
Abstract
Bird nests can be complex bio-engineered structures constructed from a range of materials to provide a site for incubation, and in many species chick rearing. Reports of the materials used in nest walls and cup linings are typically qualitative and do not assist in understanding the functional properties of the structure as a whole. This paper provides size and composition data for nests from four species of the Motacillidae family: the meadow pipit (Anthus pratensis), pied wagtail (Motacilla alba), grey wagtail (Motacilla cinerea), and the yellow wagtail (Motacilla flava), three species of the Sylviidae family: willow warbler (Phylloscopus trochilus), Eurasian reed warbler (Acrocephalus scirpaceus) and Eurasian blackcap (Sylvia atricapilla), and the dunnock (Prunella modularis) of the Prunellidae family. Three hypotheses were tested: do nest dimensions correlate with body mass? Is it possible to distinguish among species based on their nest composition; and for individual species? Is it possible to distinguish between the cup lining and outer nest wall based on the materials used in construction? Nest composition and size varied among species although the degree of intra-specific variation was different. Size of a nest was unrelated to average female adult body mass. Composition relied on similar types of materials but in differing quantities between species. It was possible to distinguish among species in terms of nest construction, and between the cup lining and the outer nest, on the basis of at least one of the component materials. By providing quantitative data on composition, it may be possible in the future to understand better the structural, insulative and hydrological properties of the nest. Such information will be invaluable as we begin to develop an understanding of the factors that have driven evolution of nest architecture.
Introduction
Nest building is a behaviour displayed by most birds, although there is considerable interspecific and intraspecific variation in nest design.1,2 Understanding of the design and function of avian nests has improved over recent years2,3 and has indicated their multifunctional roles. These can include: sexual signalling, 4 regulation of nest microclimate,5-10 structural support,11-12 protection from predators, 13 and pathogenic and parasitic defence.14-15 However, the primary function of a nest is to provide a receptacle for eggs during incubation, and in many cases, chick rearing. 3
The dimensions of the nest, and the specific materials used within the nest, may be important in determining the function of the nest.4,10,12,14,15 The amount of insulating material present in the nest has been linked to the successful incubation and survival of nestlings in many species.6,7,12,16 The presence of feathers within a nest is associated with good insulation, 17 slower rates of cooling of eggs,18-19 and increased nestling growth rate in passerines 20 but experimental heating of nests had no significant effect on the numbers of feathers found in nests of the tree swallow (Tachycineta bicolor). 21
Summary of those species for which comprehensive quantitative data are available about mass of different nest materials. Data sources are provided as references.

This study complements that of Biddle et al., 30 which described nest composition in representative species of the Fringillidae, Turdidae and Muscicapidae, in describing nest composition of seven species of passerine for which quantitative data were not generally available. These include four species of pipit and wagtail (Motacillidae), three species of warbler (Sylviidae), and we provide additional data to those already available 3 about nest composition of the dunnock (Prunella modularis; Prunellidae; here we use the English name reported by del Hoyo 31 ). Ideally, nests should be collected in sufficient numbers across the geographical range of the species of interest to allow a good understanding of intraspecific variation.32,33 However, this is often difficult to achieve due to a lack of systematic collection. Therefore, here we report data from nests that were collected from a range of different sites from around the island of Great Britain, which were all taken as one sample. We feel that, given the paucity of quantitative data about nest composition in bird species, 3 to understand in greater detail the factors that affect inter-specific nest construction behaviour there is a need to report data from as wide a range of species as possible.
In this study we test three hypotheses. First, do nest dimensions positively correlate with average female adult body mass? Secondly, is it possible to distinguish among species based on their nest composition 34 ? Finally, we hypothesise that, for individual species, it is possible to distinguish between the cup lining and the outer nest wall based on the materials used in construction.
Materials and methods
Nests constructed by wild birds were studied from four species representing the Motacillidae: meadow pipit (Anthus pratensis; N = 13), grey wagtail (Motacilla cinerea; N = 5), white wagtail 31 (Motacilla alba; N = 5) and yellow wagtail (Motacilla flava; N = 6). Nest construction was also investigated for three species of warbler (Sylviidae): willow warbler (Phylloscopus trochilus; N = 8), Eurasian reed warbler (Acrocephalus scirpaceus; N = 10) and Eurasian blackcap (Sylvia atricapilla; N = 12), and the dunnock (N = 12). Nests of these species were largely chosen based on availability and had been collected from various locations across Great Britain. The only exception to this was the Eurasian reed warbler; nests were all collected during 2016 at Wicken Fen, Cambridgeshire, UK. As described by Biddle et al. 30 nests were collected by British Trust for Ornithology (BTO) nest recorders. Following guidelines drawn up by the BTO, in situ nests were removed only after the known end of the breeding season (31st August) of 2014, 2015 and 2016. Most nests were posted to the University of Lincoln, where they were frozen at −20°C in a domestic freezer (Hotpoint Ltd., UK) for 4–5 days to kill any invertebrates. 35 Each nest was then air-dried for several days before being placed into a reclosable plastic bag (Waitrose Ltd., UK) of a suitable size not to crush the nest which was then placed into a cardboard box and stored at room temperature and humidity until analysed.
Nests were elliptical in shape so the diameters of the overall nest and cup were measured parallel with, and perpendicular to, the long axis of the cup.12,30,36 Digital callipers (Mitutoyo, Absolute Digimatic, Japan, accuracy ±0.01 mm) were used to measure nest and cup diameters, nest height and cup depth. Wall thickness was measured with callipers at four cardinal points around the nest and averaged for each nest. 36 Electronic scales (A and D Company Limited, USA, model FX-3000i, Sartorius, model Entris 3202i-1S, accuracy 0.01 g) were used to measure the mass (in g) of each nest and its constituent components after deconstruction. The volume of the nest cups was determined by first lining the cup with domestic cling film (Waitrose Ltd., UK) before 5 mm diameter solid glass beads (Sigma-Aldrich, UK) were added until the cup was filled level with the top edge. 11 The mass of the beads was then used to determine the volume of each cup based on a pre-determined density (g cm−3). 11
After all whole nest measurements had been recorded nest deconstruction was carefully carried out using forceps to reduce damage to the materials11,12,30 and to separate materials from the innermost cup lining and outer nest. The walls and base of the outer nest were first removed until the lower limit of the outer part of the cup, recognised by a distinct change in materials, was reached. 12 Individual nest components, as defined by Biddle et al., 30 were extracted using laboratory forceps and separated into broad categories, such as leaves, moss, or hair. The total mass of the nest was determined by summing all of the materials deemed to be used in the nest and excluding other materials, e.g., insects, faeces, pulli bones, that would have been added during incubation and rearing.11,12,30,35
Analysis broadly followed the methodology of Biddle et al. 30 The mean value for each measurement of the eight species was calculated and mean female adult body mass was derived from Dunning. 37 These values were used because the masses of the individual birds that built the nests were not available from the BTO’s Nest Record Scheme (NRS). Interspecific relationships between mean values of variables, i.e., nest mass, wall dimensions, and nest composition, were investigated by calculating phylogenetically independent contrasts (PICs) using the ape package 38 in R3.6.3 39 and a phylogram of species determined using http://birdtree.org. 40 Shapiro tests showed that data were normally distributed so Pearson correlations were run on PICs to control for the possible effect of phylogeny on the relationships between the variables. 41
Analysis was carried out to test for to interspecific differences in composition. Phylogenetically-controlled principal component analysis (PCA) based on a covariance matrix was carried out for the masses of the different nest materials using R3.6.3 39 using the phytools 42 and ape packages. The first two principal components were then used to distinguish the composition of nests for the species. Levene’s tests were carried out in R using the car package 43 on each of the two principal components to determine if the variance was homogeneous among the species. Furthermore, Kruskal-Wallis tests were used in R to determine whether differences between species in the average values for principal component scores were significant.
To compare the composition of the outer nest and the cup lining, a stepwise discriminant analysis (IBM SPSS Statistics 21) was carried out to compare the proportions of each material within the cup lining and the outer nest as a proportion of the cup lining and outer nest for each species, respectively. Proportion data were normalised before analysis using an ASIN-transformation. 44 The significance level was set at an F-value of 3.84, i.e., α threshold of 0.05, which is the default for the test.
Results
Nest dimensions
Mean (±SD) values for nest mass and structural dimensions for the eight species of bird. Sample size is indicated in the square brackets for each species. Wall thickness is given as the mean of all four sides. Spearman’s rho values are at the bottom of each column for the correlation between the mean nest dimensions with mean female adult body mass (data from Dunning 37 ). Nests were collected from a variety of locations from around Great Britain and were obtained from the BTO’s Nest Record Scheme.

General descriptions of nests
Mean (g, ± SE) values for each category of animal or plant material present within the total nest, the cup lining and the outer nest for the species indicated. Hyphen indicates that the materials was not present in either the cup lining or the outer nest.


The mean (+SE) values for the materials used in nests constructed by four members of the Motacillidae. Values are for the different materials expressed as a proportion of (a) the total nest mass, (b) the cup lining mass, and (c) the outer nest mass.

The mean (+SE) values for the materials used in nests constructed by three species of the Sylviidae and one species of the Prunellidae. Values are for the different materials expressed as a proportion of (a) the total nest mass, (b) the cup lining mass, and (c) the outer nest mass.
Grass also featured highly in Sylviidae nests with Eurasian reed warbler nests containing very little else (Table 3, Figure 2). Willow warbler nests contained more moss than grass in the outer nests (Figure 2). Eurasian blackcap nests were loosely woven from grasses and lined with a relatively high proportion of roots. Hair featured in the cups of the willow warbler and Eurasian blackcap but was almost absent in Eurasian reed warbler nests (Figure 2, Table 3).
In contrast to warbler nests dunnock nests had a high proportion of moss and woody stems but relatively little grass (Table 3, Figure 2). Hair and plant fibres were found in the highest proportions in the cup lining compared to the outer nest (Figure 2). Dunnock nests were similar to those of the grey wagtail in that moss was the greatest plant component of the cup lining, rather than grass observed in other species.
The number of materials used in the different parts of the nest were lowest in the woven nests of Eurasian reed warblers but were also low in yellow wagtails and Eurasian blackcap nests (Figure 3). The grey wagtail nests averaged more than six material types but high values were also seen in nests of the willow warbler and dunnock (Figure 3). With the exception of the Eurasian reed warbler, most species had more types of materials in the cup lining than the outer nests (Figure 3). Mean (+SE) number of different material types in different nest parts of eight species of bird. Total nest is indicated by orange bars, the outer nest by black bars, and the cup lining by blue bars. The same material may be present in both the cup lining and the outer nest.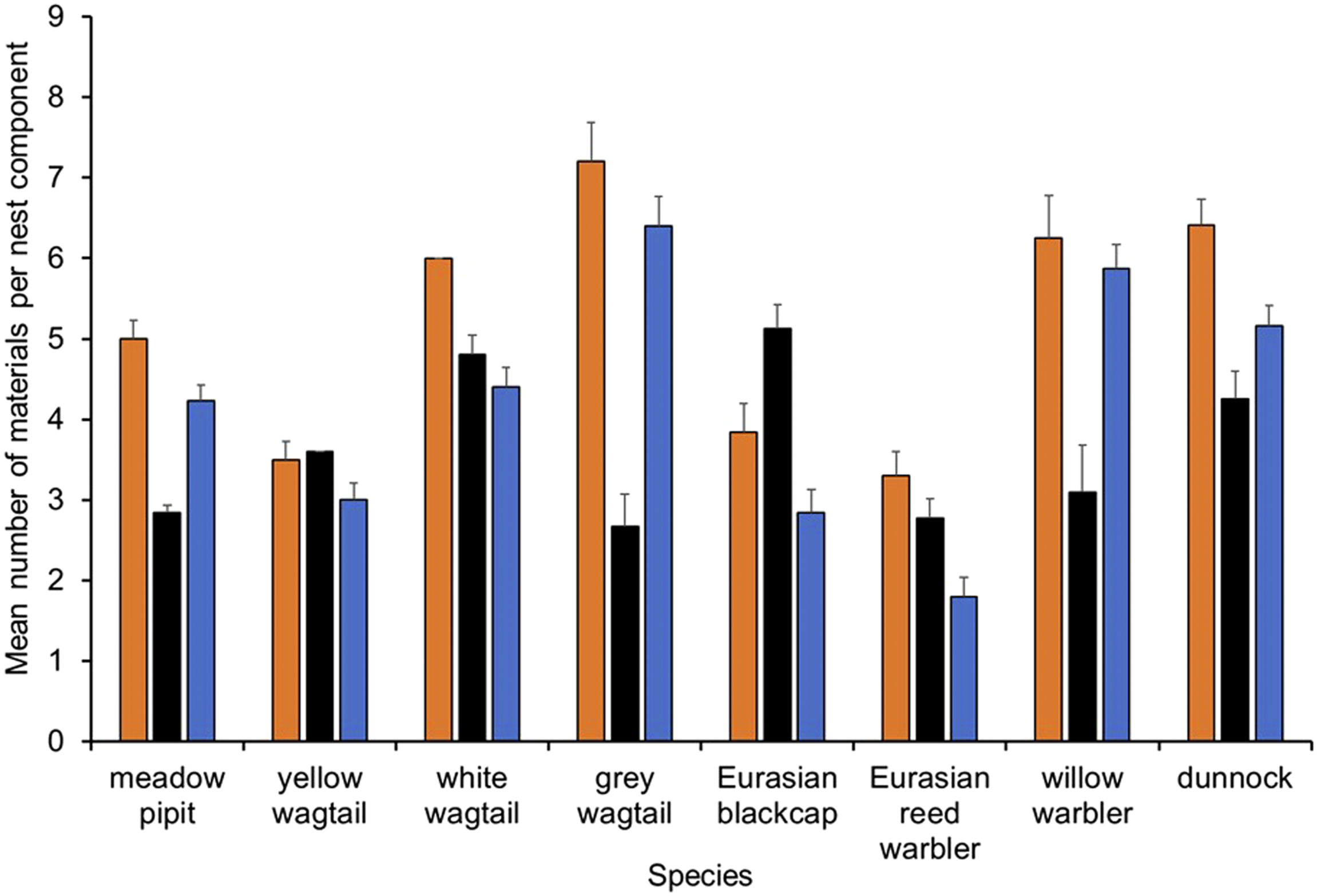
Species comparisons
Phylogenetically-controlled principal component loadings for the composition of total nest, cup lining, and outer materials for nests constructed by pipits, wagtails, warblers and the dunnock. Data include SD, eigenvalue, proportion of variance and cumulative proportion of variance of each. The results of the Levene’s test tested the effect of species on the variance and Kruskal-Wallis tests compared the differences between species for mean PC scores.

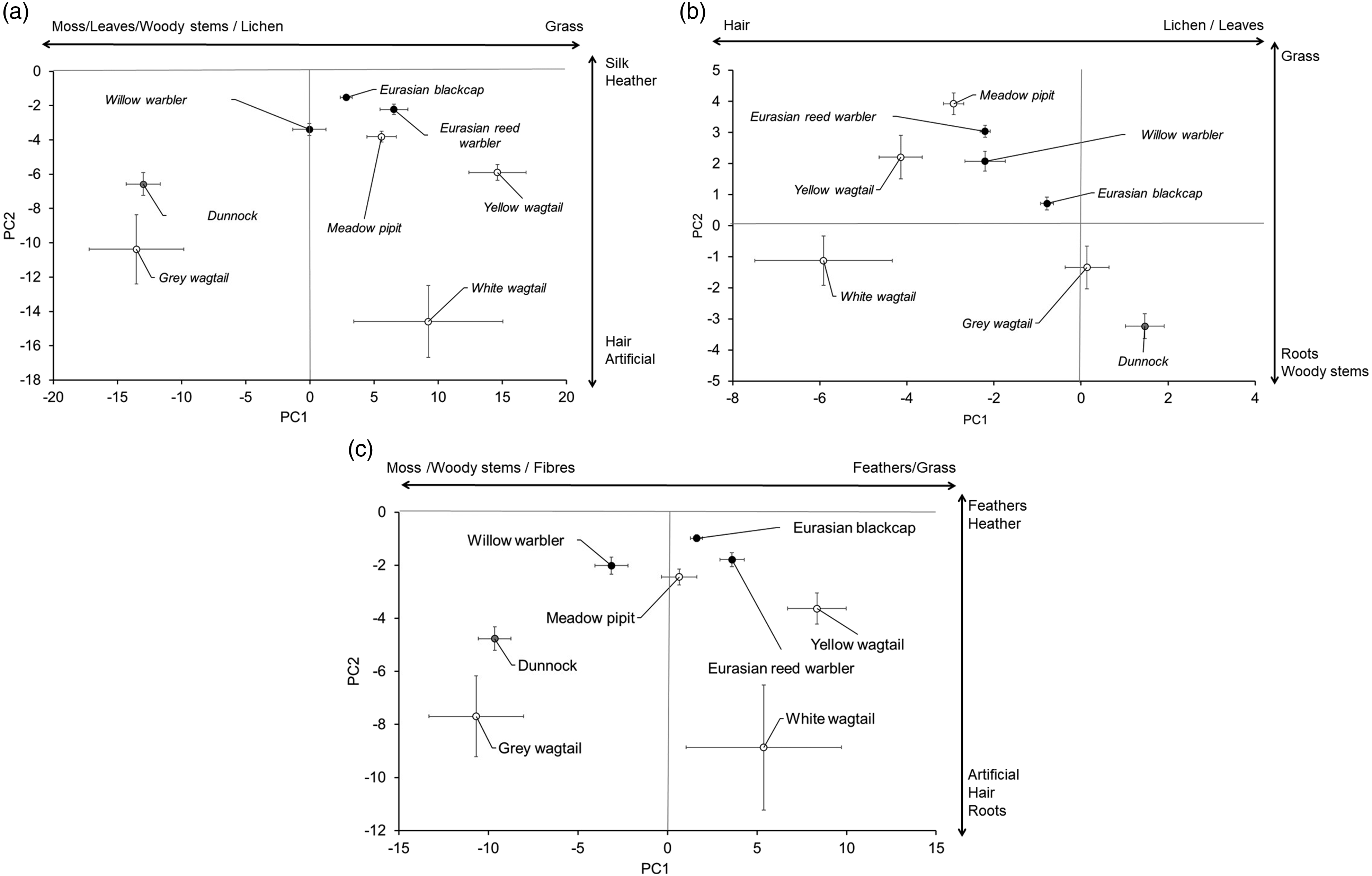
The relationship between the mean (±SE) values for the first two principal components when all components for the total mass (a), cup lining (b) or outer nest (c) were included in the phylogenetically-controlled principal component analysis. The interpretation of the PC scores is as indicated by the arrows parallel to the axis. Open symbols indicate members of the Motacillidae with black symbols indicating members of the Sylviidae and the grey symbol indicating the Prunellidae.
The separation between species for principal component analysis of cup lining data is shown in Figure 4(b). Almost 95% of the variance in the data was explained by the first two PC axes (Table 4). The more negative loadings on PC1 were associated with hair (Figure 4(b), Table 4). For PC2 most species were similar in the loadings except for the white wagtail that had more roots and woody stems (Figure 4(b)). As for the total nest, Levene’s tests and Kruskal-Wallis tests showed significant effects of species on variances and medians, except for the Levene’s test for PC2 (Table 4).
For the outer nest, phylogenetically-controlled principal component analysis explained most of the variation (Table 4) and there was a very similar pattern of species separation to that for the total nest – the warbler species were more clustered than the wagtails (Figure 4(c)). The key differences were for PC2 the species exhibited less variation where more negative values were associated with artificial materials, hair and roots and more positive values were associated with feathers and heather (Figure 4(c)). Again, Levene’s tests and Kruskal-Wallis tests showed significant effects of species on variances and medians (Table 4).
Comparison of cup lining and outer nest
Results of stepwise discriminant analysis comparing the proportion of different animal or plant materials found in the cup lining (C) and the outer nest (N) of species of pipit, wagtail, warbler or the dunnock. An overall Wilk’s lambda value (P-value) comparing the two groups and individual Wilk’s Lambda values indicate whether the nest material contributes significantly in the discrimant function. Only components that were significant for the species indicated are presented.

Discussion
We found that nest size and composition varied among and within species. For instance, despite differences in sample size, Eurasian reed warbler nests were much more similar to each other in terms of size and composition than those of white wagtails. Size of a whole nest was unrelated to average adult body mass in this sample of species. This study used body mass values from Dunning 37 for consistency across species but these values were similar to masses reported by Cramp.45,47 Nests are composed of similar types of materials but in differing quantities. It was possible to distinguish between species in terms of total nest composition, and between the cup lining and the outer nests on the basis of at least one of the component materials.
Intra- and intra-specific variation in composition
Of the species of the Motacillidae considered, dimensions are only previously reported by Cramp 45 for nests of the yellow wagtail and are generally comparable in terms of diameter but the walls and base are much thinner in this study. Grey wagtails nesting in Algeria, built nests that were almost twice the mass of those described here and were mainly constructed from plant material. 46 White wagtails nesting in Finland also built slightly larger nests than reported here when built in natural cavities but when built in domestic attics nest mass was three-times heavier. 47 Data for nest dimensions for the dunnock are generally comparable to published values. 45 Nest dimensions reported for warbler nests here are comparable to those for other Phylloscopus warblers. 48 In Finland, wood warbler (Phylloscopus sibilatrix) and willow warbler nests were lighter than willow warbler nests reported here but these were, in turn, lighter than nests of the chiffchaff (Phylloscopus collybita) in Finland. 49 Willow warbler nests in Estonia were 32% heavier than the average reported here. 50 It is not clear what caused the intraspecific variation in mass between nests from the Baltic and the UK but it may reflect small sample sizes, although previous studies have also suggested that year can also influence nest mass. 35 In Canada, nests of several passerine species were heavier at higher latitudes, 51 so geographical location may play a part. Furthermore, the availability of nesting material may also have an effect on nest mass, as previously seen in great tits (Parus major) in the Mediterranean, 52 although it is not possible to determine due to the materials not being documented in the Baltic studies. There was no significant relationship between nest size and body size, which probably reflects the small range in adult body size in the species involved and mirrors a result described by Biddle et al. 30 for other passerine families.
As in previous studies24,30 inter-specific variation in nest size and composition was high in our study. White and grey wagtails constructed substantial nests that were twice the mass of those built by yellow wagtails despite their similarity in adult body mass. Why this is the case is unclear but may reflect the localised temperature and humidity of the nest sites. At the opposite extreme, as has been reported previously, 53 Eurasian blackcap nests in our study were loosely constructed and gaps were visible within the nest base and walls. Air gaps between nest materials may provide insulation by trapping a layer of air. 54 The lack of variation between the composition of cup and outer lining of Eurasian reed warbler nests is perhaps reflective of the preferred nest materials and the fact that they were all derived from one nesting location. Geographical variation in nest composition is reported in the UK for European pied flycatchers (Ficedula hypoleuca) 33 and common redstarts (Phoenicurus phoenicurus) 55 and for a variety of species in Canada. 51 Further research into geographical variability of nest composition in a wider range of species would be of interest.
The composition of nests is often only described in qualitative terms.34,56,57 Whilst studies exist that describe what materials are in nests, they are often reported in terms of the percentage of the sample of nests that contain that type of material 25 rather than how much of each of the materials was present. Alternatively reports only provide data for a single component of the nest, e.g., artificial materials 58 or aromatic plants. 59 Comprehensive quantitative data for whole nest composition is more useful in terms of understanding nest function, e.g., thermal insulation 9 or response to rainfall. 60 This report has increased the published dataset by six new species bringing the current total to 31 passerine species of mainly European origin.3,16,30 While this is a marked expansion of species coverage since 2010, this is far short of the 5000 passerine species so we remain largely ignorant of the composition of most passerine nests and detailed composition of non-passerine nests has not been reported. This is hampering our understanding of how inter-specific variation in the materials affects the various functions of a nest. For instance, the insulative properties of a nest wall are similar for a wide range of species despite differences in nest composition. 9 In addition, there may be, as yet unexplored, differences between the reasons for the use of materials between open-nesting and cavity-nesting species. 3
A few studies have demonstrated that geographical location can impact upon nest size and composition.23,32,33,51,55 Given that the nests studied here were from a wide range of locations geographical trends may have been masked within our dataset. Unfortunately, the small samples for each species from each site prevented a more rigorous statistical comparison. However, given the paucity of quantitative data for nest composition of the vast majority of passerines, we felt that it was important to report these data to facilitate a better understanding of the variation that exists within birds. Future studies should aim to obtain larger numbers of nests from a wider geographical range of locations, especially outside of Europe, and different habitats, e.g., tropical forests, in order to understand better the effects of geographical variation on composition. 33
Variation in nest morphology
Our study complements the report by Biddle et al. 30 that described similar data and patterns for thrush, old world flycatcher and finch nests. Different regions within avian nests have previously been identified by the types of material present within specific parts of the nest and have been suggested to perform distinct functions.1,7,11,16,30,61 Choice of nest components and specific placement of components in nest regions suggest that the bird may have an awareness of the materials’ properties and are using them in response to the localised physical environmental conditions they experience.11,62,63 The results of our study provide empirical support for the notion that nest materials can be used to create recognisable nest regions 30 but the functional properties of these distinguishing features have yet to be fully explored. Moss is more common in the outer base and sides of nests of many bird species, particularly those breeding in cavities,16,35,32,64 although it is also present in nests built in the open.16,30 Although previously considered to be involved in moisture absorption, 65 moss may provide structural support in the nest wall 64 or may possibly be used in camouflaging the nest. 66 Grass is a common structural element in nests of small passerines, including many species described here, but as adult body masses increase across species there is a shift towards use of more rigid and stronger woody stems.62,67 Materials also have differing insulative properties9,17,19,68 or absorb water, or dry out, at different rates. 60 As we increase our understanding of what nests are constructed of then this will help us to understand the roles that nest components play in overall nest structural integrity in far more detail.
To understand the functional properties of a nest, a complex bio-engineered structure, we need to know what it is it made of. There are 11 more species of warbler, and two more species of pipit, that regularly breed in the UK, 69 and 12 more species worldwide in the Prunellidae, 70 that have yet to be investigated in terms of nest composition. Whilst it may seem that collecting data from additional species is a little unnecessary, such data will be needed to further our understanding of the factors affecting plasticity in nest construction in relation to function.71,72
It is important to note that the nests studied to date, here and in other studies, have been studied ex situ and have been removed from an ecological context. The inter-relationships between nest sites and the size and composition of the nest have yet to be explored. Future studies should start by considering the biotic, e.g., predation, and abiotic, e.g., environmental, factors that are determining the location of a nest site and how these subsequently impact on nest structure and composition. Armed with more information on nest composition and structure 12 it should be possible to understand better nest construction behaviour and make predictions about how birds should construct nests in captive situations where access to nest materials can be controlled. A greater number and diversity of species for which nest composition has been quantified will also allow us to explore how variability in nest composition and structure evolved.
Different materials confer varying levels of insulation to the nest wall 9 and so may reflect a bird’s response to localised weather conditions during construction. 74 Nest construction by the same birds exhibits a high degree of repeatability between years75,76 but longer-term studies have yet to confirm whether nest composition is affected by prevailing weather conditions during nest construction. We have no idea how ongoing climate change, especially increased unpredictability of weather conditions, will impact on avian reproduction. 77 The data presented here contribute to a database that sets a baseline for nest composition, which may prove useful in future studies of how climate change will impact on avian distributions and reproduction. 77
Footnotes
Acknowledgements
We thank the numerous members of the British Trust for Ornithology’s Nest Record Scheme (NRS), and to Professor Nick Davies, who generously supplied the nests used in this study. LB is grateful for the financial support of the School of Life Sciences, University of Lincoln. We are grateful to reviewers of previous iterations of this manuscript, and the journal editor, for constructive comments that helped in its improvement report. Many thanks to Carl Soulsbury for his statistical advice and help.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.