
Abstract
Tissue inhibitor of metalloproteinase-3 (TIMP-3), a secreted glycoprotein, plays an important role in carcinogenesis. It can bind to many proteinases to suppress their activity and thus protect the extracellular matrix from degradation. TIMP-3 may have many anticancer properties, including apoptosis induction and antiproliferative, antiangiogenic, and antimetastatic activities. This review summarizes the structure, proteinase inhibition ability, genetic and epigenetic regulation, cancer therapy potential, and contribution to cancer development of TIMP-3. Furthermore, in this review we discuss its potential as a biomarker for predicting cancer progression and the current state of drugs that target TIMP-3, either alone or in combination with clinical treatment. In conclusion, TIMP-3 can be a biomarker of cancer and a potential target for cancer therapy. This review article can serve as a basis to understand how to modulate TIMP-3 levels as a drug target of cancers.
Keywords
Introduction
Despite recent improvements, cancer treatment remains associated with several challenges. Cancer development is a multifactorial and multistep process and involves several genetic and epigenetic regulations. In addition, cancer cells possess several unique characteristics, also known as the cancer hallmark, that confer the cells with resistance against the human immune system and cancer treatment. The cancer hallmark includes tumor-promoting inflammation, enabling replicative immortality, avoiding immune destruction, evading growth suppressors, sustaining proliferative signaling, deregulating cellular energetics, resisting cell death, genome instability and mutation, inducing angiogenesis, and activating invasion and metastasis.1,2 Metastasis, which is the major cause of death among cancer patients, involves multiple processes including extracellular matrix (ECM) remodeling and degradation. Degradation of the ECM is required for tumor cell metastasis; this is achieved by several proteinases such as the plasmin system and particularly, the matrix metalloproteinases (MMPs).3–6 MMPs are known to play an important role in the tissue invasion and metastasis of cancer cells. The tissue inhibitors of MMPs (TIMPs) are endogenous inhibitors of MMPs, and regulation of MMPs by TIMPs is particularly important for the maintenance of the ECM. Disruption of the balance between the activities of MMPs and TIMPs during carcinogenesis may affect invasion and metastasis7–9 and may worsen patient outcomes. 10 TIMP-3, a member of the TIMP family, is a 24-kDa secreted glycoprotein, and its gene is located on chromosome 22q12.1–q13.2. Knockout of the TIMP-3 gene in mice resulted in increased MMP, a disintegrin and MMPs with thrombospondin motifs (ADAMTS) activity, and cartilage degradation, suggesting that reduced TIMP-3 levels may cause osteoarthritis. 11 In addition, the absence of TIMP-3 leads to poor cardiac remodeling and has been associated with myocardial infarction or hypertension.12,13 In cancer studies, TIMP-3 plays an important role in the cancer hallmark by controlling cell death, angiogenesis, tumor inflammation, and tumor cell invasion and dissemination. 14 For instance, TIMP-3 restoration in cancer cells inhibits cell growth and promotes cell apoptosis.15,16 In addition, TIMP-3 overexpression improves the sensitivity of osteosarcoma to clinical drug treatment through interleukin (IL)-6 inhibition. 17 TIMP-3 also acts as a potential antiangiogenesis agent by inhibiting endothelial cell tube formation. 18 Moreover, TIMP-3 can inhibit cancer cell migration, invasion, and metastasis in vitro and in vivo.19,20 Clinical studies have reported reduced TIMP-3 expression in cases of several cancer types compared with normal controls;19–22 the loss of TIMP-3 may lead to poor outcomes, including large tumor size, high tumor stage, and metastasis.23–25 Herein, we review the structure and function of TIMP-3 and discuss its contribution to carcinogenesis and its potential in cancer therapy.
TIMP-3
TIMP classification
The TIMP family contains four members: TIMP-1, TIMP-2, TIMP-3, and TIMP-4. The molecular weight of the TIMPs is approximately 21 kDa, and they contain an N-terminal domain and a C-terminal domain. In contrast to that in TIMP-1 and TIMP-3, the C-terminal domain in TIMP-2 and TIMP-4 is negatively charged. Compared with the nonglycosylation of TIMP-2 and TIMP-4, TIMP-1 contains two N-glycosylation sites at Asn 26 and Asn, 27 and TIMP-3 contains a single N-glycosylation site in the C-terminal domain. 28 Glycosylated forms of TIMP-3 have higher affinity for glycan-bound MMPs and are protected from endocytosis and degradation. 29 Unlike other TIMPs, TIMP-3 is the only TIMP that binds firmly to the ECM after its secretion. 30 This binding is via the interaction of the N-terminal domain with heparan sulfate and sulfated glycosaminoglycans. 31
Transcriptional regulation of TIMP-3
The expression of TIMP-3 can be regulated by transcriptional regulation. Transcriptional regulation contains two major parts: the first part involves transcription factors and the transcription apparatus and the second part involves chromatin and its regulators. 26 Gene expression regulated by transcription factors is one of the most common transcriptional regulations. Transcription factors including Elf3, sp1, smad2, and smad4 have been reported to target on the promoter of TIMP-3 and regulated TIMP-3 expression.32–36 Jobling et al. discovered that ETS transcription factor Elf-3 was expressed in human retinal pigment epithelium (RPE) cell lines. Transfection of Elf3a and Elf3b overexpression vector increased promoter activity of TIMP-3. 32 TIMP-3 promoter contains four sp1 binding sites in the region near the transcription start site. 35 Zerrouqi et al. indicated that P14ARF increased expression of TIMP-3 in human glioblastoma cell line is sp1 dependent. Knockdown of sp1 by siRNA suppressed TIMP-3 promoter activity that is enhanced by P14ARF. 34 Other studies also demonstrated that sp1 regulated TIMP-3 promoter transcription activity via the ERK pathway.33,35 Treatment of ERK inhibitor decreased binding ability of sp1 to DNA. 35 TIMP-3 is also a target for Smad pathway mediated by transforming growth factor (TGF)-β. Qureshi et al. suggested that the transcription factors Smad2 and Smad4 must bind to the promoter of TIMP-3 in the presence of TGF-β. 36 In addition, TIMP-3 expression can also be regulated by histone modification such as histone acetylation and histone methylation. Shinojima et al. used chromatin immunoprecipitation and showed that transcriptional repression of TIMP-3 was associated with increased H3K27me3 and decreased H3K9ac histone marks at TIMP-3 promoter. 37 Many proteins have also been reported to be involved in the process of histone modification. HDAC9 is one of the histone deacetylases (HDACs) that has been indicated to suppress TIMP-3 via promoter histone hypoacetylation. 38 KDM1A, also known as LSD1, caused TIMP-3 repression through H3K4me2 demethylation at TIMP-3 promoter. 39 The enhancer of zeste homolog 2 (EZH2), which has histone methyltransferase activity, is known to reduced TIMP-3 expression by catalyzing H3K27me3. 40
MMP inhibitory activity of TIMP-3
TIMPs are endogenous inhibitors of MMPs and exhibit marked antiproteinase activity against MMPs, ADAMs, and ADAMTSs. 41 TIMPs can use the N-terminal region to bind to the catalytic domain of MMPs to inhibit their activity and form a stable bond with the C-terminal hemopexin domain of proMMPs via the C-terminal region. 42 However, the extent of MMP inhibition differs between each TIMP; TIMP-1 strongly inhibits MMP-9 but poorly inhibits MT1-MMP, MT3-MMP, MT5-MMP, and MMP-19, 30 and TIMP-2 strongly inhibits MMP-2 and can inhibit other MMP members. TIMP-1, TIMP-2, and TIMP-4 inhibit only a few ADAMs.43–45 In addition, TIMP-2 can form a ternary complex composed of TIMP-2-pro-MMP-2-MT1-MMP, which resulted in the activation of pro-MMP-2. 30 TIMP-4 can also form a TIMP-4-pro-MMP-2-MT1-MMP complex, but unlike TIMP-2, leading to inhibit the activation of pro-MMP-2 via inhibition of MT1-MMP. 46 TIMP-3 can form a similar terminal complex to inhibit pro-MMP-2 activation. Knockout of TIMP-3 in cell promoted activation of pro-MMP-2 mediated by MT1-MMP. 47 In contrast to other members of the TIMP family with limited inhibitory activity for ADAMs, TIMP-3 can effectively inhibit ADAM10, ADAM12, ADAM17, ADAM28, ADAM33, ADAMTS-1, ADAMTS-2, ADAMTS-4, and ADAMTS-5. 30 For instance, the ECM protein-degrading activity of ADAM12 can only be blocked by TIMP-3, but not by TIMP-1, TIMP-2, and TIMP-4. 48 In addition, TIMP-2 and TIMP-3 inhibit the aggrecanase activity of ADAMTS1, but TIMP1 and TIMP4 have no significant inhibitory effect. 49
TIMP-3 in cancer
Although TIMPs may have substantial roles in cancer, the mechanisms and outcomes for each TIMP vary. For example, high TIMP-1 expression has been reported to be associated with poor prognosis in most cancers. In an in vitro study, TIMP-1 expression was found to promote cancer cell survival in acute myeloid leukemia (AML) and melanoma.50,51 Clinical studies have shown that a high level of TIMP-1 is associated with a shortened relapse-free and cancer-specific survival in endometrial carcinoma 52 and poor overall survival in laryngeal squamous cell carcinoma. 53 Both increased and decreased expressions of TIMP-2 and TIMP-4 have been reported to be associated with greater cancer risk. Overexpression of TIMP-2 has been reported to increase proliferation of choriocarcinoma cells. 54 By contrast, TIMP-2 inhibits angiogenesis via an MMP-independent mechanism. 55 TIMP-4 is expressed in some cancers; however, its role in carcinogenesis remains unclear. In a recent study, TIMP-4 was found to regulate stemness in cervical cancer by enriching tumor progenitor cells. 56 Studies in human cancer tissues have suggested that TIMP-3 may play a tumor suppressive role, and that TIMP-3 gene expression is downregulated in brain tumors, esophageal adenocarcinoma, gastric adenocarcinoma, clear cell renal cell carcinoma, meningiomas, and pancreatic endocrine tumors.57–61 In an in vitro study, TIMP-3 expression was observed to induce cancer cell apoptosis in cervical cancer, fibrosarcoma, and breast cancer cell lines. 62 In an animal model, TIMP-3-deficient prostate tumors showed increased expression of inflammation markers, including monocyte chemoattractant protein (MCP) 1, COX2, tumor necrosis factor (TNF)-α, and IL-1β. 63 In addition, TIMP-3 has been widely described as a potential angiogenesis inhibitor. A study reported that purified and renatured TIMP-3 inhibits angiogenesis, as revealed by the CAM assay. 64 Moreover, TIMP-3 can inhibit the migration and invasiveness of cancer cells in vitro.65–67 By contrast, some reports have suggested that TIMP-3 promotes cancer. Higher TIMP-3 mRNA levels were observed in the stroma of head and neck cancer cells than in the normal epithelial cells, and high levels of TIMP-3 have been shown to be associated with a significant reduction in the overall survival rate. 68 Our previous study also revealed that plasma TIMP-3 is a potential biomarker of the tumor stage in patients with oral squamous cell carcinoma. 21 TIMP-3-knockout mice demonstrated resistance to developing breast and liver cancer despite the increase in their inflammatory response.69,70
Genetic and epigenetic regulation of TIMP-3 in cancer
Gene polymorphism
Gene expression may be regulated by gene polymorphisms, which are variations in the DNA sequence resulting from a >1% nucleotide change within a population.71,72 Polymorphisms of TIMP-3 have been reported in many cancers including adenocarcinoma, bladder cancer, breast cancer, hepatocellular carcinoma, oral cancer, and prostate cancer.23,73–77 The TIMP-3 polymorphic rs9862 allele is associated with increased plasma levels of TIMP-3 and higher risk of oral cancer than with the wild-type allele. 23 In adenocarcinoma of the gastroesophageal junction, polymorphisms (rs130274, rs715572, rs1962223, and rs5754312) in TIMP-3 and polymorphism rs9862 in the TIMP-3 promoter are associated with survival. 77 Moreover, rs8136803 (TT) is associated with decreased disease-free survival in breast cancer patients. 78
DNA methylation and histone modification
Loss or downregulation of TIMP-3 expression has been linked to promoter hypermethylation of the TIMP-3 gene in several types of cancer including esophageal adenocarcinoma, head and neck squamous cell carcinoma, ovarian cancer, and pancreatic endocrine tumors.27,61,79 Hypermethylation in the promoter region usually causes transcriptional silencing because it affects the ability of the transcription factor to bind to the target gene. TIMP-3 methylation in the sp1 binding site and the TATA box of the promoter are associated with low expression of TIMP-3 protein in gastric cancer cell lines. 80 DNA methyltransferases (DNMTs) are key enzymes causing gene methylation, and dysregulation of DNMTs has been reported in tumorigenesis. 81 Knockout of both DNMT1 and DNMT3B in colorectal carcinoma cell lines caused gene demethylation of TIMP-3, which resulted in recovery of TIMP-3 mRNA expression. 82 TIMP-3 expression is also regulated by ten–eleven translocation 1 (TET1), a dioxygenase involved in cytosine demethylation. TET1 can maintain the expression of TIMP-3 by inhibiting its methylation and, thus, suppressing cancer cell invasion. 83 Loss of TIMP-3 expression can also be regulated by histone H3K27 methylation via upregulation of the EZH2 in non-small cell lung cancer (NSCLC). 40 In addition, TIMP-3 expression was suppressed by KDM1A, a histone demethylase that removes H3K4me2 from TIMP-3 promoter and promotes tumor cells invasion in NSCLC. 39 In prostate cancer, treatment with histone methylation inhibitor 3-deazaneplanocin A and trichostatin A restored expression of TIMP-3. 37
MicroRNA and long noncoding RNAs
Accumulating evidence suggests that microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) affect cancer development. 84 MiRNAs involved in the post-transcriptional regulation of TIMP-3. The mechanism of MicroRNA to suppress gene expression is by controlling mRNA stability and translation through base pairing to the 3′ untranslated region (3′-UTR). TIMP-3 regulation by miRNAs including miR-17-3p, miR-17-5p, miR-21, miR-21-5p, miR-181b, miR-191, miR-221, miR-222, and miR-373 has been widely reported in different cancers (Table 1).85–101 For instance, mature miR-17-5p and the passenger strand miR-17-3p promoted prostate cancer growth and invasion by targeting TIMP-3. 85 Upregulation of miR-21 was correlated with decreased TIMP-3 expression in patients with breast cancer, cervical cancer, cholangiocarcinoma, and pancreatic ductal adenocarcinoma.87,88,90,91 In an in vitro analysis, miR-373 could induce esophageal squamous cell carcinoma cell migration and invasion by inhibiting TIMP-3 expression. 101 In an in vivo study, miR-181b upregulation by TGF-β reduced TIMP-3 expression, and transfection with anti-miR-181b in the hepatocellular carcinoma cell line SK Hep-1 suppressed tumor growth in nude mice. 94 Some studies have suggested that miRNAs targeting TIMP-3 are also crucial in cancer resistance development. For instance, miR-21-5p was upregulated in patients with gastric cancer and induced drug resistance to doxorubicin by targeting TIMP-3. 92 Upregulation of miR-221 in oral squamous cell carcinoma cells increases resistance to doxorubicin by silencing TIMP-3 expression. 97 Moreover, miR-221 inhibition enhances the sensitivity of human oral squamous cell carcinoma cells to doxorubicin by upregulating TIMP-3 expression 98 (Figure 1). lncRNAs are RNA transcripts that are longer than 200 nucleotides. Accumulating evidence indicates that lncRNAs play critical roles in tumorigenesis through various mechanisms such as transcriptional, post-transcriptional, and epigenetic regulation.102–105 The lncRNA BC032913 enhances TIMP-3 expression and inhibits nuclear β-catenin expression, thus suppressing the migration, invasion, and metastatic potential of colorectal cancer cells. 106 The lncRNA DANCR suppresses TIMP-3 expression by increasing the binding ability of EZH2 and H3K27me3 to the TIMP-3 promoter in prostate cancer. 107
MicroRNAs target TIMP-3 during cancer progression.
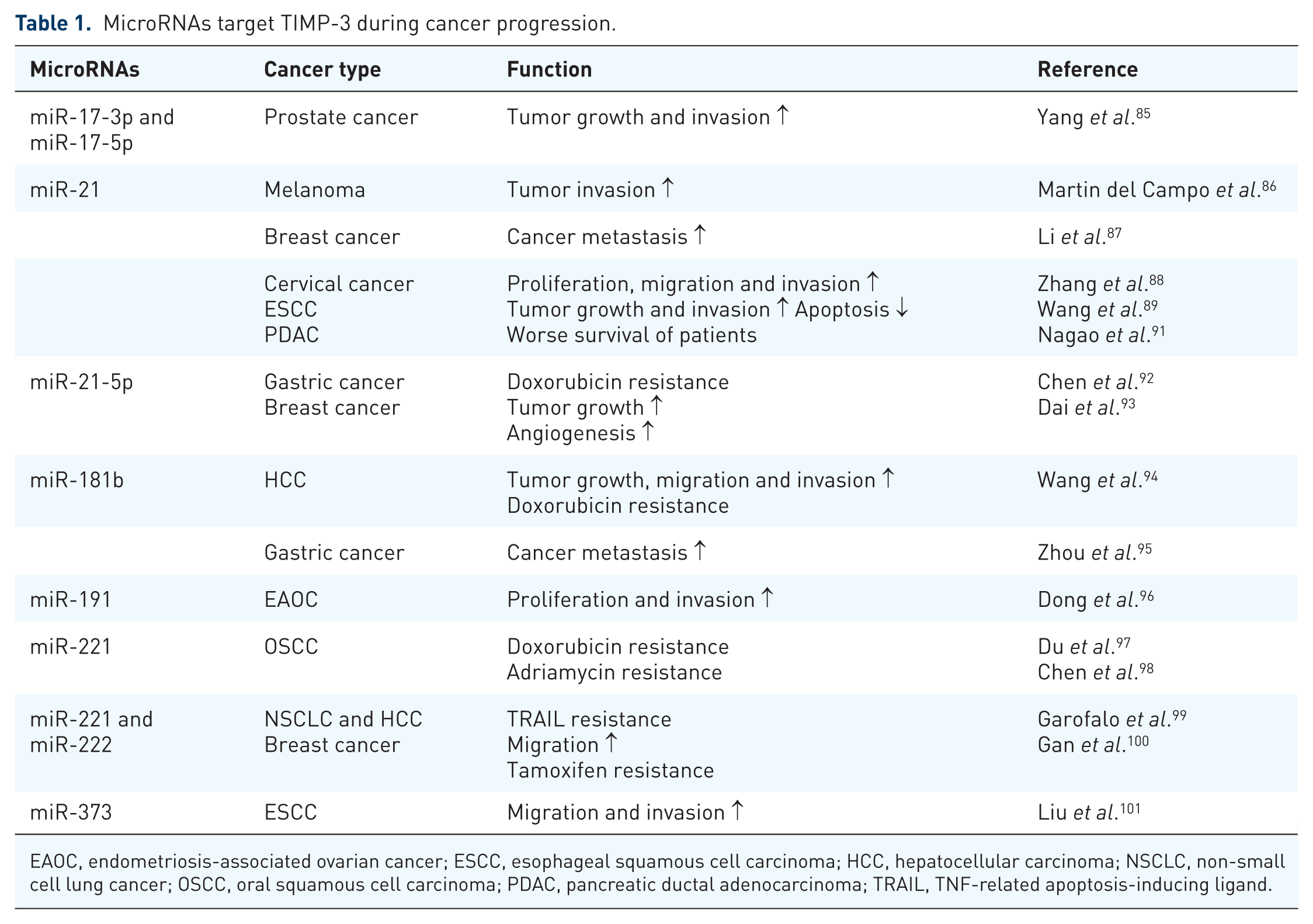
EAOC, endometriosis-associated ovarian cancer; ESCC, esophageal squamous cell carcinoma; HCC, hepatocellular carcinoma; NSCLC, non-small cell lung cancer; OSCC, oral squamous cell carcinoma; PDAC, pancreatic ductal adenocarcinoma; TRAIL, TNF-related apoptosis-inducing ligand.

Function of TIMP-3 in cancer
TIMP-3 functions as a tumor suppressor gene in many types of cancer. It exerts anticancer effects through mechanisms such as the inhibition of cell proliferation, the induction of apoptosis, the inhibition of drug resistance of cancer and inhibition of angiogenesis, migration and invasion, cancer metastasis, and epithelial–mesenchymal transition (Figure 2). In this section, we focus on the aforementioned anticancer abilities of TIMP-3.

Cell proliferation and apoptosis
Cancer cells grow in an uncontrolled fashion and evade the host immune system. Overexpression of TIMP-3 suppresses proliferation and induces apoptosis in different cancer cell lines. 62 TIMP-3 has been shown to induce cell apoptosis by activating mitochondrion-mediated caspase-3 in highly metastatic prostate cancer cell lines PC-3 and DU-145. 108 In addition, TIMP-3 can enhance the sensitivity of cancer cells to apoptosis by stabilizing the death receptor. Mark et al. reported that TIMP-3 induces a type II apoptotic pathway via a Fas-associated death domain-mediated mechanism. 109 TIMP-3 overexpression in melanoma cells induces apoptosis by stabilizing the TNF receptor-1 (R1), FAS, and TNF-related apoptosis-inducing ligand receptor-1 (TRAIL-R1) on the cell surface; stabilization of death receptors results in the activation of apoptosis markers caspase-8 and caspase-3. 110 Other studies have also suggested that TIMP-3 can block ADAM17 activity, which induces shedding of TNF-R1 and TNF-α from the cell surface.111,112 Notably, TIMP-3 can also promote death of nonadherent small-cell lung carcinoma cells even if the cells do not present cell surface death receptors or caspase-8. 113 In vivo studies have shown that prostate cancer cells transfected with TIMP-3 suppress tumor growth and induce tumor apoptosis in nude mice. 114 Similar results have also been shown in colon carcinoma, melanoma, and neuroblastoma; TIMP-3 overexpression suppresses tumor growth in vivo.15,115,116
Angiogenesis
Angiogenesis, which is the process of new blood vessel formation, plays a key role in tumor growth and metastasis. TIMP-3 has been shown to inhibit angiogenesis through regulating angiogenesis-related proteins or directly inducing apoptosis of endothelial cells. Vascular endothelial growth factor (VEGF) is a key mediator of blood vessel development. TIMP-3 can inhibit angiogenesis by blocking VEGF from binding to VEGF receptor-2. 117 In leukemia cells, TIMP-3 can inhibit the proliferation and migration of human umbilical vein endothelial cells (HUVECs) and reduce VEGF-mediated MMP-2 and MMP-9 expression. 67 Kang et al. used a yeast 2-hybrid system and found a TIMP-3-interacting partner angiotensin II type 2 receptor (AGTR2). They suggested that combination treatment with TIMP-3 and AGTR2 inhibits VEGF-mediated proliferation of HUVECs. 118 TIMP-3 can also inhibit angiogenesis by promoting endothelial cell apoptosis. Qi et al. suggested that TIMP-3 may induce apoptosis of endothelial cells by triggering a FAK/Paxillin cell survival pathway but not a caspase-dependent cell death pathway. 119 Upregulation of TIMP-3 by transcription factor sp1 via P14ARF inhibited endothelial cell migration and vessel formation. 34 In an in vivo study, TIMP-3 knockout in nude mice but not in the tumor showed enhanced growth of tumor and increased angiogenesis. 120
Migration and invasion and metastasis
Metastasis is the process of cancer cells breaking apart from the primary tumor and traveling through the blood or lymph vessels to form new tumors in other organs or tissues.2,7,121,122 Metastasizing cancer cells can secrete proteinases to degrade the ECM and further migrate and invade into the blood. Overexpression and unrestricted activation of MMPs may promote malignant conversion of cancer cells.123,124 TIMP-3 has been shown to block proteinase activity and inhibit tumor migration and invasion. Overexpression of TIMP-3 in osteosarcoma decreases MMP-1 and MMP-2 expression and suppresses cell migration and invasion ability. 66 In addition, TIMP-3 can protect ECM degradation mediated by ADAM12, the expression of which is correlated with the status and stage of breast cancer. 48 Inhibition of the invasive activity of melanoma cells is more pronounced in the case of TIMP-3 overexpression compared with TIMP-1 and TIMP-2 overexpression. 16 Loss of TIMP-3 promotes NSCLC cell invasion via TNF-mediated IL-6 production. 24 High levels of soluble CD44 are associated with malignant cancer and metastasis; however, TIMP-3 potentially inhibits CD44 shedding by targeting ADAM-like proteases and MT1-MMP.125,126 In an animal study, melanoma and lymphoma cells in TIMP-3-/- mice were observed to have higher metastatic ability, metastasizing to multiple organs, and lung tissues from TIMP-3-/- mice showed higher MMP-2 and MMP-9 enzyme activity than did those from wide-type mice. 127
EMT
EMT refers to the transition of an epithelial cell into a mesenchymal cell and is considered an important indicator of cancer cell metastasis.128,129 Restoration of TIMP-3 expression in thyroid tumor cells increased their cell adhesion ability, thereby increasing the expression of the epithelial marker cytokeratin 8/18 and decreasing that of the mesenchymal marker vimentin. 19 In addition, transfection of pcDNA-TIMP-3 in gastric cancer cells followed by treatment with TGF-β revealed that TIMP-3 partially rescues EMT-related marker expression induced by TGF-β. 95
TIMP-3 as a target for cancer therapy
In addition to traditional surgery, chemotherapy, and radiotherapy, current cancer research is increasingly focusing on identifying biomarkers for early prediction of cancer progression, immunotherapy, and the use of targeted therapy against tumor suppressor genes or oncogenes.
TIMP-3 as a biomarker for predicting cancer progression
Cancer may rapidly progress from an early stage to an advanced stage, which hinders early diagnosis and accurate monitoring of progression. Plasma and serum levels of TIMP-3 can be easily detected using enzyme-linked immunosorbent assay. A clinical study showed that the increase in plasma levels of TIMP-3 was significantly higher in those with large tumors (>T2) than in those with small tumors among betel quid chewers with oral cancer. 23 TIMP-3 protein and mRNA can be extracted from tissues of patient with cancer and detected using Western blotting, immunohistochemistry, and real-time polymerase chain reaction. TIMP-3 protein expression has been shown to be correlated with tumor stage in NSCLC. 24 Low TIMP-3 mRNA expressions was observed in patients with high-grade clear cell renal cell carcinoma. 59 Loss of TIMP-3 expression in gastric cancer tissues is associated with tumor size, histologic grade, lymphatic invasion, venous invasion, invasive depth, lymph node metastasis, distant metastasis, and TNM stage. 25 Gene polymorphism and DNA methylation could be identified from DNA extracted from blood, body fluids, and salivary rinse and used as biomarkers to predict cancer progression in many cancers. TIMP-3 polymorphism rs9862 has been associated with an increased risk of developing a tumor of size >T2 among betel quid chewers with oral cancer. 23 A study revealed that TIMP-3 hypermethylation in patients with esophageal squamous cell carcinoma is associated with poorer prognosis for both disease-free and overall survival than that of patients without TIMP-3 methylation. 130 Patients with AML harboring methylation of TIMP-3 show higher frequency of adverse cytogenetic prognosis than those with a favorable or intermediate prognosis. 131 TIMP-3 promoter hypermethylation detected from pretreatment salivary rinse is significantly associated with local recurrence-free survival in patients with head and neck squamous cell carcinoma. 132 Methylation of the TIMP-3 promoter identified from body fluids has been reported to be a useful biomarker for predicting tumor size, differentiation, T stage, lymph node metastasis, distant metastasis, and clinical stage in patients with gastric cancer 25 (Table 2).
TIMP-3 as biomarker in cancer progression.

AML, acute myeloid leukemia; CCRCC, clear cell renal cell carcinoma; ESCC, esophageal squamous cell carcinoma; HNSCC, head and neck squamous cell carcinoma; NSCLC, non-small cell lung cancer.
Combination of TIMP-3 and clinical treatment in cancer therapy
Cancers cells can develop drug resistance, which could limit the efficacy of traditional therapies. Combination therapy, the use of more than one type of therapy in treatment, has the potential to delay or reduce drug resistance of cancer. In prostate cancer, adenovirus-mediated expression of TIMP-3 highly sensitizes prostate cancer cells to the chemotherapeutic drug paclitaxel (Taxol) and promotes synergistic enhancement in cell death. 108 TIMP-3 increases cisplatin-induced apoptosis in laryngeal carcinoma by facilitating a mitochondria-dependent apoptosis mechanism such as cytochrome c release and caspase activation. 133 In addition, TIMP-3 overexpression in osteosarcoma cells facilitates cisplatin-induced apoptosis, whereas TIMP-3 knockdown by siRNA has an opposite effect. 66 In an animal study, combination therapy with adenovirus-TIMP-3 and the broad-spectrum antitumor agent cisplatin inhibited cervical cancer xenograft growth more effectively than cisplatin only. 134
Inhibition of cancer progression through TIMP-3
Because TIMP-3 expression is downregulated or deregulated in cancer, current studies are focusing on normalizing or reactivating the expression of TIMP-3 (Table 3). In recent years, the use of natural products has gained much attention in cancer chemoprevention. For example, Andrographolide, a Chinese herbal medicine that is isolated from the stem and leaves of Andrographis paniculata, can inhibit angiogenesis by suppressing miR-21-5p expression and enhancing TIMP-3 expression. 93 Diallyl disulfide, one of the organosulfur compounds derived from Allium vegetables, can inhibit migration and invasion in gastric cancer and upregulate tumor suppressor gene expression, including that of TIMP-3 and E-cadherin. 135 Green tea polyphenols and their major component epigallocatechin-3-gallate restore TIMP-3 expression by attenuating epigenetic silencing of EZH2 and HDACs, thus inhibiting invasion in breast cancer. 136 Arctigenin, derived from the seeds of Arctium lappa, may also increase TIMP-3 expression and inhibit tumor growth in prostate cancer. 137 Mithramycin A (MMA), an antibiotic against Gram-positive soil bacteria, has antitumor and antimetastatic effects; MMA can interact with the catalytic pocket of DNMT1, which results in promoter demethylation and mRNA restoration of TIMP-3. 138 Although beneficial, cancer chemoprevention is associated with many challenges, such as toxicity, side effects, efficacy only at high doses, low bioavailability, and rapid metabolism. 139 In addition to the natural products, some synthetic products can also be used against cancer. NucAnt 6L (N6L), a synthetic peptide, can suppress the invasion ability of melanoma through TIMP-3 release from sulfated glycosaminoglycans. 140 The synthetic peptide p700, which is derived from the N-terminal domain of TIMP-3, inhibits VEGF-family receptors, angiogenesis, and tumor growth. 141 An artificially designed i-lncRNA can target multiple oncogenic miRNAs, thereby protecting the tumor suppressor gene TIMP-3 in diffuse large B-cell lymphoma. 142 TAPI-0, a functional analog of TIMP-3, which inhibits AML cell-induced short-term natural killer (NK) cell abnormalities, may contribute to NK cell-based immunotherapy of AML. 143 Other studies have focused on the epigenetic regulation of TIMP-3. Jharna et al. suggest that the quinoline-based compound SGI-1027 acts as a hypomethylation agent by inhibiting DNMT1 activity and inducing its degradation. In addition, they claimed that treatment with SGI-1027 facilitates re-expression of the tumor suppressor genes MLH1, P16, and TIMP-3 in cancer cell lines. 144 MPT0G013, a HDAC inhibitor, has been reported to induce TIMP-3 expression and further inhibit angiogenesis and tumor growth in vivo. 145 Moreover, TIMP-3 may be a target for demethylation treatments in AML patients. TIMP-3 demethylation by azacytidine (AZA) or decitabine (DAC) can inhibit the release of soluble NKG2D ligand (NKG2DL) mediated by ADAM17 and enhance the lytic activity of NK cells through immune recognition mediated by the NKG2D–NKG2DL engagement. 131 Although hypomethylating agents (HMAs) such as AZA and DAC have been widely used in the clinical treatment of AML. However, there are still many limitations and side effects in the use of HMAs, 146 and it is easy to cause excessive inhibition of DNA methylation. Moreover, DNA hypomethylation may cause some autoimmune diseases such as systemic lupus erythematosus (SLE),147,148 and may also activate some cancer metastasis genes such as u-PA to accelerate cancer metastasis. 149 Therefore, second-generation HMAs and other combined treatments are still evolving and ultimately expected to improve efficacy and reduce side effects. 150
Anticancer agents targeting to TIMP-3.

DNMT1, DNA methyltransferase 1; EGCG, epigallocatechin-3-gallate; NK, natural killer; p700, synthetic peptide derived from N-terminal domain of TIMP-3; STNK, short-term natural killer.
Conclusion
TIMP-3 is unique among the TIMP family members because it is the only TIMP that can bind firmly to the ECM after secretion. In addition, TIMP-3 can inhibit not only MMPs but also a wide range of ADAMs and ADAMTSs. Accumulating evidence indicates that the TIMP-3 gene acts as a tumor suppressor gene by inducing apoptosis and inhibiting proliferation, angiogenesis, and metastasis. However, TIMP-3 expression is downregulated by genetic and epigenetic alternation in most cancers. In this review, we have systematically described the contribution of TIMP-3 in cancer and its potential in cancer therapy including its utility as a predictor of cancer progression, its efficacy in combination therapy for cancer treatment, and its potential as a direct target for cancer therapy. Further research is required to harness the potential of TIMP-3 in diagnosing and treating cancer.