
Abstract
Introduction
Gene therapy has now reached clinical trials for the treatment of human tumors, including those of the prostate, and antisense oligonucleotides (oligos) have targeted bcl-2 and clusterin, in efforts to restore apoptosis following radiotherapy [Mu et al. 2005; Yip et al. 2005] or chemotherapy [Yamanaka et al. 2005]. If such therapy is to be successful, it is important to examine mechanisms by which tumors can evade this therapy through compensation. We recently reported [Rubenstein et al. 2011a] that, in the LNCaP tumor model, inhibition of bcl-2 with antisense oligos suppressed the expression of the apoptotic promoter caspase-3. In another form of compensation, we also found that expression of the androgen receptor (AR) was increased [Rubenstein et al. 2011b]. In this study we evaluated similar effects upon the AR transcriptional coactivator p300, which is usually associated with and more highly expressed in advanced prostate tumors [Bouchai et al. 2011]. Enhanced AR and p300 expression could not only select cells which again evade apoptosis and enhance tumor progression (even with bcl-2 suppression), but could also suggest the emergence of a more aggressive (hormone-insensitive) phenotype. For gene therapy to ultimately be successful it must be made more specific or mechanisms of compensation must be identified and subsequently suppressed.
Effective therapeutics target unique characteristics of etiologic agents including bacterial cell walls and ribosomes or viral encoded proteolytic enzymes. The development of tumor resistance is less specific or distinct since cancer cells are not substantially different from noncancerous (differentiated cells). They use the same biochemical pathways and, unless virally induced most are (even antigenically) similar to normal cells. The effectiveness of chemotherapy capitalizes on the fact that within a tumor mass a greater proportion of cells are in the process of replicating. Therefore, anticancer drugs frequently target some aspect of DNA synthesis. In prostate and breast cancers, growth factor (hormonal or protein) deprivation provide another type of therapy where orchiectomy or biochemical analogs and agonists block or interfere with steroid production or activity. An additional approach could target transcriptional activity initiated by the DNA hormone response elements recognized by the hormonal binding receptors (AR or estrogen receptor [ER]) and their coactivators (p300 and CREB binding protein [CBP]), which are more prevalent in advanced, hormone-insensitive disease [Bouchai et al. 2011]. However, for most chemotherapies, tumor cell specificity is relative, often lacking, and most agents administered (such as paclitaxel) have significant toxicity towards other replicating cells, producing side effects in tissues or organs (gut or bone marrow) with rapid cell replenishment.
Gene therapy is based on a similar premise and while effective protocols can employ either translational suppression (mediated via oligos) or replacement (of inactivated, mutated or deleted suppressor genes such as PTEN) technology [Huang et al. 2001] both tumor and normal cells express the same genes. Targets for gene therapy are found in many pathways and it is likely that hundreds (or thousands) of genes can ultimately become involved in the malignant process. Although tumors can express an overall altered pattern of gene expression the levels of many growth regulatory genes are often similar to those of normal cells. Resistance develops because the biochemical pathways involved are complex and highly regulated by many stimulatory and inhibitory factors which can be altered by therapy, therefore it has been suggested that tumors can alter their dependence upon single influences by relying upon others through compensation [Rubenstein et al. 2011a].
Tumors are essentially heterogeneous masses of rapidly growing and selectively adapted cells whose sole purpose is to survive, replicate and, while doing so, evade therapeutic interventions. The best example is the emergence of hormone-insensitive prostate cancer cells following androgen deprivation therapy, resulting in the increased expression of the autocrine loop consisting of transforming growth factor-α (TGF-α) and its binding site the epidermal growth factor receptor (EGFR) in prostate and breast tumors [Rubenstein et al. 1994].
As bacteria and viruses mutate to evade antibiotic and antiviral agents, tumor cells are under similar selective pressure to evade chemotherapy. Although newly developed forms of gene therapy provide specific ways to inhibit uncontrolled growth or promote (re-establish) apoptosis, the unintended consequences of intervention are poorly understood, and some may compensate for the originally intended effect.
Methods
Oligonucleotides
Oligos (monospecific or bispecific) were purchased from Eurofins MWG Operon (Huntsville, AL). Each was phosphorothioated on three terminal bases at 5’ and 3’ positions. Stock solutions were made to a final concentration of 625 μM in sterile Dulbecco PBS.
Base sequences
Each oligo contained at least one
Cell culture
LNCaP cells were grown in RPMI 1640 supplemented with 10% bovine serum, 1% L-glutamine and 1% penicillin/streptomycin in a 5% CO2 incubator. Log phase cells were harvested using EDTA/trypsin and equally distributed into 75 cm2 flasks (Corning, NY, USA). At intervals media were either supplemented or replaced with fresh.
Oligo treatment prior to PCR
Four days prior to oligo addition, when cell density approached 75% confluence, 10 ml of fresh media was added. Cells were incubated for an additional 3 days before 5 ml of media was replaced with fresh the day before oligos were added. We added 100 μl of stock oligos to bring the final concentration to 6.25 μM. Incubation proceeded for an additional 24 hours in the presence or absence of monospecific MR4, or the MR24 and MR42 bispecifics.
RNA extraction
Following treatment, media was removed, a single milliliter of cold (4°C) RNAzol B was added to each 75 cm2 culture flask and the monolayer lysed by repeated passage through a pipette. All procedures were performed at 4°C. The lysate was removed, placed in a centrifuge tube to which 0.2 ml of chloroform was added, and shaken. The mixture stayed on ice for 5 min, was spun at 12,000g for 15 min, and the upper aqueous volume removed and placed in a fresh tube. An equal volume of isopropanol was added, the tube shaken, and allowed to stay at 4°C for 15 min before similar centrifugation to pellet the RNA. The supernatant was removed, the pellet washed in a single milliliter of 75% ethanol, then spun for 8 min at 7500g. The ethanol was pipetted off and the formed pellet air dried at –20°C.
RNA quantitation
RNA was resuspended in 250 μl of DEPC-treated water, and quantitated using a Qubit fluorometer and Quant-iT RNA assay kit (Invitrogen). DEPC is an inhibitor of RNase activity.
RT-PCR
Extracted RNA was diluted in DEPC-treated water to 40 µg/μl, then 1–4 μl of this RNA was added to 1 μl of both sense and antisense primers (forward and reverse sequences) for bcl-2, AR and p300. From a kit purchased from Invitrogen, the following reactants were added for RT-PCR: 25 μl of 2× reaction mixture, 2 μl SuperScript III RT / platinum Taq mix, tracking dye, and 3 μl of MgSO4 (of a 5 mM stock concentration). DEPC-treated water was added to yield a final volume of 50 μl. RT-PCR was performed for 2 × 25 cycles using the F54 program in a Sprint PCR Thermocycler. As a control for RT-PCR product production, human actin expression was tested in RNA extracted from HeLa cells which was provided in a kit purchased from Invitrogen (in the reaction mixture, no MgSO4 was included, the difference compensated for by 3 μl of DEPC treated water).
Primers
Actin
Forward primer sequence: 5’ CAA ACA TGA TCT GGG TCA TCT TCT C 3’
Reverse primer sequence: 5’ GCT CGT CGT CGA CAA CGG CTC
PCR product produced was 353 base pairs in length
bcl-2
Forward primer sequence: 5’ GAG ACA GCC AGG AGA AAT CA 3’
Reverse primer sequence: 5’ CCT GTG GAT GAC TGA GTA CC 3’
PCR product produced was 127 base pairs in length.
AR
Forward primer sequence: 5’ CGG AAG CTG AAG AAA CTT GG 3’
Reverse primer sequence: 5’ ATG GCT TCC AGG ACA TTC AG 3’
PCR product produced was 155 base pairs in length.
p300
Forward primer sequence: 5’ CGC TTT GTC TAC ACC TGC AA 3’
Reverse primer sequence: 5’ TGC TGG TTG TTG CTC TCA TC 3’
PCR product produced was 167 base pairs in length.
Detection and quantitation of product
Agarose gel electrophoresis
We prepared 1.5% agarose gels in a 50 ml volume of TBE buffer (1× solution: 0.089 M Tris borate and 0.002 M EDTA, pH 8.3), containing 3 μl of ethidium bromide in a Fisher Biotest electrophoresis system. Samples were run for 2 hours at a constant voltage of 70 V using a BioRad 1000/500 power supply source. To locate the amplified PCR product, 3 μl of a molecular marker (Invitrogen) which contained a sequence of bases in 100 base pair increments (Invitrogen) as well as 2 μl of a sucrose-based bromophenol blue tracking dye were run in each gel.
Quantitation
Gels were visualized under UV light and photographed using a Canon 800 digital camera. Photos were converted to black and white format and bands quantitated using Mipav software provided by the National Institutes of Health. Means and standard deviations were compared using Student’s t-tests to determine significance.
Results
bcl-2 expression
As a control (data not shown) for RT-PCR product production, human actin expression was tested in RNA extracted from HeLa cells [Rubenstein and Guinan, 2010a].
LNCaP cells incubated for 24 hours in the presence of 6.25 μM of oligos suppressed bcl-2 expression, and support the finding of comparable biologic activity in both monospecific and bispecific oligos measured in the in vitro cell growth inhibition experiments [Rubenstein and Guinan, 2010a]. When photographs of the identified product bands were scanned on agarose gels and quantitated using Mipav software, in a series of runs, the greatest expression of bcl-2 was always found in untreated LNCaP cells. Those treated with oligos, whether monospecific or bispecific, produced bands which indicated obvious (to the naked eye) suppression. For each oligo evaluated, the greatest amount of suppression measured approached 100% for the monospecific MR4; and for the bispecifics MR24 and MR42, 86% and 100%, respectively. Suppression was found in both repeat PCR runs with bcl-2 primers, as well as in repetitive agarose gel quantifications. Figure 1 presents a bcl-2 product band in the expected 127 base pair region which in this run was inhibited 23% by treatment with the monospecific MR4, and 86% and 74%, respectively, by bispecifics MR24 and MR42, as measured by Mipav software.

bcl-2 expression is suppressed by oligos as indicated in a representative agarose gel.
AR expression
Comparable amounts of extracted RNA from LNCaP cells treated with either monospecific or bispecific oligos directed against bcl-2 (and EGFR in the bispecifics) were then evaluated by RT-PCR using primers directed against AR. When background intensity was subtracted, the relative intensity of all bands corresponding to AR representing cells treated with MR4, MR24 and MR42 compared with controls were enhanced 31.2% ± 26.0% (p = 0.015), 58.5% ± 51.4% (p = 0.019) and 53.1% ± 45.9% (p = 0.019). These results were pooled from both duplicate PCR runs and multiple gels (a total of six gels were evaluated), and indicate similar (significant) enhancement of AR activity is produced by each oligo type. A representative band is depicted in Figure 2.

Androgen receptor expression is enhanced by oligos as indicated in a representative agarose gel.
p300 expression
Comparable amounts of extracted RNA from LNCaP cells treated with either monospecific or bispecific oligos directed against bcl-2 (and EGFR in the bispecifics) were then evaluated by RT-PCR using primers directed against p300. When background intensity was subtracted, the relative intensity of all bands corresponding to p300 representing cells treated with MR4, MR24 and MR42 compared with controls were increased 82.9% ± 51.9% (p = 0.006), 93.0% ± 87.3% (p = 0.044) and 105.4% ± 65.9% (p = 0.007). These results were pooled from both duplicate PCR runs and multiple gels (a total of six gels were evaluated), and indicate similar (significant) enhancement of p300 activity is produced by each oligo type. A representative band is depicted in Figure 3.
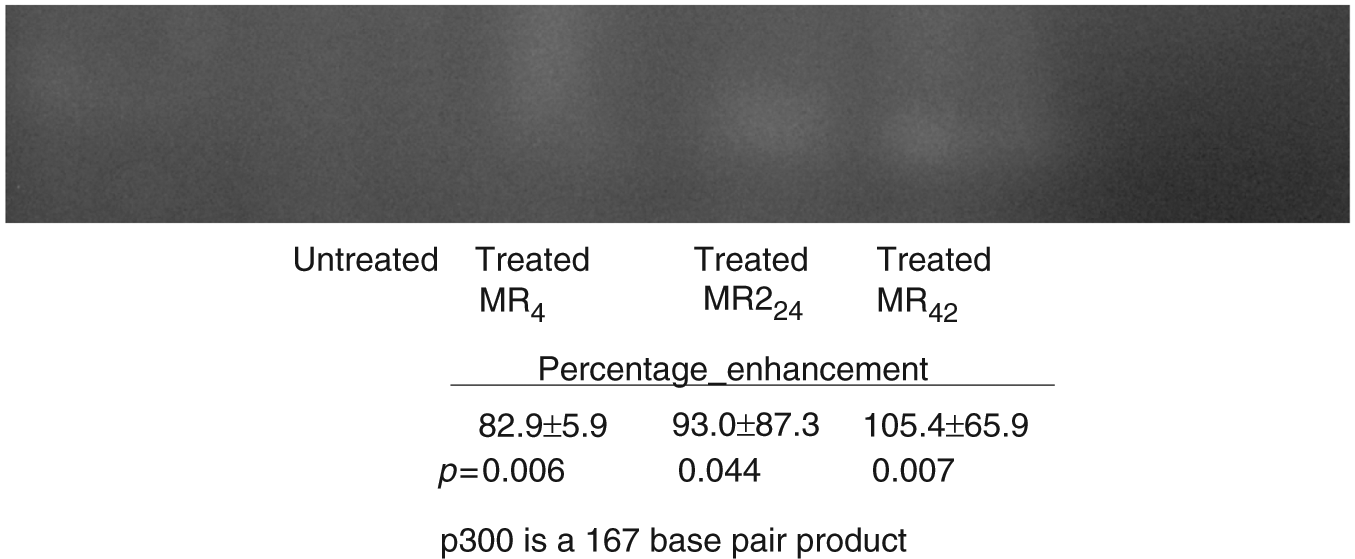
p300 expression is enhanced by oligos as indicated in a representative agarose gel.
Discussion
The AR (also known as NR3C4; nuclear receptor subfamily 3, member 4) plays a principle role in male sexual development, prostate function, cancer progression and various treatment strategies. Following the cytoplasmic binding of the AR to testosterone or its metabolite, dihydrotestosterone (DHT), it undergoes a conformational change which is accompanied by dissociation of heat shock proteins and translocation into the cell nucleus. The AR dimerizes, binds to hormone response elements of the DNA, and acts as a transcription factor to enhance the synthesis of growth stimulating proteins, including insulin-like growth factor (ILGF) [Pandini et al. 2005]. In addition to ILGF, other growth factors such as TGF-α acting through their respective receptors (such as the EGFR which binds TGF-α) contribute to unregulated prostate cancer growth. These protein factors and their receptors are considered targets for suppressive gene therapy with antisense oligos [Rubenstein et al. 2004]. Although disruption of this process by androgen deprivation provides the rationale for most types of prostate cancer treatment, most tumors recur in an androgen-insensitive form within a few years. At this stage, genes are driven towards transcription by both AR and coactivating transcription factors p300 and its homolog CBP. p300 is essential for cell growth and governs the expression of the cyclins regulating the transition between G1, S, G2 and M phases of mitosis [Heemers et al. 2007]. Acting with IL-6, p300/CBP plays a role in the androgen independent expression of prostate specific antigen (PSA) [Debes et al. 2005]. In the LNCaP model, administration of R1881 reduces both CBP mRNA and the encoded CBP protein suggesting, that following androgen ablation, the expression of some coactivators increase and contribute to a state of AR hypersensitivity [Klocker et al. 2007]. Treatment of prostate cancer cells with siRNA directed against p300 reduces cancer cell growth [Klocker et al. 2007], and eliminates the ability of IL-6 to induce PSA [Klocker et al. 2007]. Since both transcriptional coactivator proteins p300 and CBP are expressed to a greater extent in advanced prostate cancer [Heemers et al. 2007], well-differentiated, androgen-sensitive LNCaP cells would be expected to have relatively low expression of p300, and this is evident in Figure 3 where in the untreated group p300 expression is barely detectable. The enhanced expression seen following oligo treatment makes its induction appear more impressive, and might indicate a possible transition to a pattern of gene expression more associated with later stage (androgen insensitive) disease. This suggests that oligo treatment directed against bcl-2 not only can be evaded through compensatory changes in expression which encourage tumor growth, but could also contribute to further dedifferentiation and hormone insensitivity.
Innovative protocols to disrupt androgen driven tumor progression have employed antisense oligos directed against the enzyme for conversion of testosterone to dihydrotestosterone (5-alpha reductase), heat shock proteins, p300 and the AR itself. LNCaP cells express an AR which is mutated in the binding domain however the groups of Eder et al. and Rubenstein et al. have separately demonstrated growth inhibition in this in vitro model employing oligos [Eder et al. 2000; Rubenstein et al. 2010].
Gene therapy is a complex process requiring multiple pathways (and the regulatory proteins) to be simultaneously regulated. In addition, the ‘driver genes’ (usually kinases) which greatly influence tumor growth must be distinguished and targeted rather than similarly mutated ‘passenger genes’.
Oligos (produced by Oncogenex Pharmaceuticals) have reached clinical trials for the treatment of prostate cancer (OGX-011), while others remain in preclinical development (OGX-225). Often administered in combination with traditional chemotherapy, these oligos target bcl-2, clusterin (OGX-011 in phase II testing), heat shock protein 27 (OGX-427) or insulin growth factor binding proteins (OGX-225) [Oncogenex, 2011]. Many represent efforts to restore tumor apoptosis by eliminating suppressive bcl-2 [Mu et al. 2005; Yamanaka et al. 2005; Yip et al. 2005] associated with treatment resistance. Similar approaches are directed at clusterin. For (tumor suppressor) genes which are either diminished or lacking in expression gene transfection has been attempted in prostate cells which contain a mutated PTEN [Huang et al. 2001]. Although antisense oligos are specifically directed through complementary base pairing to inhibit mRNA translation of genes, there can be nonspecific effects on nontargeted genes following oligo-mediated bcl-2 suppression. The effectiveness of bcl-2 and overall apoptosis activity is highly regulated and dependent upon the expression of many stimulatory, inhibitory, stabilizing factors, as well as the ratio between these proteins. As demonstrated, the specific suppression of one apoptosis inhibitory protein (bcl-2) is nonspecifically compensated for by the suppression of a nontargeted promoter, caspase-3 [Rubenstein et al. 2011a]. Clinically these types of experiments are important because they suggest that for oligo-mediated bcl-2 suppression to be effective caspase-3 activity should be either maintained or enhanced [Rubenstein et al. 2011a]. Other compensatory alterations have been evaluated (Table 1), and include additional regulators of apoptosis as well as surface antigen and cytokine expression. Additional coactivators of the AR (IL-4) and autocrine loops involving TGF-α, its binding site the EGFR and insulin-like growth factor (IGF1) should also be evaluated. In these experiments bispecific oligos also targeted the EGFR. However, in previous experiments we found that growth inhibition produced by monospecific oligos directed against EGFR was not accompanied by decreased mRNA expression [Rubenstein et al. 2002]. Currently Genta is conducting a phase III test using oligos (Genasense; oblimersen) directed against bcl-2 for treating melanoma, chronic lymphocytic leukemia and various solid tumors [Genta, 2011], but compensatory effects produced by this agent have not (yet) been reported. Tumors are resilient in their efforts to overcome (even newly developed) therapeutics and become resistant. If gene therapy is to be effective, we must understand how primary effects evoke compensatory changes. It would also be significant to see whether these changes are replicated in an in vivo model. If these lead to enhanced expression of undesired proteins, the oligo approach can again be applied. If antisense technology is to be fully exploited multivalent forms (as proposed) can be developed to suppress multiple proteins [Rubenstein et al. 2006].
Summary of gene expression results directed against bcl-2.

This year (2011) the American Cancer Society (ACS) estimates that in spite of early detection, screening for PSA and effective treatments for localized disease, in the United States there will be 33,720 deaths from prostate cancer with 240,890 newly diagnosed cases [Seigel et al. 2011]. New types of treatment, including gene therapy and translational inhibition must be developed and employed (probably in combination with traditional androgen ablation).
Footnotes
The Cellular Biology laboratory at the Hektoen Institute is supported, in part, by the Blum Kovler Foundation, the Cancer Federation, Safeway/Dominicks Campaign for Breast Cancer Awareness, Lawn Manor Beth Jacob Hebrew Congregation, the Max Goldenberg Foundation, the Sternfeld Family Foundation, and the Herbert C. Wenske Foundation.
The authors declare no conflicts of interest in preparing this article.