies levels and investigation of their relation to P.g abundance is planned for future studies.
Within the limits of our study, salivary P.g DNA was linked to serum anti-CCP2 in the presence of systemic inflammation, and individuals who were anti-CCP2 positive showed additional reactivity to fine-specific ACPAs. Individuals with double positive anti-CCP2 and ACPA fine-specificity also had high P.g and were likely suffering from RA. Longitudinal multicentre studies with larger study populations to investigate the relationship between P.g colonization and infection and ACPAs are warranted.
Supplemental Material
Supplementary_file_TAB-19-07-034.R1 – Supplemental material for A cross-sectional investigation into the association between Porphyromonas gingivalis and autoantibodies to citrullinated proteins in a German population
Supplemental material, Supplementary_file_TAB-19-07-034.R1 for A cross-sectional investigation into the association between Porphyromonas gingivalis and autoantibodies to citrullinated proteins in a German population by Kolade Oluwagbemigun, Tülay Yucel-Lindberg, Thomas Dietrich, Gregory Tour, Natalia Sherina, Monika Hansson, Manuela Bergmann, Karin Lundberg and Heiner Boeing in Therapeutic Advances in Musculoskeletal Disease
Footnotes
Acknowledgements
We thank the staff of the Human Study Centre of the German Institute of Human Nutrition Potsdam-Rehbrücke, Germany, namely of the trustee and the examination unit for the data collection, of the data hub for the processing, and of the Biobank for the processing of the blood samples. We thank the participants of the EPIC-Potsdam study for the provision of their data and biological samples. We also acknowledge the valuable help of Jana Foerster and the text-editing support of Friederike Querengässer.
Kolade Oluwagbemigun (KO), Tülay Yucel-Lindberg (TY), Karin Lundberg (KL), Manuela Bergmann (MB), and HB (Heiner Boeing) designed the current study. KO, TY, and Gregory Tour isolated genomic DNA from saliva and detection of P.g. KO, Natalia Sherina (NS), and KL screened participants for anti-CCP2. Monika Hansson and KL screened participants for other ACPA fine-specificities. KO, TY, and KL analysed and interpreted the data. KO was the main contributor in writing the manuscript. TY, KL, NS, Thomas Dietrich, MB and HB revised the manuscript. All authors read and approved the final manuscript. KL and HB share senior authorship.
The Ethics Committee of the State of Brandenburg, Germany, approved the study and all participants provided informed consent. Consent forms of the participants are available at the Human Study Centre of the German Institute of Human Nutrition, Potsdam-Nuthetal, Germany. The datasets analysed during the current study are not publicly available, but are available from the corresponding author on reasonable request
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The recruitment part of the EPIC-Potsdam study, Germany was supported by the German Federal Ministry of Science, Bonn, Germany (01 EA 9401) and the European Union (EU), Brussels, Belgium (SOC 95201408 05F02), while the follow up of the EPIC-Potsdam Study was supported by the German Cancer Aid, Bonn, Germany (70–2488–Ha I) and the European Community (now EU), Brussels, Belgium (SOC 98200769 05F02). The work concerning P.g and autoantibodies to citrullinated proteins analyses was supported by grants from the EU-funded ITN project RAPID (290246), the EU-funded FP7 project TRIGGER (FP7-Health-2013-306029), Swedish Research Council, and Stockholm County Council. KO is currently supported by the Federal Ministry of Education and Research, Germany (grant number: 01EA1705A) through the European Joint Programming Initiative ‘A Healthy Diet for a Healthy Life’ project, Metabolic HEALTH through nutrition, microbiota and tryptophan bioMARKers (HEALTHMARK). The funders had no influence on the current study, as regards its design, analysis of data, decision to publish, and preparation of the manuscript.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.