
Abstract
Upon ischaemic stroke, brain-resident and peripheral immune cells accumulate in the central nervous system (CNS). Interestingly, these cells express pattern specific to neurotransmitter receptors and, therefore, seem to be susceptible to neurotransmitter stimulation, potentially modulating their properties and functions. One of the principal neurotransmitters in the CNS, dopamine, is involved in the regulation of processes of brain development, motor control and higher brain functions. It is constantly released in the brain and there is experimental and clinical evidence that dopaminergic signalling is involved in recovery of lost neurological function after stroke. Independent studies have revealed specific but different patterns of dopamine receptor subtypes on different populations of immune cells. Those patterns are dependent on the activation status of cells. Generally, exposure to dopamine or dopamine receptor agonists decreases detrimental actions of immune cells. In contrast, a reduction of dopaminergic inputs perpetuates a pro-inflammatory state associated with increased release of pro-inflammatory molecules. In addition, subsets of immune cells have been identified to synthesize and release dopamine, suggesting autoregulatory mechanisms. Evidence supports that inflammatory processes activated following ischaemic stroke are modulated by dopaminergic signalling.
Keywords
Introduction
Ischaemic stroke represents the endpoint of pathological cascades as a consequence of multiple determinants such as risk factors and comorbidities. Unhealthy lifestyle habits significantly contribute as a risk factor for stroke and associated comorbidities. In addition, the risk of suffering ischaemic stroke increases with age and changes of sex hormones; rarely, genetic predispositions contribute to pathological alterations up to occlusion of brain arteries. Adaptive processes to compensate for reduced blood flow to the brain have been identified. Hence, a sudden drop in blood perfusion for a critical period of time causes acute cell death involving well-characterized mechanisms, 1 with the release of molecules from dysfunctional and disintegrating cells. These so-called damage-associated molecular patterns contribute to initiation of an inflammatory response, including the activation of brain-resident and blood-borne immune cells. 2
Monoamine signalling is of fundamental importance for almost all brain functions, including motor control, learning and reward-motivated behaviour. In particular, the role of dopamine in physiological processes and neurodegeneration has been extensively studied during the last decades. 3 Effects of the neurotransmitter are mediated by activation of dopamine receptors (DRs), a class of G protein-coupled receptor molecules containing five different subtypes (D1R to D5R). There is an increasing number of studies showing that the neurotransmitter does not exclusively affect neurotransmission, but also modulates functions of non-neuronal cells in the central nervous system (CNS). Interestingly, independent studies have shown the expression of different types of DR on astroglial cells4–6 and microglia upon injury.5–7 The expression of D2R has been found on oligodendrocytes in the developing brain. 8 The expression of functional DR in astrocytes and treatment with levodopa is associated with increased levels of growth factors – that is, glial cell-line-derived growth factor in the ischaemic territory of rats subjected to transient middle cerebral artery occlusion (tMCAO). 9 The role of dopamine signalling in immune cells has been investigated by a number of studies. This article provides an overview on expression profiles of DRs on different types of immune cells (Figure 1) and effects of dopaminergic treatments on immune cells, with an emphasis on poststroke inflammation following experimental stroke.
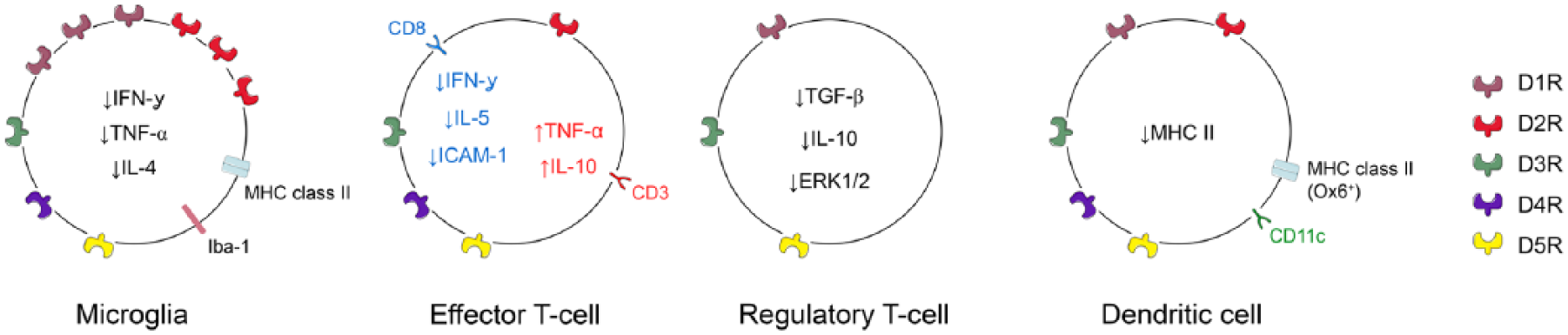
Expression of different dopamine receptors on microglial cells, effector and regulator T cells and dendritic cells, respectively.
Expression and function of dopamine receptors on immune cells
Microglia/macrophages
Microglia represent the principal brain-resident immune cells immediately responding to changes in tissue homeostasis. Upon focal ischaemia, microglia accumulate in the lesioned hemisphere and show different morphologies, dependent on localization and time point after stroke onset. Many studies have associated particular functions of microglia with different morphologies and associated the expression of certain protein markers with cellular functions. 10 Importantly, based on morphology, brain-resident microglial cells are not distinguishable from invading monocytes (macrophages) due to overlap of marker protein expression in both cell populations in situ. Microglia/macrophages represent the largest population of immune cells expressing major histocompatibility complex (MHC) class II molecules in the post-ischaemic brain during the first weeks following tMCAO. Upon injury, the expression mainly of D1R and D2R receptors is upregulated on MHC class II+ and ionized calcium-binding adapter molecule 1 (Iba1) positive microglia in the lesioned hemisphere (Figure 1).6,7 Treatment with levodopa/benserazide downregulates levels of MHC class II proteins in the peri-infarct area without affecting the number of MHC class II+cells. 6 Interestingly, levodopa treatment increased the number of MHC class II+ cells in the corpus callosum contralateral to the lesioned hemisphere. In addition, treatment with levodopa decreased the level of the T cell-associated pro-inflammatory cytokines interferon gamma (IFN-γ), tumour necrosis factor alpha (TNF-α) and interleukin (IL)-4 in the ischaemic territory, without effects on the number of immune cells accumulating in the lesioned hemisphere. 5 All changes are in contrast to the non-lesioned brain, where microglial cells essentially lack DR. 7
In vitro, subpopulations of microglia are immunoreactive for all five DR (D1R to D5R), dependent on the species from which cells have been derived.7,11,12 Application of dopamine resulted in changes of membrane currents and inhibition of inward currents. Moreover, dopamine significantly reduced the release of nitrite in lipopolysaccharide (LPS) stimulated microglia and increased microglial migratory activity.11,12 Susceptibility and response to neurotransmitters and hormones could be modulated by stimulation with LPS, IFN-γ and IL-4. 13
Primary macrophages express all DR on the mRNA level; in addition, all receptors except D5 are translated into proteins. 14 After tMCAO, bone marrow-derived invading macrophages (BMDMs) express D2R in the post-ischaemic brain. 7 Moreover, macrophages express tyrosine hydroxylase (TH) and aromatic amino acid decarboxylase involved in dopamine synthesis, as well as dopamine transporter and vesicular monoamine transporter 2, which are both involved in dopamine and catecholamine transport, respectively. 14 Upon activation, but not in naïve cells, catecholamine synthesis has been observed in RAW264.7 cells, a macrophage cell line.15,16 BMDMs have been shown to express D1R and produce endogenous dopamine in vitro. 17 Activated macrophages are susceptible to catecholamines, and stimulation with low concentrations of dopamine increased IFN-γ-induced phagocytic activity while high concentrations inhibited phagocytosis.18,19 Dopamine also facilitated the clearance of IgG-sensitive erythrocytes by macrophage Fc-gamma receptors in vivo, mediated by both D1R and D2R. 20 These studies consistently show that microglia/macrophages have an autochthone production and release of dopamine and express functional DR; however, both are dependent on activation of cells by diverse stimuli.
The response of macrophages to dopaminergic treatment depends on their activation status and expression of DR. Direct stimulation of naïve macrophages with dopamine increased the release of IL-6 and chemokine (C-C motif) ligand 2. LPS-stimulated cells also showed increased levels of chemokine (C-X-C motif) ligand 8 and IL-10, whereas the level of pro-inflammatory TNF-α was decreased. 14 Interestingly, risperidone, an atypical antipsychotic acting as a DR antagonist (D1R, D2R, D4R, D5R), and clozapine reduced the release IL-12 p40. In addition, risperidone decreased the production of nitric oxide and enhanced the level of IL-10 by LPS-stimulated BMDMs. Interestingly, CD4 T cells modulating effects have been observed in risperidone-treated macrophages, decreasing the level of IL-17a, IL-2 and IL-4. 17 In contrast, Haskó and colleagues demonstrated that dopamine treatment of LPS-stimulated J774.1 cells and mouse peritoneal macrophages suppressed the production of IL-12 p40 21 via a beta-adrenoceptor-mediated mechanism, since DR antagonists were unable to reverse the dopamine-induced effects. In contrast, application of the beta-adrenoceptor antagonist propranolol completely prevented the inhibitory effect of dopamine on IL-12 p40 production. The authors also reported increased production of the anti-inflammatory cytokine IL-10 by macrophages upon dopamine treatment, both in an adrenoceptor-dependent and -independent mechanisms. The D1-agonist SKF 38393 abolished the chlorpromazine (a reversible blocker of D1R and D2R) amplified staphylococcal enterotoxin B-mediated release of IL-10 from activated macrophages. 22 Summarizing findings from these studies, it becomes evident that, dependent on the type of macrophage and the activation status, both agonists and antagonists on D1R and D2R result in the same biological effects.
Peripheral blood lymphocytes
Expression of DR in peripheral blood lymphocytes (PBLs) has been studied by independent research groups using different methodologies – that is, radioligand binding studies, reverse transcription polymerase chain reaction (rt-PCR) and flow cytometry. Results show no expression of D1R on PBL, whereas all studies reported an expression of the D5R subtype.23–25 However, the expression of D2R to D5R showed high interindividual differences due to low sample sizes and different detection methods.24–26 Specifically, D2R has been found in three, D3R in three, D4R in four and D5R in five samples out of 19 individuals. 24 The following paragraphs further elaborate on subsets of PBLs, namely T cells, natural killer (NK) cells and B cells.
T cell populations
The dynamics and magnitude of appearance of T cell populations in the post-ischaemic brain have been investigated by several independent research groups. There is a well-orchestrated pattern of accumulation of different T cell populations in the post-ischaemic brain.27,28 All five types of DR are expressed on T cells; however, distribution of subtypes varies between subpopulations, as is discussed below. Interestingly, stimulation with dopamine showed increased expression of TNF-α and IL-10 in T cells. Increased expression of TNF-α was observed 24 h after stimulation and mediated by activation of D3R and D1/D5R, while the expression of IL-10 was upregulated at 72 h following stimulation with specific agonists at D2R and D1/D5R. 29 Interestingly, treatment with D2R and D3R agonists only exerted effects on activated, differentiated T cells, and did not affect the function of resting T cells or quiescent differentiated T cells after antigen exposure. 30
In human resting CD3+ T cells, dopamine activated D2R and D3R resulting in an increase of beta1 integrin-mediated adhesion to fibronectin, a major extracellular matrix component. From this study, authors have suggested that dopamine might play a role in integrin-mediated cellular trafficking and extravasation of T cells into the CNS. 31 Moreover, co-stimulation with dopamine dose-dependently not only inhibited concanavalin A (ConA)-mediated proliferation and differentiation, but also synthesis of IFN-γ in lymphocytes.32,33
In addition, physiological levels of dopamine inhibit the release of cytokines, namely IL-2, IFN-γ and IL-4, and expression of lymphocyte-specific protein tyrosine kinase, Lck and Fyn, in T cells activated by an anti-CD3 antibody. Effects were mediated via D2/3R. Neither antagonists on the D1R, D5R or D4R affected cytokine release or the expression of Lck and Fyn. 34 Bromocriptine, a D2R agonist, has been shown to inhibit proliferation of T cells activated either by ConA or phytohaemagglutinin-P through inhibition of IL-2 production in CD25+ T cells. 35 Interestingly, DR agonists like the D3R agonist quinpirole were only functional on activated T cells. 30 Likewise, stimulation of all types of DR on anti-CD3/28 activated human T cells inhibited proliferation of cells and induced cell quiescence. No effect was observed on resting T cells. Mechanisms through activation of D4R partially involve an upregulation of kruppel-like factor-2 through an inhibition of extracellular signal-regulated kinases (ERKs) 1/2 phosphorylation. 36
CD4+ cells
CD4+ T cells express all subtypes of DR with a preponderance of D1-like receptors compared to D2-like receptors. However, high interindividual differences in distribution of receptors have been found on CD3+/CD4+ T cells: 2.5–22.8% D1R; 2.0–20.9% D5R; 1.1–7.9% D2R; 1.9–15.0% D3R; and 0.8–17.0% D4R, respectively. 37 While naïve CD4+ T cells (CD3+CD4+CD45RA+CCR7+), differentiated cells without selection process, showed a higher expression of D1R, the expression of D2R was higher in central memory cells (CD3+CD4+CD45RA–CCR7+) and effector memory cells (CD3+CD4+CD45RA–CCR7–). Different expression patterns were found in cultivated CD4+ T cells with reduced expression of D5R and increased expression of D2R. 37 Viable CD4+ T cells activated by anti-CD3/anti-CD28 showed an increased expression of all DRs. Interestingly, apoptotic cells showed a higher DR frequency. In addition, it has been demonstrated that D3R-activated CD4+ T cells shift to a TH1-like phenotype since the cytokine expression changes from IL-4 and IL-10 to IFN-γ. 30
T memory cells
These subsets of T cells (CD3+CD4+CD45RA–CCR7– and CD3+CD4+CD45RA–CCR7+) express all DR subtypes. Higher levels of D2-like receptors have been found in central and effector memory cells, which represents an inverted ratio compared to naïve T cells. 37 The expression of DR has not been studied in γδ T cells.
CD8+ T cells
Although not exclusively, D3R seems the most abundant DR on resting CD8+ T cells. Dopamine induces chemotaxis in CD45RA+ naïve CD8+ T cells, but not in CD45RO+ memory/effector CD8+ T cells, via D3R, suggesting that endogenous dopamine is involved in homing of naïve CD8+ T cells. D3R was found to be downregulated once cells are in an activated state. 38 Increased migratory effects induced by dopamine together with inactivation were observed in naïve CD8+ T lymphocytes activated by CD3/CD28 cross-linking. Inactivation was shown by a downregulation of IL-2 expression via ERK1/2 and NF-kappaB inhibition. No effect of dopamine was seen on the release of cytotoxic granules from activated cells in response to CD3 cross-linking.39,40
Following stroke, CD8+ cytotoxic T cells accumulate mainly in the hemisphere ipsilateral to the lesion.5,27,41,42 Recruitment of CD8+ T cells involves the activation of very late antigen-4 (VLA-4) and vascular cell adhesion molecule-1 (VCAM-1). Inhibition of VLA-4- and VCAM-1-initiated T cell entry mechanisms reduces the number of cytotoxic T cells in the post-ischaemic brain, associated with enhanced recovery of lost functions. 43 Treatment with levodopa significantly reduces the number of CD8+ T cells in the lesioned hemisphere following tMCAO. Accompanied T cell-associated cytokines including IFN-γ and IL-5, as well as intercellular adhesion molecule 1 (ICAM-1), were downregulated in the peri-infarct brain tissue. 5 This is in contrast to in vitro investigations on isolated peripheral CD8+ T cells harvested either from levodopa-treated rats or human blood and treated directly with quinpirole. Here, dopaminergic treatment showed elevated levels of IFN-γ mediated via D3R. 30 Further in-depth studies are required to elucidate the role of dopaminergic signalling in different pathological conditions. Hence, treatment with levodopa enhances recovery of lost function after stroke.
Regulatory T cells
Regulatory CD4+CD25+ T cells (Tregs) are involved in a variety of poststroke inflammatory mechanisms that contribute to stroke outcome. We are only beginning to understand the underlying mechanisms in detail. Partially divergent results in terms of beneficial or detrimental actions of these cells most likely relate to different functions of these cells in different spatiotemporal contexts after stroke. 44
Interestingly, Tregs express TH, and besides the other catecholamines, norepinephrine and epinephrine, release dopamine upon stimulation with reserpine. Thus, release of dopamine may downregulate Tregs, interpreted as an autoregulatory mechanism.29,31,45 Further studies will also elicit distribution and functionality of different DR subtypes on these cells. Lymph node- and spleen-derived primary Tregs from dopamine-treated mice predominantly express D1R and D5R. 46 In contrast, human Tregs and CD4+CD25− effector T cells obtained from blood express on the cell membrane both D1-like and D2-like dopaminergic receptors to a similar extent (12–29% of the cells). In addition, human Tregs do not express D1R subtypes. Current knowledge on functionality of DR in Tregs indicates that dopamine-related effects are mediated via D1-like receptors coupled to cyclic adenosine monophosphate (cAMP)-mediated intracellular cascades, once pharmacological blockade of D1-like receptors blocked D1-like receptor-mediated inhibition of Tregs. Dopamine-mediated inhibition of Tregs involves inhibition of the ERK1/2 pathway 46 and dopaminergic cascades are involved in normalization of IL-10 and transforming growth factor beta levels in reserpine-treated Tregs. Inhibitory effects of reserpine treatment on Tregs increased proliferation of effector T lymphocytes, without influencing the production of TNF-α or IFN-γ. 45
Natural killer cells
NK cells represent a population of cytotoxic lymphocytes with functions to be considered similar to cytotoxic T cells. In addition to the MHC-driven response of NK cells, cells show the property of recognizing stressed cells in the absence of antibodies and MHC. After stroke, cells in the ischaemic territory may release chemoattractant molecules – that is, endangered neurons release fraktalkine 47 and thereby recruit NK cells, exacerbating the local inflammatory response and subsequently brain infarction. 48 In a flow cytometry study, all DRs have been found on NK cells except D1R. 24 In NK cells isolated from mouse spleens, all five DRs have been detected and the effects of dopaminergic drugs affecting NK cell toxicity were modulated via the cAMP–PKA–CREB signalling cascade. 49 In particular, activation of D1-like receptors (including D1R and D5R) with the D1R agonist SKF38393 enhanced NK cell-mediated cytotoxicity associated with elevated levels of D1R and D5R, as well as cAMP content and phosphorylated cAMP-response element-binding (CREB) in NK cells, while activation of D2-like receptors (including D2R, D3R and D4R) with the agonist quinpirole attenuated NK cells associated with the reduced D3R and D4R expression and decreased levels of the abovementioned mediators. In contrast, upregulation of D5R inhibited cell proliferation and the production of IFN-γ in activated NK cells. No effects of dopaminergic drugs were found on resting NK cells in this study. 50
B cells
Similar to other immune cell populations, B cells infiltrate the post-ischaemic brain. 27 In mice, the highest number of B cells has been found on day 3 following stroke. Discrepant results have been obtained in regard to the role of circulating B cells in the acute phase following stroke. Depletion and reconstitution of cells, did not affect infarct volume and functional outcome determined on days 1 and 3 after the insult. 51 On the other hand, a previous investigation showed beneficial effects of IL-10-secreting B cell populations in terms of poststroke inflammation and functional outcome 48 h after tMCAO in mice.52,53 Hence, beneficial effects of regulatory B cells might have been solely attributed to release of IL-10. 54 This, however, does not exclude long-term effects of B cells on local inflammatory cascades, mechanisms of gliosis or neuronal plasticity. 55
All types of DR have been identified on circulating human B cells, hence with high variability in expression. 24 Naïve CD19+ B cells express all DRs except D1R. Resting B cells faintly express D4R, which is downregulated upon activation with pokeweed mitogen (PWM), a mitogen for B cells.24,38 Apparently, high concentrations of dopamine increase apoptosis and apoptotic proteins such as Bcl-2/Bax and Fas/FasL. 32 B cells also contain TH mRNA and produce TH, which is upregulated in stress conditions or stimulation with mitogens.56,57 Increased levels of TH may lead to the production and storage of intracellular dopamine and other catecholamines by a protein kinase C-dependent mechanism. 57 Repeated stress is also accompanied by elevation of apoptotic cells and markers Bcl-2/Bax, and reduction of IFN-γ mRNA, IL-2 and IL-4 in B cells. 56 Further studies are needed to understand dopaminergic-mediated mechanisms involved in B cells response after ischaemic stroke.
Dendritic cells
Accumulation of dendritic cells (DCs) in the post-ischaemic brain during the first weeks following the insult has been described.58,59 DCs are professional antigen presenting cells and typically express the surface antigen CD11c and MHC class II (Ox6+) molecules. In addition, subpopulations of DCs with different surface marker profiles have been identified in the post-ischaemic brain (reviewed by Ludewig and colleagues 60 ). Interestingly, accumulating MHC class II+ cells are not only found in the ischaemic territory. Long-term analysis for up to 9 weeks after tMCAO showed an accumulation of stellate MHC class II+ cells in white matter tracts of the ischaemic and contralateral hemipshere. 6 These cells express D1R and treatment with levodopa reduced the levels of MHC class II without affecting the total number of cells in the ischaemic hemisphere. In contrast, treatment increased the number of cells in the corpus callosum of the contralateral hemisphere, suggesting multiple roles of dopamine signalling on antigen presenting cells dependent on the local tissue environment. 6 Additional studies also found other subpopulations of DR on dendritic cells (Table 1). 61 How dopaminergic signalling further affects the function of DCs in the post-ischaemic brain remains to be investigated. As shown before in an LPS exposure model, dopaminergic signalling mediated by D5R modulates CD4+ T cell activation and differentiation into T helper 17 cells. 61
Expression of dopamine receptors on different immune cell populations.
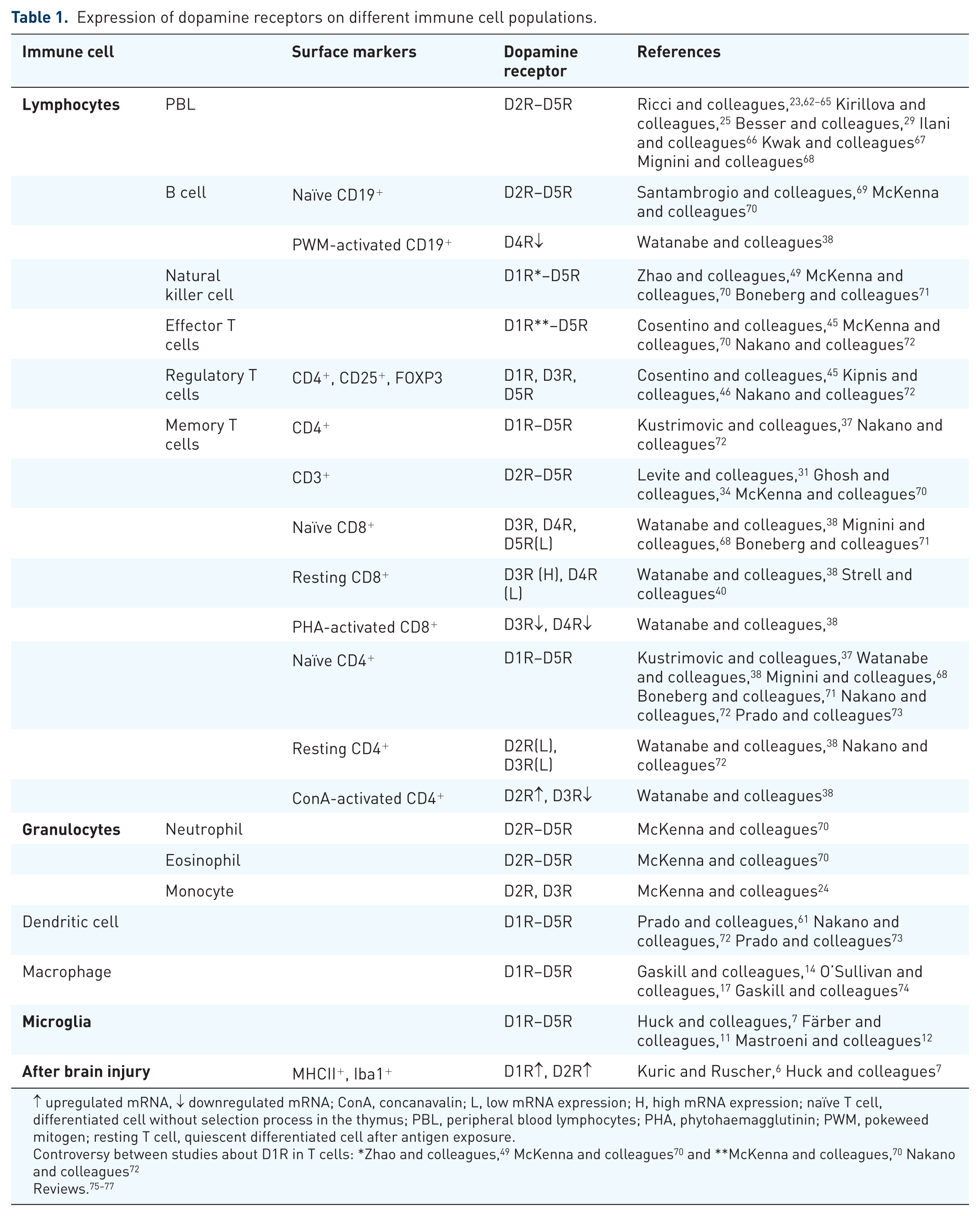
↑ upregulated mRNA, ↓ downregulated mRNA; ConA, concanavalin; L, low mRNA expression; H, high mRNA expression; naïve T cell, differentiated cell without selection process in the thymus; PBL, peripheral blood lymphocytes; PHA, phytohaemagglutinin; PWM, pokeweed mitogen; resting T cell, quiescent differentiated cell after antigen exposure.
Catecholamine synthesis in immune cells
As already mentioned above, immune cells synthesize catecholamines; in particular, dopamine has been found in human cerebrospinal fluid (CSF), CD4+ T cells and B cell extracts obtained from prostate cancer tissues by capillary electrophoresis. Concentrations of dopamine found in CD4+ T cells were approximately 3 × 10–17 mol/cell, in CSF lymphocytes 2 × 10–18 mol/cell and B cells 3.1 × 10–19 mol/cell, compared to striatal levels of 10–20 × 10–9 mol/L. Although dopamine levels seem low, one should not be misled towards speculations that local concentrations are too low to affect functions of cells. The idea that autochthone dopamine synthesis is involved in autoregulatory mechanisms sounds plausible, but warrants further investigation.
Dopamine signalling and poststroke immunodepression
Initial experiments demonstrating increased degradation of genomic DNA in thymus from mice subjected to tMCAO (Ruscher, unpublished observations) provided the basis for subsequent studies identifying a transient phase of immunodepression associated with a higher susceptibility of mice to different types of infections. 78 Interestingly, depletion of CD4+ T cells found in rats subjected to tMCAO was abrogated by treatment with levodopa for 5 days. These experiments showed the susceptibility of peripheral immune cell populations to levodopa. Treatment did not influence the level of circulating cytokines. 79 Elevated inflammatory cytokines/chemokines only show a reliable increase within the first hours following stroke. 80 Further studies are required to unravel the mechanisms through which dopamine increases the number of CD4+ cells after tMCAO. Modulation of the number of peripheral immune cell populations might be due to downmodulation of adhesion molecules CD44 and CD18, as has been shown for CD3–CD56+ NK cells stimulated by exercise-induced catecholamines. 81 Also, direct inhibitory effects on mobilization and migration of CD4+ cells can be envisaged. Interestingly, upon treatment with dopamine reduced transmigration and attenuated chemoattractant effect of IL-8 was observed in cultured LPS/TNF-α-activated primary neutrophils (CD11b+CD18+, PMN). Furthermore, dopamine attenuated the adhesion molecules E-selectin and ICAM-1 in PMN and endothelial cells, respectively. 82
Conclusion
Immune cell populations express DR. Interestingly, their expression is strongly upregulated in activated immune cells and shows specific patterns in different immune cell populations. Therefore, increased susceptibility of activated cells to dopaminergic agents has been observed in several independent studies. Interestingly, some populations of immune cells (CSF lymphocytes) also show autochthone synthesis of dopamine. After experimental stroke, we and others found an upregulation of DR on microglia/macrophages and other immune cell populations and reduced levels of pro-inflammatory molecules in the post-ischaemic brain. The exact mechanisms through which dopaminergic signalling is involved in attenuating the inflammatory response are not yet fully understood. Hence, dopaminergic treatment provides enhanced recovery of lost neurological functions after stroke.
Footnotes
Funding
This study was supported by National Funds by FCT – Foundation for Science and Technology (grant SFRH/BD/104679/2014), the Swedish Brain Fund, the Crafoord Foundation, the Swedish Research Council, Sveriges Stroke Riksförbundet, the Hans-Christian and Alice Wachtmeister Foundation and the Stiftelsen Sven-Olof Jansons livsverk.
Conflict of interest statement
The authors declare that there is no conflict of interest.