
Abstract
Urbanization modifies bird and plant communities, affecting ecological processes such as fruit consumption and the structure of interaction networks. Our study evaluated how the abundances of synanthropic and non-synanthropic birds, along with native and non-native plants, influence fruit consumption and bird-plant feeding networks in 12 urban green areas in Mérida, Yucatán, Mexico. Over a year, we recorded bird-plant interactions, measured species abundances, and analyzed feeding networks using metrics of specialization, nestedness, and evenness. We found that, in Mérida, urban green areas are dominated by synanthropic birds and non-native plants, promoting feeding networks that are more resilient but less specialized. We found that non-native fruit consumption was significantly associated with the abundances of synanthropic birds and non- native plants, while native fruit consumption depended primarily on the abundance of native plants and showed no significant relationships with birds. Feeding networks in areas with higher synanthropic bird abundance were less specialized, more nested, and more even, indicating a homogenization of interactions in urbanized sites. In contrast, non-synanthropic birds and native plants abundances showed weak or non-significant relationships with network structure. While synanthropic birds play an important role in feeding networks in urban environments and may stabilize certain interactions, it poses risks to the conservation of native biodiversity and ecological processes. Therefore, it is essential to implement management strategies that prioritize native species to maintain the ecological functionality of urban green areas.
Keywords
Introduction
Urbanization drastically alters the landscape by fragmenting natural habitats creating urban environments with isolated remnants of vegetation and reduced resource availability.1–3 Although a clear relationship between urbanization and species loss has been established,4,5 there are also some emerging patterns of the effects of urbanization on species interactions. For example, in urban environments, bird species with a generalist diet have been found to function as connectors of multiple nodes in feeding networks. 6 Similarly, some bird species may be able to incorporate resources from the non-native plants that may be abundant in cities. However, native plants maintain essential connections with birds such as species with specialist diets, preserving the complexity and specialization of feeding networks.7,8 Measuring changes in interaction networks along urbanization gradients is important since interactions represent a fundamental process in community dynamics and provide a deeper understanding of species composition in urban habitats.9,10
In cities, green areas are important habitats for many animal species such as birds11,12 because they may mitigate against adverse conditions present in the rest of the city and provide more constant food resources. 13 However, the availability of food resources in green areas can also vary due to the influence of urbanization on bird and plant species composition and abundances. 14 For example, it is common for the diversity and abundance of native plants to be low in highly urbanized green areas, having been replaced with non-native species.15,16 In addition, urbanization reduces the diversity and abundance of many bird species while promoting an increase in the abundance of a small group of “synanthropic” species, those that are able to adapt to urbanized environments and possibly attain greater population sizes compared to natural habitats.17,18 The term “synanthropic” encompasses both urban “exploiters” and “adapters”18,19 and it is broader than simply “urban”, which refers to those species exclusively found in urbanized environments. In contrast, species that have smaller populations or avoid urbanized environments are classified as “non-synanthropic.”.20–22 However, synanthropic responses can vary depending on the urban context. While some species, such as pigeons and sparrows, show a consistently positive response to urbanization, in many others the synanthropic response depends on the specific characteristics of each city, such as resource availability, landscape structure, or the degree of disturbance. 19 It has been suggested that, despite declining in highly urbanized areas, the presence of native vegetation remains essential for non-synanthropic birds because their fruits are an important part of their diet. 23 Non-synanthropic species may not be adapted to consume the fruits of non-native plants,24,25 so they may interact more frequently with native than with non-native plants.26,27 On the other hand, synanthropic birds tend to have a broader dietary niche compared to non-synanthropic birds, 28 so they can take advantage of novel food resources in green areas. 20 Furthermore, non-native plants can produce a greater number of fruits more frequently than native plants, 29 so synanthropic birds may consume non-native plants more frequently than native plants. Understanding whether the influence of bird abundance on fruit consumption varies depending on bird species can provide valuable insights for predicting the impact of species turnover on ecological –particularly feeding– interactions within urban green spaces.
Ecological interaction networks are a useful tool to characterize bird-plant interactions in green areas and detect changes therein. 30 These visual tools represent the relationships and connections between species. They are composed of nodes that represent the species involved, and links that symbolize the interactions between them. 31 Metrics describing interaction network structure provide indicators of species function and the resilience or vulnerability of interactions to disturbances,32,33 making interaction networks useful for species conservation in heavily disturbed environment such as cities. Some studies suggest that variation in the abundances of plants and birds in urban green areas lead to more generalist and nested feeding networks (networks formed primarily by species with several interactions, where species with few interactions are nested within the interactions of generalist species) with fewer interactions compared to networks in natural environments.15,34 However, it remains unclear whether the structure of feeding networks is affected by variation in the abundance of birds and plants together or is independent for each group of species. If interactions are driven by changes from non-synanthropic to synanthropic birds irrespective of changes in the plant community, then the generalization of networks may occur even in urban environments with few non-native plants. Conversely, if interactions are driven by changes from native to non-native plant species irrespective of changes in the bird community, then the generalization of networks might occur even in scarcely urbanized systems.
The purpose of our study was to evaluate the degree to which changes in bird (synanthropic and non-synanthropic) and plant (native and non-native) abundances affect the consumption of fruits and the structure of bird-plant feeding networks. We used 12 green areas in the city of Mérida a rapidly growing city on the Yucatan Peninsula of Mexico. Our hypothesis is that the changes in bird and plant abundances similarly affect native and non-native fruit consumption and the structure of bird-plant feeding networks in green areas. We predict that urban green areas with higher abundances of synanthropic birds and non-native plants will show higher consumption of non-native fruits and lower consumption of native fruits, while sites dominated by non-synanthropic birds and native plants will exhibit the opposite pattern. (Figure 1(a)). In addition, we expect the shift in the bird and plant communities from non-synanthropic to synanthropic species and from native to non-native species, respectively to increase nestedness, decrease specialization, and decrease interaction evenness in bird-plant interaction networks (Figure 1(b)). Conceptual model about the effects of the abundance of birds and plants on the consumption of native and non-native fruit consumption (a) and the structure of interaction networks (b). The black lines and red lines represent positive and negative relationships respectively.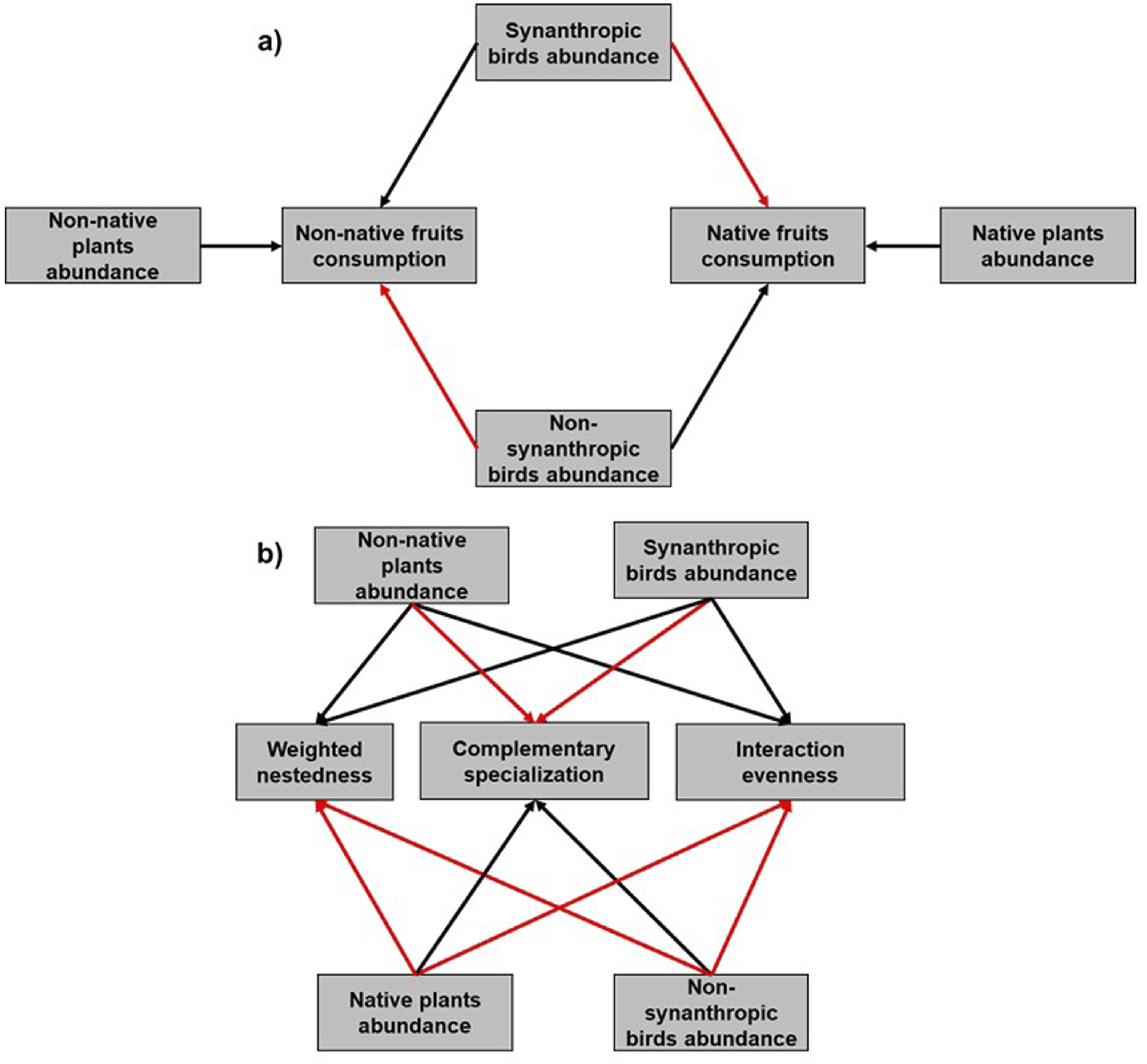
Materials and methods
Study site
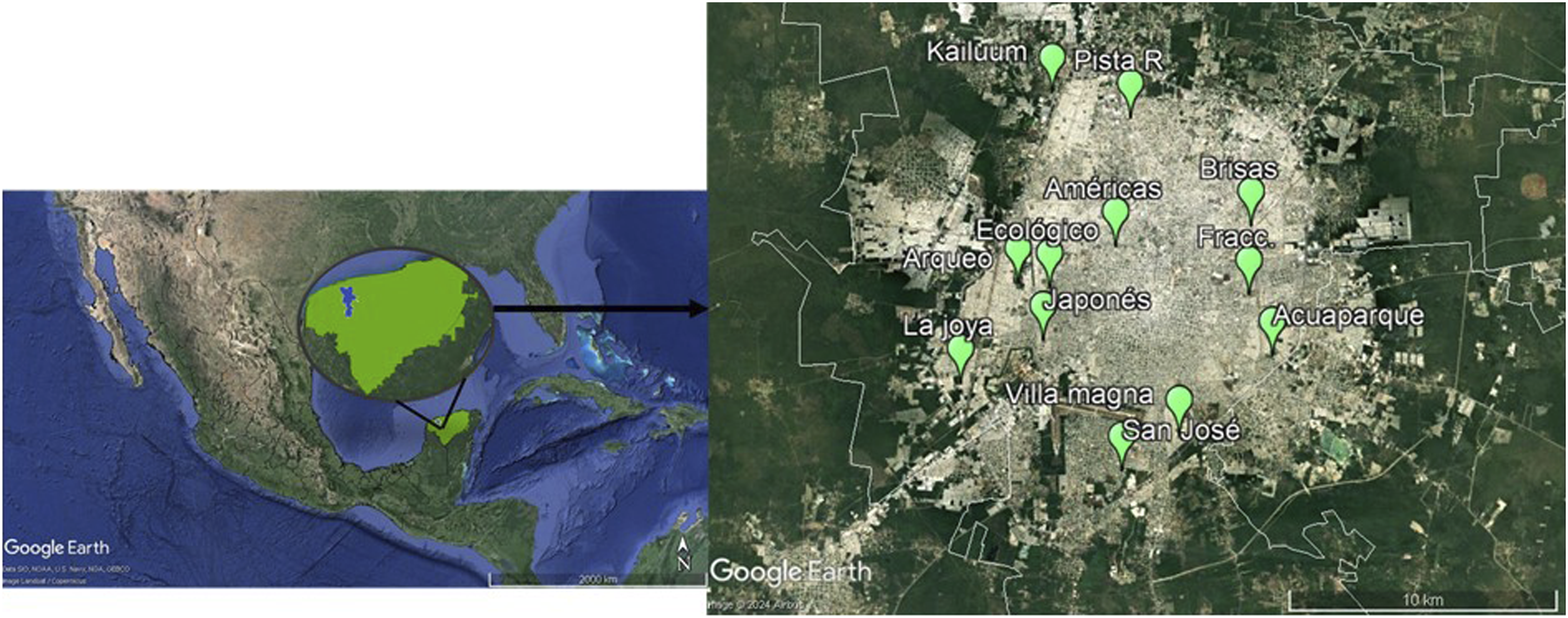
We recorded interactions between birds and plants in 12 green areas located in the city of Mérida (Figure 2, Supplemental Figure 1), located on the Yucatán Peninsula, southeastern Mexico (20°58′ N, 89°37 ′ W; population 995,129.
35
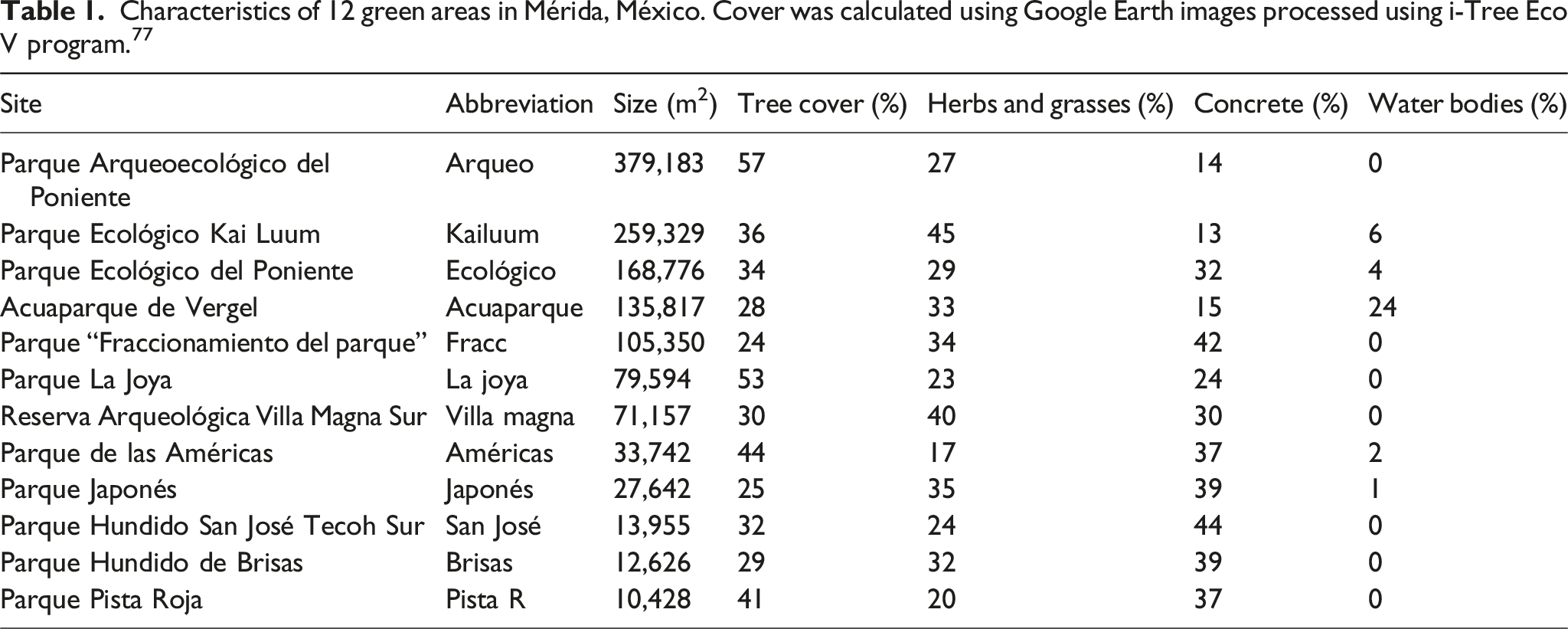
The selected green areas varied in size and land use (Table 1, Supplemental Table 1). The composition and location of the seven urban greenspaces in Merida, Yucatan, Merida. Greater Mérida is the blue polygon embedded in the green polygon indicating Yucatan state, Mexico. Twelve green aeras were chosen as study sites, indicated by the green markers. The images were obtained from Google Earth and modified using shapefiles from CONABIO
78
and the Geographic Metadata Catalog of the Territorial Ecological Planning Program of Yucatán State. Characteristics of 12 green areas in Mérida, México. Cover was calculated using Google Earth images processed using i-Tree Eco V program.
77
Data collection
We visited each site twice a month from August 2, 2021 to September 30, 2022, (28 visits per site, 336 in total). We measured the abundance of all visually-identified bird species by establishing circles of 12 m radius (452 m2 in area), with each point separated by a distance of 100 m. The point size was selected following a previous study in Merida. 36 The number of points per site varied from three to eight depending on the size of each green area and we averaged abundance across the points. We excluded aquatic species from the analysis because they respond to different factors than terrestrial birds. We classified land birds into two groups: “synanthropic” and “non-synanthropic”. 19 Since there is no specific classification of synanthropic birds for the city of Mérida, we relied on a previous study conducted in the city’s urban green areas. 37 The study characterized seven sites using an urbanization index calculated as the weighted average of concrete cover and demographic variables (i.e., human population density and number of buildings and homes). We downloaded the data from Mexico´s Instituto Nacional de Estadística y Geografía (INEGI). We calculated variable weights by first standardizing each variable on a 0–1 scale and then using a principal component analysis to find the contribution of each variable to total inertia. The resulting index ranges from 0 to 1, with values closer to 1 indicating higher levels of urbanization. The study measured bird diversity and abundance in each site based on weekly visits from March 2016 to February 2018 over which a trained observer recorded all birds seen and heard while walking a 1 km transect. For our current analysis, we selected the sites with the highest (UI=0.32) and lowest (UI=0.08) levels of urbanization and classified species as “synanthropic” if their total abundance at the highly urbanized site was at least double that recorded at the least urbanized site. All other species were considered “non-synanthropic”. (Supplemental Table 2).
To measure bird-plant interactions, we generated an event matrix that registered the interactions between birds and plants in each green area. Over a 10-min period at each point, we documented each time we observed a bird feeding on a specific plant. Using these data, we created an interaction matrix that recorded the species of bird that initiated the interaction, whether or not it was synanthropic, the species of plant with which it interacted, the origin of the plant (native or non-native), and the number of fruits consumed. We classified plants as 'native’ or 'non-native’ based on whether or not they are native to the Biotic Province of the Yucatán Peninsula, which encompasses the Mexican states of Campeche, Yucatán, and Quintana Roo and parts of northern Belize and Guatemala. 38 To verify plant distribution, we consulted the catalog of the native flora of the Biotic Province of the Yucatán Peninsula. 39 We calculated the abundance of plants for the species that interacted with birds by adding the number of individuals in each 452 m2 circle. We subsequently determined the total consumption of both native and non-native plants, considering the total number of interactions for each type of plant in each of the study sites throughout the 14 month-long study period. To check whether the samples obtained were able to adequately capture the interactions present in the green areas, we generated rarefaction curves with a 95% confidence interval of the total number of interactions recorded between birds and plants and the days sampled. To do this, we used the “iNEXT” package 40 version 3.0.0.37 in R version 3.3.1 41 .
Feeding networks
Using the “bipartite” package version 2.81 42 in R version. 3.3.1 41 we calculated three metrics indicative of the level of specialization in ecological interaction networks. We calculated Complementary Specialization (H2) to measure the extent to which a node is specialized in interacting with a particular subset of nodes in the other set, compared to a random distribution of interactions. 42 The index ranges between 0 and 1 with higher values indicating more specialized nodes (species). We calculated Weighted Nestedness (WNODF) to indicate the degree to which nodes with few connections connect to more highly connected nodes.43,44 We calculated Interaction Evenness (IE) to quantify the level of homogeneity in the distribution of interactions between nodes in the network. 42 IE is calculated by dividing Shannon´s diversity index of interactions by the total number of species in the network. We used igraph package version 1.2.6 45 in R version. 3.3.1 41 to visualize the interaction networks of each green area.
Data analysis
To test the proposed conceptual model of fruit consumption (Figure 1), we used piecewise structural equation models (SEMs). SEMs test hypotheses with multiple variables using path diagrams. Piecewise SEMs extend traditional SEM models by separating the model into parts or pieces using linear or multiple regressions for each response variable, evaluating them separately and then combining them to generate information about the entire SEM. In addition, they allow detecting significant relationships between the selected variables that were not considered in the original model called “tests of directed separation”. 46 First, we ran a piecewise SEM model for native and non-native fruit consumption using linear mixed-effect models with the abundance of birds (synanthropic or non-synanthropic) and plants (native or non-native) as independent variables and site as a random factor. We also compared the standardized parameter estimates of each variable in the model and the test of directed separation to assess whether there were important relationships in the model path that we did not include in our original conceptual model (Supplemental Table 3).
Second, we ran piecewise SEM models for interaction network metrics using linear mixed-effect models, with the abundance of birds (synanthropic or non-synanthropic) and plants (native or non-native) as independent variables and site as a random factor. We used the PiecewiseSEM package version 2.3.0 46 in R version 3.3.1. 41 We evaluated the fit of the fruit consumption and interaction network models using the Fisher C statistic under the null hypothesis of no difference between the model developed and the observed data when comparing their covariance matrices. 46
Results
We recorded 1,090 interactions between 27 bird species and 23 plant species across all study sites. Of these, 11 bird species were classified as synanthropic and 16 as non-synanthropic. For plants, six species were classified as non-native and 17 as native (Supplemental Tables 2 and 4). The most frequently observed bird species were all synanthropic: Hooded Oriole (Icterus cucullatus) with 150 interactions, Great-tailed Grackle (Quiscalus mexicanus) with 124 interactions, Social Flycatcher (Myiozetetes similis) with 115 interactions, Cinnamon-bellied Saltator (Saltator grandis) with 111 interactions and Clay-colored Thrush (Turdus grayi) with 110 interactions. Together they accounted for 56% of the total interactions. For native plants, Alamo tree (Ficus cotinifolia) and Strawberry tree (Muntingia calabura) had the greatest number of interactions with 125 and 111, respectively. For non-native plants, the species with the highest number of interactions (148) was Weeping fig (Ficus benjamina).
For each of our sites, the rarefaction curves describing the relationship between the number of bird-plant interactions and the number of sampling events reached an asymptote indicating that additional site visits would yield few new interactions. More specifically, the observed interaction richness represented more than 85% of the estimated asymptotic richness. The confidence intervals of the rarefaction curves were narrow. (Supplemental Figure 2).
Across urban green areas of Mérida, we expected that sites with higher abundances of synanthropic birds and non-native plants would show increased consumption of non-native fruits. We found that both variables were significantly associated with consumption, though the effect was stronger for bird than for plant abundance (Figure 3). We also found that the consumption of non-native fruits was weakly and non-significantly associated with the abundance of non-synanthropic birds. We also expected that sites with lower abundances of non-synanthropic birds and native plants would have decreased consumption of fruits of native plants. We found that the consumption of native fruits was strongly and significantly related only to the abundance of native plants and was not significantly associated with the abundance of either bird group. (Figure 3). In terms of feeding networks, we found that their structure varied across sites. Generally, more urbanized sites had fewer interacting species and were composed mainly of synanthropic birds and non-native plant species (Figure 4, Supplemental Figures 3,4). As expected, networks in sites with greater synanthropic bird abundance were more nested, less specialized, and more even than feeding networks with fewer synanthropic birds (Figure 5). Contrary to expectations, the abundance of non-synanthropic birds was not significantly associated with feeding network structure. Surprisingly, the abundance of native plants was not significantly associated with feeding network structure (Figure 5). On the other hand, we found that non-native plant abundance was negatively associated with complementary specialization and positively associated with interaction evenness. The relationship with nestedness was not significant. Piecewise SEM results for the effects of bird and plant abundance on native and non-native fruit consumption. The rectangles represent the measured variables. Solid lines represent significant relationships between variables (p ≤ 0.05) while non-significant relationships (p > 0.05) are represented with dotted lines. Positive relationships are marked in black and negative relationships in red. Two-way lines represent the covariances. The numbers represent the values of the standardized coefficients of the regressions between variables. Bird-plant feeding networks and matrices at the sites with the highest (San José) and lowest (Kailuum) percentage of concrete cover. Red squares and blue diamonds represent synanthropic and non-synanthropic birds respectively, named by their American Ornithological Society code. Native plants are represented with green circles and non-native plants are yellow triangles and assigned a four-letter code made of the first two letters of the genus and first two letters of the species. The thickness of the lines is proportional to the frequency of the interaction. The matrices show bird species on the x axis and plant species on the y-axis. Each box in the matrix represents an interaction between a bird and a plant while the gray scale represents the frequency of feeding interactions, with darker shading indicating more frequent interactions. San José has a simple matrix with few species and no modules while Kailuum has a more complex matrix with more species and up to five modules. Bird-plant feeding networks of all sites are shown in Supplemental Figures 3 and 4. Piecewise SEM results for the effects of bird and plant abundance on feeding network metrics in green areas of Mérida. The rectangles represent the measured variables. Solid lines represent significant relationships between variables (p ≤ 0.05) while non-significant relationships (p>0.05) are represented with dotted lines. Positive relationships are marked in black and negative relationships in red. The numbers represent the values of the standardized coefficients of the regressions between variables. WNODF= Weighted nestedness, H2= Complementary specialization, IE= Interaction evenness.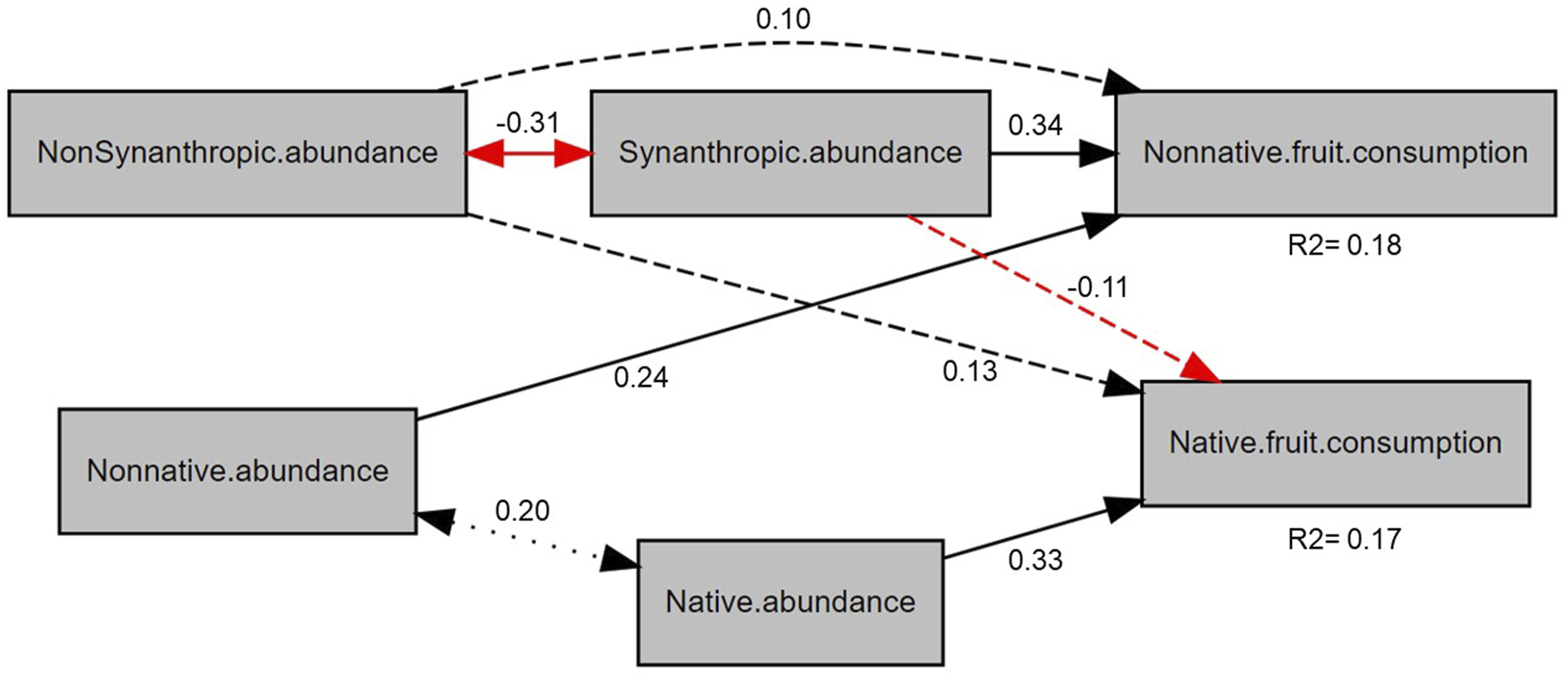
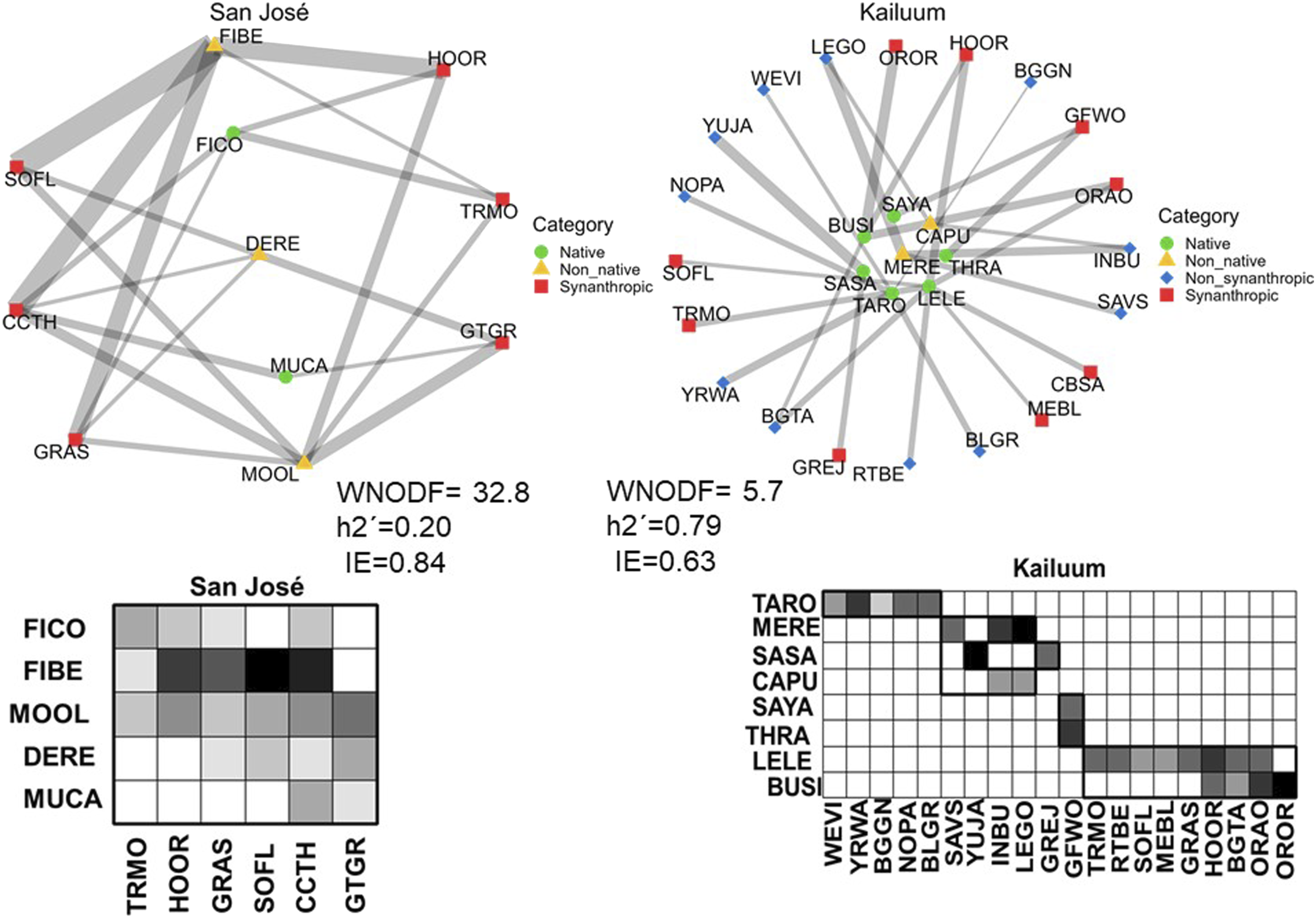
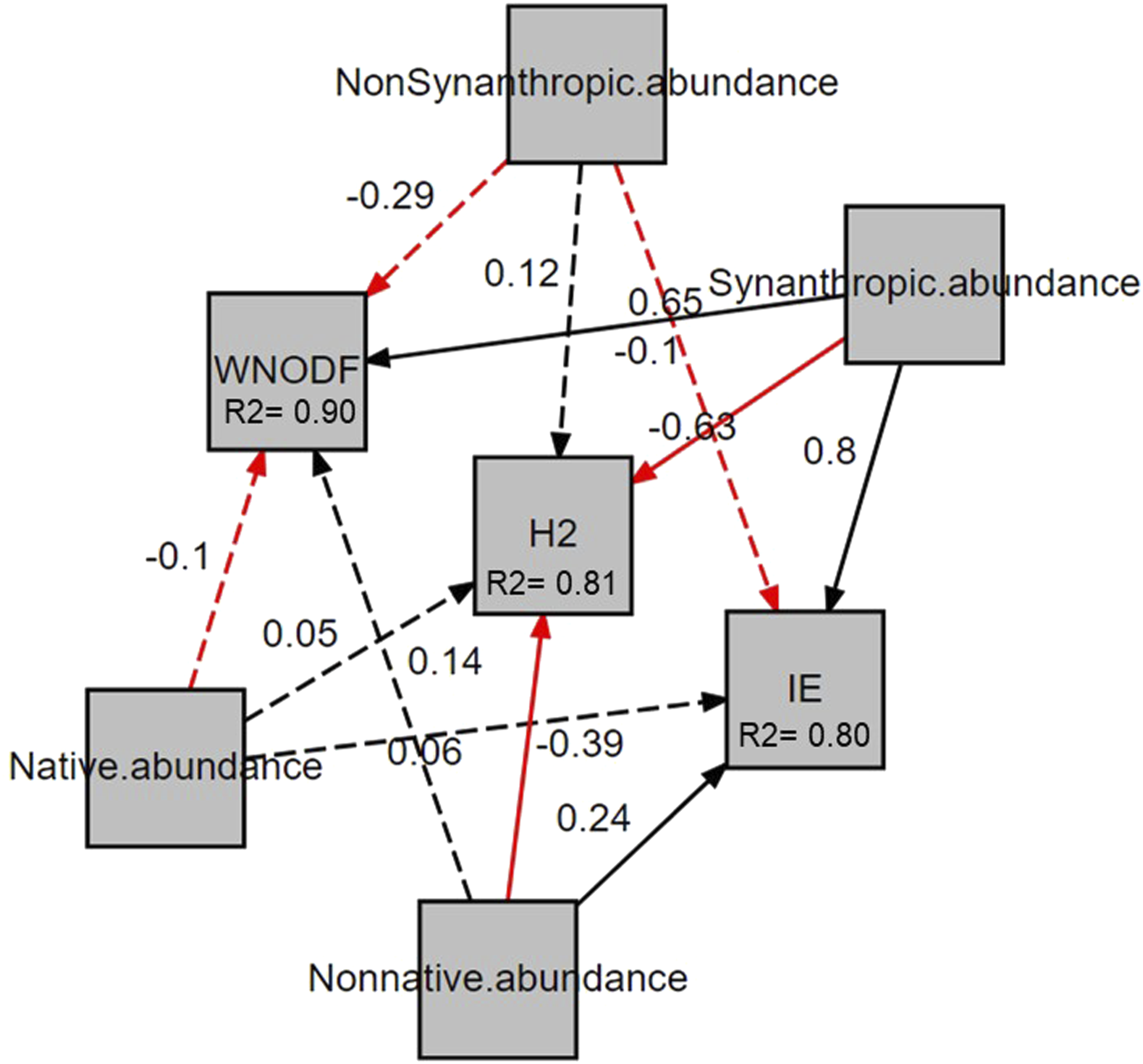
Discussion
We proposed that the consumption of fruits of native and non-native plants by birds in green areas of Mérida, Mexico would vary as a function of a shift in the bird and the plant communities from being dominated by non-synanthropic bird species and native plants to synanthropic bird species and non-native plants. We also expected that variation in bird and plant abundances would affect the structure of bird-plant feeding networks. As expected, non-native fruit consumption in green areas was associated with changes in synanthropic bird and non-native plant abundance. Contrary to expectations, native fruit consumption was not strongly associated with bird abundance, whether non-synanthropic or synanthropic species. Instead, native fruit consumption was associated primarily with changes in plant abundance. While other studies have shown that non-synanthropic birds prefer fruits from native plants even when non-native fruits are present,47,48 we did not find evidence for a similar pattern in our system.
The weak association between bird abundance and native fruit consumption could occur because consumption rates are dictated by fruit nutritional content. If fruits have a low energy content, many birds may prefer to feed on other resources. 49 In general, many bird species may forage on fruit only opportunistically 50 and focus most of their foraging effort on consuming insects or seeds. 51 In our study, none of our species are exclusively frugivorous, so fruit consumption rates in a particular urban green area may be a byproduct of how often birds are finding and foraging on other resources. As well, native fruit production tends to be seasonal, peaking at only a few points during the year.52,53 While bird abundance may be associated with fruits during those peaks, the relationship may be diluted by our analysis that did not take into account temporally varying bird-fruit relationships. Finally, bird species forage within breeding territories established around native trees.54,55 If the territory holder successfully excludes competitors, then just one individual might be responsible for the majority of fruit consumption. Furthermore, differences in average territory size and movement patterns between synanthropic and non-synanthropic species could mean that some birds are consuming fruits at scales larger than we could observe. 56 Although we did not directly measure territory size or spatial use by birds, we recognize that incorporating this dimension into future research will provide a more comprehensive understanding of resource use and interaction network dynamics in urban landscapes.
Unlike native fruits, we did find a strong relationship between non-native fruit consumption and synanthropic bird abundance. The finding that synanthropic bird species dominate highly urbanized areas facilitating the consumption of non-native fruits is known from studies of other tropical cities4,57,58. Synanthropic birds take advantage of the novel resources provided by non-native plants and incorporate them into their diet more readily than non-synanthropic birds.59,60 Moreover, synanthropic birds may favor non-native over native fruits because even though we only registered a few individuals of non-native plants, they tended to produce a large number of fruits, especially when summed over the 14 month-long study period. Thus, birds can exploit a high-energy resource without having to expend a lot of energy searching or competing for the resource. These results are consistent with other studies indicating that dietary preferences may have a greater influence on fruit selection by frugivorous birds. 61 Synanthropic birds may exploit non-native plants more intensively due to their higher quality or accessibility in urban environments.62,63 None of the non-native plants recorded in this study are classified as invasive species. Rather, their presence in urban green areas is primarily due to their ornamental value.
We found that the structure of bird-plant feeding networks was associated more strongly with synanthropic bird and non-native plant abundance compared to non-synanthropic birds and native plants. Sites with higher synanthropic bird and non-native plant abundance exhibited less specialized and more nested networks with more uniform interactions. The shift in structure we observed has been reported in various studies documenting more homogeneous bird-plant feeding networks in highly urbanized sites.64,15 Networks in green areas with high levels of urbanization are usually composed of a low number of generalist birds that interact with all or most of the plants in the network,65,15 so networks tend to be more nested with interactions more evenly distributed compared to networks present in natural environments.66–68 The positive association between synanthropic bird abundance and network evenness that we observed indicates that networks with a higher abundance of synanthropic birds may be more resilient to disturbance compared to those with low interaction evenness.69,70
Interestingly, we found only weak and non-significant relationships between feeding network structure and the abundance of non-synanthropic birds and native plants. Some studies suggest that, in fragmented forests, abundant and small-sized birds like non-synanthropic birds are central to interaction networks. 71 However, the presence of non-synanthropic birds in green areas is influenced by both local and landscape factors, such as area size and the presence of native forest.54,72 Small green areas or areas with little native vegetation cover reduce the abundance of non-synanthropic birds and their role in the networks. Furthermore, a higher abundance of synanthropic birds may directly or indirectly reduce the presence of non-synanthropic birds and limit their participation in feeding networks. 73 Although it was not an objective of our study, we further explored the influence of bird and plant species richness on network metrics. Interestingly, and in contrast with abundance-based models, we observed that non-synanthropic bird species richness had a significant and opposite effect on network metrics compared to synanthropic bird abundance (Supplemental Figure 5). These results are consistent with other studies suggesting that, although the abundance of non-synanthropic birds is limited, their diversity increases the specialization and complexity of networks. 74 This finding underscores the importance of preserving the diversity of non-synanthropic birds in urban landscapes, as their presence can contribute to maintaining more structured and less homogenized feeding networks, even when they are not numerically dominant.
The relationship between native plants and bird-plant feeding networks may depend on the characteristics of each urban environment, making it difficult to find general patterns. In some cities, the presence of native plants in feeding networks remains constant. On the other hand, some cities have networks where native plants are replaced by non-native species.26,75 In addition, the phenology of native and non-native plants are different, so their role in urban interaction networks differs. Native plants are more important at times of peak fruit production, which is limited to certain months of the year. Meanwhile, the presence of non-native plants in the networks is constant, likely reflecting a more constant fruit production throughout the year. 76 In our study, we considered the feeding networks of green areas for the whole year and not by season, which may explain the dominance of non-native plant species in urban interaction networks. We suggest that future research measure directly fruit abundance within and between seasons to better understand how temporal heterogeneity influences feeding networks in urban green spaces. Finally, it is possible that plant size has a stronger influence on fruit consumption than plant abundance, especially for the native plant species recorded in this study, since they were highly dominated by trees and large palms (15 out of 17 plant species), compared to non-native plants (3 out of 6 species; see Supplemental Table 3).
Our study only assessed the effects of bird and plant abundance in a single city and at a local scale, such as in well-defined green areas. Considering the landscape effect and comparing interactions between different cities can help to understand the effect of changes in bird and plant abundance on fruit consumption and interaction networks. The study of bird-plant interaction dynamics in urban environments points to the importance of habitat characteristics, bird community composition, and the diversity of food resources on the frequency and structure of feeding networks.33,64 Our study is a first approach to understand how shifts in bird and plant communities in urban areas of the Yucatan Peninsula, particularly the city of Mérida, influences food preferences and the structure of bird-plant interactions. In this regard, our results highlight the increasingly dominant role of synanthropic bird and non-native plant species and their homogenizing effect on bird-plant feeding networks in urban green areas with higher levels of urbanization. Determining whether these changes may pose a risk to the ecological functioning of bird and plant communities in cities will allow us to take actions aimed at conserving the ecological processes of urban green areas and maintaining the biodiversity immersed in our cities.
Supplemental Material
Supplemental Material - Synanthropic birds increase their consumption of non-native plants and affect the structure of feeding networks in urban green areas of Mérida, Yucatán, México
Supplemental Material for Synanthropic birds increase their consumption of non-native plants and affect the structure of feeding networks in urban green areas of Mérida, Yucatán, México by Gerardo Iván Linares-Hernández, Juan Manuel Dupuy-Rada, Víctor Parra-Tabla and Richard Evan Feldman in Avian Biology Research
Footnotes
Author contributions
Gerardo Iván Linares Hernández, Juan Manuel Dupuy-Rada, Victor Parra-Tabla and Richard Feldman designed the study. Gerardo Linares Hernández carried out the field research. Gerardo Linares Hernández and Richard Feldman analyzed the data and wrote the manuscript. All authors reviewed and contributed writing to the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following support for the research, authorship, and/or publication of this article: Gerardo Iván Linares-Hernández was supported by a scholarship from Mexico´s Consejo Nacional de Humanidades, Ciencias y Tecnologías.
Data Availability Statement
Please contact authors for the data
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.