
Abstract
There is evidence of aberrations in the vitamin D–endocrine system in subjects with respiratory diseases. Vitamin D deficiency is highly prevalent in patients with respiratory diseases, and patients who receive vitamin D have significantly larger improvements in inspiratory muscle strength and maximal oxygen uptake. Studies have provided an opportunity to determine which proteins link vitamin D to respiratory pathology, including the major histocompatibility complex class II molecules, vitamin D receptor, vitamin D-binding protein, chromosome P450, Toll-like receptors, poly(ADP-ribose) polymerase-1, and the reduced form of nicotinamide adenine dinucleotide phosphate. Vitamin D also exerts its effect on respiratory diseases through cell signaling mechanisms, including matrix metalloproteinases, mitogen-activated protein kinase pathways, the Wnt/β-catenin signaling pathway, prostaglandins, reactive oxygen species, and nitric oxide synthase. In conclusion, vitamin D plays a significant role in respiratory diseases. The best form of vitamin D for use in the treatment of respiratory diseases is calcitriol because it is the active metabolite of vitamin D3 and modulates inflammatory cytokine expression. Further investigation of calcitriol in respiratory diseases is needed.
Introduction
Inadequate serum 25-hydroxyvitamin D3 (25OHD) levels are common in adults with cystic fibrosis (CF) and are associated with elevated parathyroid hormone (PTH) levels. Aiming to maintain serum 25OHD levels ≥35 ng/ml in individuals with CF decreases the risk of having a PTH level associated with secondary hyperparathyroidism and bone loss [West et al. 2011]. Short-term 1α,25-dihydroxyvitamin D3 (calcitriol) administration improved markers of calcium balance in adults with CF by increasing fractional absorption of 45Ca and lowering PTH concentrations [Brown et al. 2003]. PTH levels were significantly higher in chronic obstructive pulmonary disease (COPD) patients compared with healthy controls [Jackson et al. 2013]. In addition, there is a relationship between the tuberculin skin test, serum calcium, and PTH levels in patients with pulmonary tuberculosis (TB) [Deniz et al. 2005]. Randomized trials of vitamin D (VD) supplementation have shown effects that are likely to be protective against the development of bronchiectasis [Bartley et al. 2013]. Both calcitriol and other VD receptor (VDR) agonists significantly reduce the pro-inflammatory cytokine response to antigen challenge in CF airway epithelial cells [McNally et al. 2011]. The relationship between VD and TB has been discussed previously, and VD may have a role in TB treatment [Lương and Nguyễn, 2011]. In pulmonary TB, VD downregulated chemokine expression at the site of infection and acted as a potential anti-inflammatory agent [Selvaraj et al. 2012]. Higher maternal VD levels were associated with a lower risk of lower respiratory tract infections (RTIs) in offspring in the first year of life [Morales et al. 2012] and decreased rates of asthma in infants [Carroll et al. 2011]. Serum 25OHD levels correlated with CD4+Foxp3+ T-cell numbers in moderate/severe asthma [Chambers et al. 2012]. In the Nord-Trøndelag Health Study, low VD status was significantly associated with an increased incidence of asthma risk among men without allergic rhinitis [Mai et al. 2012]. VD deficiency enhanced allergen-induced lymphocyte responses in a mouse model of allergic airway disease [Gorman et al. 2012]. Calcitriol pretreatment enhances the efficacy of allergen immunotherapy in a mouse allergic asthma model [Ma et al. 2010]. Calcitriol also induced CD200 expression by human peripheral and airway-resident T-cells [Dimeloe et al. 2012]. VD plays a role in asthma and exerts its action through genomic and/or non-genomic pathways [Lương and Nguyễn, 2012]. Pulmonary rehabilitation is an important treatment for patients with COPD. VD supplementation during rehabilitation in COPD resulted in significantly higher median vitamin D levels compared with the levels resulting from the placebo. Patients receiving VD had significantly larger improvements in inspiratory muscle strength and maximal oxygen uptake [Hornikx et al. 2012]. In a randomized trial, high doses of VD reduced COPD exacerbations [Lehouck et al. 2012]. VD improves respiratory adjustment to fatigue and H-reflex responses in paraplegic adult rats [Bianco et al. 2011]. Hyperphosphorylation of the retinoblastoma protein (Rb) integrates growth signals from a variety of mitogen receptor pathways. Checkpoint kinase (Chk) may modulate Rb hyperphosphorylation [Verlinden et al. 2007]. Calcitriol inhibits the growth of human airway smooth muscle cells through the growth factor-induced phosphorylation of Rb and Chk1 [Damera et al. 2009]. These findings suggest that VD may play a role in respiratory diseases. Calcitriol is the best form of vitamin D for use in patients with respiratory diseases because it modulates adipokine expression and inhibits anti-inflammatory cytokine expression. However, monitoring serum 25OHD after calcitriol intake is not necessary because calcitriol inhibits the production of serum 25OHD in the liver [Bell et al. 1987; Luong and Nguyen, 1996]. Herein, we further discuss the potential role of VD in various respiratory diseases.
The role of vitamin D in respiratory diseases
Increased frequency of various respiratory diseases in VD hypovitaminosis
Abnormal VD metabolism has been reported in patients with respiratory diseases. Subjects with more severe obstructive sleep apnea (OSA) indices (apnea–hypopnea indices ≥ 15) tended to present with lower VD levels that were correlated with an increased prevalence of insulin resistance, prediabetes, and diabetes [Bozkurt et al. 2012]. COPD was associated with an increased risk of VD deficiency [Persson et al. 2012; Janssens et al. 2010]. In patients with COPD, serum 25OHD levels were significantly correlated with forced expiratory volume in 1 second (FEV1) [Janssens et al. 2010]. Total VD intake was positively associated with FEV1 [Shaheen et al. 2011]. The levels of 25OHD in patients with acute exacerbations of COPD and those with stable COPD were significantly lower than the levels in the control group. VD status is associated with bone mineral density and functional exercise capacity in patients with COPD [Romme et al. 2013]. The level of 25OHD in the COPD acute exacerbation group was significantly lower than that in the stable COPD group. VD deficiency may have a negative impact on the quality of life in COPD patients [Zhang et al. 2012a]. Low VD levels are associated with increased rejection and infections after lung transplantation [Lowery et al. 2012]. VD deficiency in healthy neonates is associated with respiratory syncytial virus bronchiolitis [Belderbos et al. 2011]. The mean serum levels of serum 25OHD among subjects with recurrent group A streptococcal tonsillo-pharyngitis were significantly lower than those of the controls [Nseir et al. 2012], suggesting an association between VD levels and recurrent group A streptococcal tonsillo-pharyngitis in adults. Children with a VD intake of less than 80 IU/kg/day were greater than four times more likely to have acute lower RTIs compared with children with a VD intake exceeding 80 IU/kg/day [Leis et al. 2012]. There was a significant correlation between VD deficiency (<15 ng/ml) and the need for supplementary oxygen and ventilator management in acute lower RTIs in hospitalized Japanese children [Inamo et al. 2011], suggesting that the immunomodulatory properties of VD may influence the severity of acute lower RTI. In a double-blind randomized controlled trial, supplementation with VD reduced the disease burden in patients with frequent RTIs [Bergman et al. 2012]. VD supplementation significantly reduced the risk of acute RTIs in winter among Mongolian children with VD deficiency [Camargo et al. 2012]. Compared with the healthy children, serum 25OHD, IgG, and IgA levels in children with bronchiolitis in the acute phase were significantly lower, and, in contrast, serum IgE levels were significantly higher. Serum 25OHD levels increased and serum IgE levels decreased significantly in the recovery phase compared with the acute phase in children with bronchiolitis. However, compared with healthy children, serum 25OHD and IgA levels were significantly lower and serum IgE levels were significantly higher in children with bronchiolitis in the recovery phase [Jiang et al. 2012], suggesting that 25OHD and immunoglobulins may play important roles in the pathogenesis of bronchiolitis. VD deficiency is common in bronchiectasis and is associated with chronic bacterial colonization and correlated with markers of disease severity in bronchiectasis [Chalmers et al. 2013].
Association of respiratory diseases with certain subtypes of the major histocompatibility complex class II molecules
Major histocompatibility complex (MHC) plays an important role in the immune system and are essential in the defense against infection. Human MHC class II molecules are encoded by three different human leukocyte antigen (HLA) isotypes, namely HLA-DR, -DQ, and -DP. Studies have suggested that several genes in the MHC region promote the susceptibility to respiratory diseases. HLA-DQB1 * 05 was the susceptibility gene identified in Uygur patients with allergic rhinitis in the Xinjiang area of China [Cui et al. 2011]. DR0301 alleles conferred resistance against severe acute respiratory syndrome (SARS) infection [Wang et al. 2011b]. The HLA class II allele DRB1*1501 is over-represented in patients with idiopathic pulmonary fibrosis (IPF) [Xue et al. 2011]. Compared with normal controls, HLA-DRB1*1454 was less prevalent in anti-neutrophil cytoplasmic antibody (ANCA)-associated systemic vasculitis patients, whereas DRB1*1101 was significantly more frequent in patients with microscopic polyangiitis. HLA-DRB1*1202 was prevalent among patients with PR3-ANCA-positive Wegener’s granulomatosis [Luo et al. 2011]. There is an increased prevalence of HLA-DRB5 gene carriers among scleroderma patients with interstitial lung disease (ILD) relative to those without ILD and healthy controls [Odani et al. 2012]. The HLA-DRB1 shared epitope was found to be associated with a reduced risk for ILD, whereas the DR2 serological group (DRB1*15 and *16) was found to be associated with an increased risk for ILD, in Japanese patients with rheumatoid arthritis [Furukawa et al. 2012]. The HLA DRB1*130101-DQB1*060101 haplotype is associated with acute chest syndrome in sickle cell anemia patients [Mahdi et al. 2009]. The HLA DRB1*1501-DQB1*0602-DPB1*0501 haplotype was found to be a genetic marker for the development of toluene diisocyanate-induced occupational asthma in Koreans [Choi et al. 2009]. Flow cytometric analysis revealed that a major portion of CD14+ macrophages in COPD have lower forward scatter, i.e. they are small macrophages. While these small macrophages accounted for 6.9% of all macrophages in control donors, the percentage of these cells in COPD patients was 45.7%. CD14 and HLA-DR expression was high on the surface of these small sputum macrophages, whereas the large sputum macrophages expressed only low levels of these surface molecules, both in control donors and COPD patients [Frankenberger et al. 2004]. In airway epithelial cells, the expression of HLA-DR in COPD patients was significantly higher than that in healthy volunteers [Qu et al. 1998]. Knockout mice with defined HLA class II deficiencies were infected with Mycobacterium bovis BCG, whereas controls were able to inhibit bacterial growth and resist infection [Ladel et al. 1995]. The HLA-DRB1*14 frequency was higher in the TB patients than in the healthy exposed patients [Duarte et al. 2011]. Numerous case-controlled studies have shown associations between class II HLA genes and asthma. HLA-DRB1, DQA1, and DQB1 alleles and haplotypes are linked to susceptibility to asthma and serum IgE production [Lara-Marquez et al. 1999; Torio et al. 2003; Movahedi et al. 2008; Choi et al. 2009; Knutsen et al. 2010], as well as to asthma severity [Juhn et al. 2007]. However, calcitriol inhibits the differentiation, maturation, activation, and survival of dendritic cells and downregulates MHC class II expression [Penna and Adorini, 2000, 2001]. Calcitriol and its analogs modulate human dendritic cells by inhibiting HLA-DR expression [Ferreira et al. 2013]. The vitamin D analog ZK203278 potently inhibits lymphocyte proliferation in the mixed lymphocyte reaction and downregulates MHC class II expression by 70% [Zügel et al. 2009]. In addition, 1α-calcidol significantly modulates the expression of HLA-DR in human peripheral blood monocytes [Scherberich et al. 2005]. Intrinsic 25-OHD activation inhibited human dendritic cell antigen presentation and chemotaxis and reduced HLA-DR expression [Bartels et al. 2010]. Calcitriol also improved graft survival in renal allografts by reducing macrophage infiltration and renal HLA-DR expression [Özdemir et al. 2011]. These findings suggest that calcitriol may have an effect on respiratory diseases by suppressing the expression of MHC class II antigens.
The role of VDRs in respiratory diseases
VDRs have been detected in the epithelia of normal and malignant human bronchial tissue [Menezes et al. 2008]. Ramirez and colleagues demonstrated VDR expression in lung tissue homogenates and lung fibroblasts [Ramirez et al. 2010]. Under calcitriol stimulation, smooth muscle cells in the airway express functional VDRs [Bossé et al. 2007]. Receptors play a crucial role in the regulation of cellular function, with small changes in their structure influencing intracellular signal transduction pathways. VDR knock-out mice exhibited an increased influx of inflammatory cells and phospho-acetylation of nuclear factor-kappa B (NF-ĸB) associated with increased pro-inflammatory mediators. This influx was linked to emphysema and reduced lung function associated with lymphoid aggregate formation compared with that of wild-type mice [Sundar et al. 2011], suggesting that VDR deficiency in mouse lung tissue can lead to an early onset of emphysema/COPD because of chronic inflammation, immune dysregulation, and lung destruction. Moreover, invariant natural killer (NK) T-cell defects in VDR knockout mice result in the failure to generate airway hyper-reactivity responses in the lung [Yu et al. 2011]. VDR polymorphisms are reported to be associated with respiratory diseases. Quadriceps weakness is an important complication of COPD and is associated with impaired exercise capacity and greater mortality [Steiner et al. 2005]. The FokI common variants in the VDR gene are associated with skeletal muscle strength in both COPD patients and control subjects, whereas the BsmI polymorphism is associated with strength only in COPD patients [Hopkinson et al. 2008]. Wittke and colleagues suggested that VDR expression in the lung micro-environment is required for the maximal induction of lung inflammation [Wittke et al. 2007]. Aspergillus fumigatus was found to downregulate VDR expression in macrophages and airway epithelial cells in CF [Coughlan et al. 2012]. VD deficiency decreases the expression of VDR in the lungs of mice with allergic airway inflammation [Agrawal et al. 2012]. Children carrying the minor T allele of the VDR (Thr1Meth) single nucleotide polymorphism (SNP) were predisposed to respiratory syncytial virus infection, as this SNP was identified as a risk factor for severe respiratory syncytial virus (RSV) disease in South African children [Kresfelder et al. 2011]. SNPs in the VDR are important in determining susceptibility to RSV bronchiolitis [Janssen et al. 2007]. The FokI ff genotype was associated with an increased incidence of acute lower RTI that was approximately seven times that of FokI FF. A weaker association with the TaqI polymorphism was also found in acute lower RTI in early childhood [Roth et al. 2012]. VDR polymorphisms are reported to be associated with an increased asthma incidence in humans. In the Chinese Han population, the VDR Apa1, but not Fok1 or Bsm1, genotype demonstrated a significant association with asthma [Saadi et al. 2009; Fang et al. 2009]. The results are similar to those observed in a family-based study in a French-Canadian founder population in which the VDR Apa1 polymorphism was found to be associated with childhood and adult asthma [Raby et al. 2004]. This association was most prominent in females. In another report, six SNPs of VDR were linked to asthma [Poon et al. 2004]. In central India, the bb genotype of the BsmI polymorphism conferred a significant risk for smear-positive and multiple-drug-resistant (MDR) pulmonary TB in several tribes (Sahariya, Bhil tribe, and Chhattisgarh tribe). Subjects from a North-Central Indian population with the FF genotype of the FokI polymorphism appeared to be less likely to develop MDR TB, whereas those with the Ff and ff genotypes were at high risk of MDR and smear-positive disease, respectively. Similarly, the tt genotype of the TaqI polymorphism was found to be associated with a high risk of smear-positive TB in both a North-Central and South-Eastern Indian population [Sharma et al. 2011]. Genetic polymorphisms in the VDR gene, either alone or in combination (haplotypes), are associated with different clinical outcomes in TB. The combined genotypes AbfT and AabbFfTT of the VDR polymorphisms were found to be statistically significantly protective against TB in the Iranian population [Marashian et al. 2010]. Taken together, these results demonstrate that VDR may have a role in respiratory diseases.
The role of cytochrome P450 lung disease
Interleukin (IL)-13 exposure enhances VD-mediated expression of the human cathelicidin 18/LL-37 by increasing CYP27B1 in bronchial epithelial cells [Schrumpf et al. 2012]. The functionality of the VDR was also demonstrated by showing a >200-fold change in the expression of the 24-hydroxylase (CYP24A1) gene following calcitriol stimulation in bronchial smooth muscle cells [Bossé et al. 2007]. Gene variants involved in the VD pathway, such as CYP24A1 (cytochrome P450 vitamin D 24-hydroxylase) and CYP2R1 (cytochrome P450 vitamin D 25-hydroxylase), are associated with asthma and atopy [Wjst et al. 2006; Bossé et al. 2009; Pillai et al. 2011]. Lasky-Su and colleagues demonstrated a genome-wide analysis (WGA) of VD levels in children with asthma, including rs10741657 (CYP2R1) [Lasky-Su et al. 2012].
The role of VD-binding protein (group-specific component or Gc-globulin) in lung diseases
High levels of calbindin, a VD-binding protein (DBP), were found in human fetal lung tissue at 14–32 weeks of gestation [Brun et al. 1987], suggesting that VD may play a role in the development of fetal lung tissue. The majority of circulating VD is bound to DBP. DBP has immunomodulatory functions that are relevant in the lung, predominantly with regard to macrophage activation and neutrophil chemotaxis, and it may contribute to the susceptibility to osteoporosis in adult Japanese women [Ezura et al. 2003]. DBP is a multifunctional protein that, in addition to the transport of VD sterols, has a major role in the actin-scavenging system that removes globular monomeric actin (G-actin) released in systemic circulation following cell damage [Speeckaert et al. 2006; White and Cooke, 2000]. Hypersensitivity pneumonitis (HP) is an immune-mediated lung disease induced by the inhalation of numerous antigens. Pathologically, chronic HP tends to exhibit usual interstitial pneumonia (UIP) and fibrotic nonspecific interstitial pneumonia (fNSIP) patterns. DBP was significantly higher in the patients with fNSIP patterns than in those with UIP patterns [Okamoto et al. 2012], suggesting that DBP may play a role in the development of pathological differences between UIP and fNSIP patterns of chronic HP. The Gc2 variant predisposed subjects with α1-antitrypsin deficiency to bronchiectasis [Wood et al. 2011]. Significantly lower mean Gc concentrations were found in the sera of CF homozygotes compared with both heterozygotes and controls [Coppenhaver et al. 1981]. The Gc2 variants appear to be protective in Whites with COPD [Horne et al. 1990; Schellenberg et al. 1998; Wood et al. 2011]. The genetic association of DBP with COPD was mediated by effects on macrophage activation [Wood et al. 2011]. In the population of the Bashkortostan Republic, Tatars with the Gc1F/1S genotype have a lower risk of COPD development, and the frequency of the Gc1F/1S genotype in COPD patients was significantly lower than that in healthy individuals [Korytina et al. 2006]. The Gc1F variant has been consistently associated with a range of COPD phenotypes in Asian subjects [Ito et al. 2004; Ishii et al. 2001; Lu et al. 2004; Shen et al. 2010]. In humans, serum VD levels were reduced by 25% in carriers of specific variants of the Gc gene, which was independent of COPD severity, smoking history, age, gender, body mass index, corticoid intake, and seasonal variation [Janssens et al. 2010]. Circulating DBP was inversely related to FEV1, in direct contrast to VD, the deficiency of which was related to a low FEV1 [Wood et al. 2011]. Patients with COPD and the Gc 1F allele showed a larger annual decline of FEV1 than patients without this allele [Ito et al. 2004]. SNPs adjacent to Gc were associated with FEV1 in a GWA study [Wilk et al. 2007]. SNPs rs17467825 and rs1155563 of the Gc gene were significantly associated with the FEV1 % predicted and FEV1/forced vital capacity (FVC), respectively, in COPD populations [Bakke et al. 2011]. Gc variants have been associated with diffuse panbronchiolitis [Ishii et al. 2001]. DBP is elevated in serum during human and bovine mycobacterial infection [Emmett et al. 1987; Seth et al. 2009]. Gc2 homozygotes were more common in TB patients relative to controls in Asian populations [Papiha et al. 1983; Martineau et al. 2010]. Low circulating levels of DBP have been reported in patients with acute lung injury [Lind et al. 1988]. In addition, DBP expression was upregulated in patients with isocyanate-occupational asthma (OA) [Hur et al. 2008], and the serum DBP level was used as a serological marker for the detection of isocyanate-OA among workers exposed to isocyanate [Kim et al. 2012a]. Gc2 was significantly associated with the risk of asthma in the Chinese Han population [Li et al. 2011], and circulating 25OHD levels in premenopausal women were strongly related to Gc polymorphisms [Sinotte et al. 2009]. The neutrophil chemotactic activity of C5a and C5a des Arg can be significantly enhanced by Gc [Kew and Webster, 1988], and calcitriol, a bioactive form of VD, inhibits the C5a chemotactic function of Gc [Shah et al. 2006]. Taken together, these results demonstrate that DBP may have a role in respiratory diseases.
The role of TLRs in lung disease
Toll-like receptors (TLRs) are a group of glycoproteins that function as surface trans-membrane receptors. These receptors are involved in the innate immune response to exogenous pathogenic micro-organisms. The relative levels of TLR2, TLR4, and NF-κB expression in sinus tissues from chronic rhinosinusitis patients were significantly higher than those of controls [Sun et al. 2012b]. Cigarette smoke increases TLR4 and TLR9 expression and induces cytokine production from CD8+ T cells in COPD [Nadigel et al. 2011]. TLR4 upregulation underpins airway neutrophilia in smokers with COPD and acute respiratory failure [Pace et al. 2011]. TLR2 expression was significantly higher in COPD participants compared with healthy controls [Baines et al. 2011]. TLR2 expression was upregulated on monocytes obtained from TB pleural fluid; TLR2 and TLR4 expression was also enhanced on interferon (IFN)-γ-secreting CD4+ T cells [Prabha et al. 2008]. The TLR2 pathway is important in regulating swine facility organic dust-induced airway inflammation [Poole et al. 2011], which suggests the importance of TLR2 agonists in mediating large animal farming-induced airway inflammatory responses. TLR2 was found to mediate fatal immunopathology in mice during the treatment of secondary pneumococcal pneumonia following influenza and bleomycin-induced acute lung injury, inflammation, and fibrosis in mice [Karlström et al. 2011; Liu et al. 2010]. Cyptoporus polysaccharide prevents lipopolysaccharide (LPS)-induced acute lung injury associated with the downregulation of TLR2 expression [Yao et al. 2011]. In a model of secondary Pseudomonas aeruginosa pneumonia in post-septic mice, TLR2 deficiency improved survival by promoting efficient bacterial clearance and restoring a pro-inflammatory cytokine balance in the lung [Pène et al. 2012]. Melanoma differentiation-associated gene (MDA)-5 and TLR3 initiate pro-inflammatory signaling pathways leading to rhinovirus-induced airway inflammation and hyper-responsiveness. Rhinovirus-infected MDA5 and TLR3 null mice showed reduced lung inflammatory responses and reduced airway responsiveness [Wang et al. 2011a]. Workers in swine operations are exposed to indoor dusts and gases and are at increased risk of respiratory problems. Workers in swine operations with the TLR2-16933T/A polymorphism (AA) had significantly greater mean values of lung function than workers in swine operations with wild-type genotypes [Gao et al. 2013], suggesting the possibility that TLR2 polymorphisms are protective against airway diseases in individuals exposed to Gram-positive organisms in inhaled airborne dust. An association was also reported between a SNP (T1486C) in the TLR-9 gene and the onset of COPD and sarcoidosis [Pabst et al. 2013]. Among patients with systemic sclerosis, the rare TLR2 Pro631His variant is robustly associated with antitopoisomerase positivity, the diffuse form of the disease, and the development of pulmonary arterial hypertension [Broen et al. 2012]. A TLR5 (g.1174C > T) variant that encodes a stop codon (R392X) is associated with bronchopulmonary dysplasia [Sampath et al. 2012]. Two SNPs in TLR2 were associated with an increased risk of chronic rhinosinusitis in Korean subjects [Park et al. 2011]. However, calcitriol suppresses the expression of TLR2 and TLR4 mRNA and protein in human monocytes and triggers hyporesponsiveness to pathogen-associated molecular patterns [Sadeghi et al. 2006]. Calcitriol has also been shown to downregulate intracellular TLR2, TLR, and TLR9 expression in human monocytes [Dickie et al. 2010]. Interestingly, TLR activation results in the expression of the VDR and 1α-vitamin D hydroxylase in human monocytes, leading to the induction of the antimicrobial peptide cathelicidin and the killing of intracellular Mycobacterium tuberculosis [Liu et al. 2006]. Calcitriol can cause expression of cathelicidin in bronchial epithelial cells [Yim et al. 2007] and may enhance the production of cathelicidin LL-37 [Rivas-Santiago et al. 2008]. Poor VD intake may increase susceptibility to M. tuberculosis infection by inefficiently supporting the induction of cathelicidin mRNA expression in monocytes [Liu et al. 2006, 2007]. The addition of a VDR antagonist inhibited the induction of cathelicidin mRNA by more than 80%; consequently, the protein expression of this antimicrobial agent was reduced by approximately 70% [Liu et al. 2006]. Furthermore, the knockdown of cathelicidin in primary monocytes results in the loss of TLR-mediated antimicrobial activity against intracellular mycobacteria [Liu et al. 2009b]. Upon M. tuberculosis stimulation, 15alcitriol modulates cytokine production toward an anti-inflammatory profile by decreasing the expression of TLR2 and TLR4 while increasing cathelicidin production [Khoo et al. 2011]. Calcitriol also induces autophagy in human monocytes via cathelicidin and leads to the colocalization of mycobacterial phagosomes and autophagosomes in human macrophages in a cathelicidin-dependent manner [Yuk et al. 2009]. Taken together, these data indicate that VD may play a role in respiratory diseases by modulating the TLR pathways.
The role of poly(ADP-ribose) polymerases in lung diseases
Poly(ADP-ribose) polymerases (PARPs) comprise a family of enzymes sharing a conserved catalytic domain that support mono- or poly(ADP-ribosyl) transferase activity using nicotinamide adenine dinucleotide (NAD+) as a donor of ADP-ribosyl units. PARPs are involved in a wide range of molecular and cellular processes, including the maintenance of genome stability, regulation of chromatin structure and transcription, cell proliferation, and apoptosis [Krishnakumar and Kraus, 2010]. Expression of the DNA repair enzyme PARP is a known response to oxidative DNA damage. Pre-exposure to hyperoxia causes increased lung injury and epithelial apoptosis in mice ventilated with high tidal volumes. Analysis of these lung homogenates showed that lung injury was associated with the cleavage of executioner caspases and their downstream substrate PARP-1 [Makena et al. 2010]. Increased PARP-1 expression and activity are associated with inflammation in murine models of allergen-induced airway inflammation [Havranek et al. 2010]. Activation of PARP-1 is a central mechanism of LPS-induced acute lung inflammation [Albertini et al. 2000; Liaudet et al. 2002]. Cigarette smoke-induced autophagy is regulated by a NAD+-dependent deacetylase sirtuin 1 (SIRT1)-PARP-1-dependent mechanism and is suggested in the pathogenesis of COPD [Hwang et al. 2010]. The percentage of PAR polymer-positive lymphocytes appeared to be higher in COPD patients than in healthy age-matched controls [Hageman et al. 2003]. Overactivation of PARP played an important role in the inflammatory and transcriptional pathogenesis of lung injury induced by mechanical ventilation, and PARP inhibition has potentially beneficial effects on the prevention and treatment of lung injury induced by mechanical ventilation [Kim et al. 2008]. High tidal volume ventilation resulted in lung injury, hypotension, renal hypoperfusion, and impaired renal endothelium-dependent vasodilation, which are associated with renal dysfunction and tissue changes. Peroxynitrite-induced PARP activation is involved in renal hypoperfusion, impaired endothelium-dependent vasodilation and resultant dysfunction, and injury in a model of lung injury [Vaschetto et al. 2010]. The scavenging of peroxynitrite or inhibition of PARP afforded protection against acute kidney injury induced by a double-hit lung injury [Matejovic and Radermacher, 2010]. Increased PARP-1 activity has been shown to occur with lung inflammation in acute respiratory distress syndrome (ARDS) and hyperoxic lung injury [Pagano et al. 2007]. An increase in immunoreactivity to nitrotyrosine and PARP, as well as a significant loss of body weight and mortality, was observed in the lungs of bleomycin-treated mice. Treatment with PARP inhibitors reduced the development of inflammation and tissue injury events induced by bleomycin administration in the mice [Genovese et al. 2005]. The PARP-1-inhibiting flavonoids fisetin and tricetin were able to attenuate LPS-induced cytokine release from the leukocytes of patients with COPD and in a mouse model of LPS-induced acute pulmonary inflammation [Weseler et al. 2009; Geraets et al. 2009]. PARP-1 inhibition prevents eosinophil recruitment by modulating Th2 cytokines in a murine model of allergic airway inflammation [Oumouna et al. 2006]. Theophylline prevents NAD+ depletion via PARP-1 inhibition in human pulmonary epithelial cells [Moonen et al. 2005]. Inhibition of PARP attenuates endotoxin-induced dysfunction of pulmonary vasorelaxation [Pulido et al. 1999]. However, treatment with dexamethasone and VD attenuates neuro-inflammatory age-related changes in the rat hippocampus, and caspase-3 and PARP were all attenuated in hippocampal tissue prepared from rats that received dexamethasone and VD [Moore et al. 2007]. Increased levels of VD seem to downregulate PARP-1 expression; PARP-1 levels decrease following 18 alcitriol treatment in NB4 cells, which are acute promyelocytic leukemia cells [Bhatia et al. 1995]. VD exerts a concentration-dependent inhibitory effect on PARP-1 in human keratinocyte cells [Mabley et al. 2007]. VD-induced downregulation of PARP is further enhanced by nicotinamide in human myeloblastic leukemia cells [Shen and Yen, 2009]. These findings suggest that VD may have a protective role in respiratory diseases by down-regulating PARP.
The role of the reduced form of the nicotinamide adenine dinucleotide phosphate oxidase enzyme complex in lung diseases
Nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (Nox) mediates critical physiological and pathological processes, including cell signaling, inflammation, and mitogenesis, by generating reactive oxygen species (ROS) from molecular oxygen. The pathogenesis of IPF has been linked to oxidative stress, and evidence has implicated a role for ROS-generating Nox enzymes in pulmonary fibrosis. In the lungs of IPF patients, the expression of Nox4 has been demonstrated in hyperplastic alveolar type II cells [Amara et al. 2010]. ROS generation by Nox4 is a key player in epithelial cell death leading to pulmonary fibrosis [Carnesecchi et al. 2011]. Nox-4 is upregulated in the lungs of mice subjected to noninfectious injury and in cases of human IPF, and it also mediates myofibroblast activation and fibrogenic responses to lung injury [Hecker et al. 2009]. Nox4 mRNA and protein expression was upregulated in pulmonary fibroblasts from patients with IPF and was correlated with the mRNA expression of smooth muscle alpha-actin and procollagen I [Amara et al. 2010]. Three weeks after the administration of bleomycin, wild-type mice developed massive fibrosis, whereas Nox4-deficient mice displayed almost normal lung histology [Carnesecchi et al. 2011]. During hypoxia associated with acute lung injury in mice, Nox1 is an important contributor to ROS production and cell death in the alveolo-capillary barrier [Carnesecchi et al. 2009]. A genome-wide microarray of primary airway smooth muscle identified increased messenger RNA expression of Nox4 in asthma [Sutcliffe et al. 2012]. Nox1 and Nox4 mRNA and proteins had increased levels of expression in allergic nasal mucosa and nasal polyps compared with healthy nasal mucosa [Moon et al. 2009]. Treatment with hydrogen sulfide attenuated hyperoxia-induced acute lung injury in mice and abated oxidative stress marked by reduced Nox activity [Li et al. 2013]. Pediatric OSA may lead to neurocognitive dysfunction, although not in everyone affected. The frequencies of Noxp22phox polymorphisms were similar between children with OSA and controls, except for rs6520785 and rs4673, the latter being significantly more frequent among OSA children without deficits than in those with deficits. Similarly, 8-hydroxydeoxyguanine urine levels and Nox activity were lower among children without cognitive deficits and particularly among those with the rs4673 polymorphism [Gozal et al. 2012], suggesting that polymorphisms within the Nox gene or its functional subunits may account for important components of the variance in cognitive function deficits associated with OSA in children. Apocynin, a Nox inhibitor, significantly attenuated ovalbumin-induced airway hyper-responsiveness and inflammation [Kim et al. 2012b]. However, VD deprivation in rats decreased the activity of cytosolic NADPH-dependent 3,5,3’-triodo-L-thyronine binding in the liver. This decrease can be restored by administering calcitriol [Hashizume et al. 1991]. In heart mitochondria, NAD+-dependent isocitrate dehydrogenase notably decreased in D VD-deficient rats, but calcitriol treatment subsequently restored normal values [Stio et al. 1994]. In rat centrilobular hepatocytes, a VD-deficient diet induced a significant increase in NADPH [Bachelet et al. 1983]. Husain and colleagues reported that cardiac NOX activity increased by 300% in uremic rats compared with the normal controls [Husain et al. 2009]. Treatment with paricalcitol protected the uremic rats from cardiac oxidative stress by inhibiting NOX activity (by 50%), thus lowering superoxide production in the heart. Taken together, these findings and results indicate that VD may have a role in respiratory diseases via the suppression of NADPH expression.
The role of MMPs in lungs disease
Matrix metalloproteinases (MMPs) are proteolytic enzymes that are responsible for remodeling the extracellular matrix and regulating leukocyte migration through the extracellular matrix. This migration is an important step in inflammatory and infectious pathophysiology. MMPs are produced by many cell types, including lymphocytes, granulocytes, astrocytes, and activated macrophages. There is growing evidence that MMPs play an important role in the pathogenesis of respiratory diseases. The expression levels of the MMP-8 and MMP-9 genes in both peripheral blood mononuclear cells and neutrophils are associated with viral lower RTI severity [Brand et al. 2012], suggesting the usefulness of MMP-8 and MMP-9 as potential markers of disease severity in viral respiratory infections. The levels of MMP-9 and TIMP-1 increased significantly with increasing disease severity in both asthmatic and COPD groups [Dar et al. 2012]. The early elevation of MMP-8 and MMP-9 in pediatric ARDS may lead to complications of prolonged mechanical ventilation [Kong et al. 2011]. MMP-8 levels were strongly associated with mortality due to respiratory disease in patients with rheumatoid arthritis [Mattey et al. 2012].The absence of MMP-8 increases neutrophilic lung infiltration after intraperitoneal LPS injection in mice [González-López et al. 2012]. COPD patients had increased serum MMP-9 concentrations compared with the control group. In the COPD group, the MMP-9 levels were negatively correlated with FEV1 [Brajer et al. 2008]. MMP-12 is markedly increased in the induced sputum from patients with stable COPD compared with controls [Demedts et al. 2006]. MMP-9 plays an important role in the structural changes consequent to oxygen-induced lung injury [Chetty et al. 2008]. MMP-9 plays a pathophysiological role in the remodeling in ILD and lung fibrosis associated with systemic sclerosis [Andersen et al. 2007]. The expression of MMP-1, MMP-2, MMP-3, MMP-7, and MMP-9 was increased in human astrocytes that were stimulated by conditioned medium from M. tuberculosis-infected monocytes [Harris et al. 2007]. MMP-mediated proteolysis of the mycobacterial HSP65 protein was reported to contribute to the complex immunomodulatory interplay during the course of TB infection [Shiryaev et al. 2011]. 1G genotypes of the MMP-1 polymorphism are associated with asthma with persistent airway obstruction, and the heterozygous 1G genotype (1G/2G) results in the greatest susceptibility to persistent airway obstruction in asthma [Huang et al. 2009]. A functional polymorphism in MMP-9 is associated with childhood atopic asthma [Nakashima et al. 2006]. VDR knockout mice have been shown to have an influx of inflammatory cells, phospho-acetylation of NF-κB, and upregulated expression of MMP-2, MMP-9, and MMP-12 in the lungs [Sundar et al. 2011]. The VDR TaqI polymorphism is associated with the decreased production of TIMP-1, a natural MMP-9 inhibitor [Timms et al. 2002]. In addition, calcitriol was found to modulate tissue MMP expression under experimental conditions [Dean et al. 1996], to downregulate MMP-9 levels in keratinocytes, and to possibly attenuate the deleterious effects of excessive tumor necrosis factor (TNF)-α-induced proteolytic activity, which is associated with cutaneous inflammation [Bahar-shany et al. 2010]. Calcitriol also inhibits both the basal levels and the staphylococcus-stimulated production of MMP-9 in human blood monocytes and alveolar macrophages [Lacraz et al. 1994]. Moreover, a VD analog has also been reported to reduce the expression of MMP-2, MMP-9, and PTH-related peptide in Lewis lung carcinoma cells [Nakagawa et al. 2005]. Together, these studies suggest that calcitriol may play an important role in the pathological processes of respiratory diseases by downregulating the level of MMPs and upregulating the level of TIMPs.
The role of the mammalian family of mitogen-activated protein kinases in lung diseases
Mitogen-activated protein kinases (MAPKs) include extracellular signal-regulated kinase (ERK), p38, and c-Jun NH2-terminal kinase (JNK), with each MAPK signaling pathway consisting of at least three components, a MAPK 3 kinase (MAP3K), a MAPK 2 kinase (MAP2K), and a MAPK. The MAPK pathways are activated by diverse extracellular and intracellular stimuli, including peptide growth factors, cytokines, hormones, and various cellular stressors (e.g. oxidative stress and endoplasmic reticulum stress). These signaling pathways regulate a variety of cellular activities, including proliferation, differentiation, survival, and death [Kim and Choi, 2010]. The increased expression of p38 MAPK was observed in human bronchial epithelium after LPS exposure [Roos-Engstand et al. 2005]. Activated MAPKs are significantly increased in lung homogenates from patients with IPF compared with controls [Yoshida et al. 2002]. Phospho-p38+ alveolar macrophages and alcitri-p38+ cells in alveolar walls were increased in patients with severe and mild/moderate COPD compared with smoking and nonsmoking controls. Moreover, they were inversely correlated with values of FEV1 and FEV1/FVC [Renda et al. 2008]. Muscle atrophy in COPD is associated with reduced exercise tolerance, muscle strength, and survival. MAPK signaling in the quadriceps was significantly elevated in patients with COPD compared with controls. In addition, the phosphorylated-to-total p38 MAPK ratio and phosphorylated-to-total ERK ½ ratio were negatively associated with the mid-thigh muscle cross-sectional area [Lemire et al. 2012]. These findings suggested that the MAPKs may play a role in the development of muscle atrophy in COPD. Compared with the healthy control or convalescent cases, pro-inflammatory cytokines, such as IL-1, TNF-α, and IL-8, and the MAPK signaling pathway were significantly upregulated in SARS patients [Yu et al. 2005]. P38 MAPK signaling plays a key role in regulating CXC chemokine production in septic lung injury, and the inhibition of p38 MAPK activity abolishes the pulmonary infiltration of leukocytes as well as lung edema [Asaduzzaman et al. 2008]. Blocking p38 MAP kinase signaling inhibits complement-induced acute lung injury in a murine model [Nash and Heuertz, 2005]. SB 239063 and FR-167653, which are p38 MAPK inhibitors, reduce neutrophilia, inflammatory cytokines, MMP-9, and fibrosis in a model of lung fibrosis and also ameliorate bleomycin-induced pulmonary fibrosis [Underwood et al. 2000; Matsuoka et al. 2002]. These findings suggested that p38 MAPK inhibition offers a protective effect in the lungs from both neutrophil influx and protein leakage associated with acute lung injury. By regulating VDR mRNA expression, the p38 MAPK pathway participates in the mediation of calcium signals and affects lipid accumulation in murine pre-adipocytes [Sun et al. 2012a]. Pretreatment with calcitriol has been shown to inhibit JNK activation by all stressors, as well as to inhibit p38 activation in keratinocytes [Ravid et al. 2002]. Zhang and colleagues demonstrated that the up-regulation of MAKP-1 by VD inhibited LPS-induced p38 activation and cytokine production in monocytes and macrophages [Zhang et al. 2012b]. In another study, the VD analog (24R)-1,24-dihydroxycholecalciferol prevented neuronal damage caused by hydrogen peroxide-induced toxicity in the SH-SY5Y cell line [Tetich et al. 2004]. Interestingly, the neurotoxic effects of H2O2 were dependent on JNK and p38 MAPK. In addition, the long-term actions of VD in MCF-7 and LNCaP cancer cells can suppress the estradiol-induced activity of ERK-1 MAPK and can inhibit cell growth [Liakhovich et al. 2000]. Taken together, VD may play a role in respiratory diseases by modulating MAPK.
The role of the Wnt/β-catenin signaling pathway in lung disease
Wnt/β-catenin plays pivotal roles in regulating cell growth, cell development, and the differentiation of normal stem cells. Activation of the Wnt/β-catenin signaling pathway by mechanical ventilation is associated with pulmonary fibrosis in healthy lungs [Villar et al. 2011], suggesting that the activation of this pathway could play an important role in both induced by mechanical ventilation lung injury and lung repair. Activation of WnT/β-catenin signaling in pulmonary fibroblasts by TGF-β1 is increased in COPD [Baarsma et al. 2011]. Functional Wnt signaling is also increased in IPF [Königshoff et al. 2008]. Aberrant Wnt/β-catenin pathway activation was demonstrated in IPF [Chilosi et al. 2003]. WnT5A may play a role in fibroblast proliferation and survival characteristics in IPF and other fibrotic interstitial lung diseases [Vuga et al. 2009]. WnT1-inducible signaling protein-1 mediates pulmonary fibrosis in mice and is upregulated in humans with IPF [Königshoff et al. 2009]. Nuclear β-catenin is increased in systemic sclerosis pulmonary fibrosis and promotes lung fibroblast migration and proliferation [Lam et al. 2011]. The WnT/β-catenin pathway is modulated in asthma patients; β-catenin also plays a significant role in the LPS-induced inflammatory response, as indicated by small interfering RNA transfection experiments [Lee et al. 2012]. These results suggest that β-catenin plays a role as a negative regulator, preventing the overproduction of inflammatory cytokines. The Wnt/β-catenin pathway is activated in bleomycin-induced lung fibrosis [Liu et al. 2009a]. The blockade of the Wnt/β-catenin pathway attenuates bleomycin-induced pulmonary fibrosis [Kim et al. 2011]. Dickkopf (DKK) proteins are known to regulate WnT signaling via their interaction with Kremen receptors. Enhanced expression of DKK1 and DKK4 was detected in the lung homogenates of IPF patients compared with transplant donors [Pfaff et al. 2011]. The inhibition of Wnt/β-catenin/cAMP response element-binding protein signaling reverses pulmonary fibrosis [Henderson et al. 2010]. However, calcitriol inhibits β-catenin transcriptional activity by promoting the binding of β-catenin and VDR and the induction of E-cadherin expression [Pálmer et al. 2001]. Paricalcitol, a VD analog, has been shown to suppress β-catenin-mediated gene transcription and ameliorate proteinuria and kidney injury in calcitriol nephropathy [He et al. 2011]. Most VDR variants fail to activate the VD-responsive promoter and fail to bind β-catenin or regulate its activity [Byers and Shah, 2007]. Interestingly, VDR depletion has been shown to enhance Wnt/β-catenin signaling and tumor burden in colon cancer [Larriba et al. 2011]. Taken together, these results demonstrate that VD plays a role in respiratory diseases by modulating the Wnt/β-catenin signaling pathway.
The role of the prostaglandins in lung diseases
Prostaglandins (PGs) play an important role in inflammatory processes. Cyclooxygenase (COX) participates in the conversion of arachidonic acid into PGs. Patients with severe OSA had higher urinary 8-iso-PGF2α and lower serum levels of nitrite/nitrate. After 6 months of nasal continuous positive airway pressure (CPAP) treatment, a significant decrease in urinary 8-iso-PGF2α was observed, whereas serum levels of nitrite/nitrate showed only a minor increase [Del Ben et al. 2012]. 8-Isoprostane levels were significantly higher in subjects with COPD and smokers than in non-smokers [Inonu et al. 2012]. PGE2 is a bioactive prostanoid implicated in the inflammatory processes of acute lung injury/ARDS. PGE2 enhances IL-8 production via the EP4 receptor in human pulmonary microvascular endothelial cells [Aso et al. 2012]. In vitro and in vivo studies using PGES-/- mice, which cannot produce PGE2, showed a significantly higher mycobacterial burden than that found in control mice. These findings suggested that PGE2 plays a critical role in the inhibition of M. tuberculosis replication [Chen et al. 2008]. In TB patients, the production of PGE was increased in both lungs, but the hyperproduction of PGF2α occurred in the impaired lung [Kaminskaia et al. 1991]. Influenza A virus induces interleukin-27 through COX-2 and protein kinase A signaling [Liu et al. 2012]. Enzootic pneumonia caused by Mycoplasma hyopneumoniae and pleuropneumonia caused by Actinobacillus pleuropneumoniae are among the most common and economically relevant pulmonary diseases in swine herds. Swine lungs showed a significantly higher activity and expression of 5-lipoxygenase and COX-2 in a wide range of cell types [Giacominelli-Stuffler et al. 2012], thus suggesting the likely involvement of both enzymes in the pathogenesis of bacterial porcine pneumonia. Fetal lung prostaglandin H synthase-2 (PGHS-2) mRNA and PGE synthase (PGES) mRNA increased 2 and 7 days after LPS exposure [Westover et al. 2012]. The -197T > C and -613C > T alleles of the PGD2 receptor gene were found to be significantly associated with allergic asthma and allergy to pollen and mites [Isidoro-García et al. 2011]. Moreover, the combination of relative COX-2 selective nonsteroidal anti-inflammatory drugs (NSAIDs) and aspirin is associated with higher bone mineral density at multiple skeletal sites in men and women compared with controls [Carbone et al. 2003]. Calcitriol is reported to regulate the expression of several key genes involved in the PG pathway, causing a decrease in PG synthesis [Moreno et al. 2005]. Calcitriol pretreatment limited PG biosynthesis by cytokine-stimulated adult human osteoblast-like cells [Keeting et al. 1998]. Calcitriol and its analogs have been shown to selectively inhibit the activity of COX-2 [Aparna et al. 2008]. Calcitriol was shown to induce NAD+-dependent 15-hydroxyprostaglandin dehydrogenase (15-PGDH) in human neonatal monocytes. In addition, the COX-2 mRNA and PGE2 levels were decreased in the culture media compared with the controls [Pichaud et al. 1997]. 15-PGDH is the key enzyme of PGE2 catabolism. In addition, PGE1 was reported to increase in vivo and in vitro calcitriol biosynthesis in rabbits [Velásquez-Forero et al. 2006]. These findings suggest that VD may play a role in modulating the inflammatory process in respiratory diseases.
The role of the ROS in lung disease
ROS are produced by activated phagocytes as part of their microbicidal activities. The presence of oxidative markers, such as nitrogen oxides and hydrogen peroxide (H2O2), was demonstrated in an animal model of LPS-induced acute lung injury [de Broucker et al. 2012]. LPS treatment of pulmonary vascular endothelial cells induced the generation of ROS and caused oxidative stress associated with endothelial cell hyperpermeability, cytoskeletal remodeling, and formation of paracellular gaps, as well as the activation of Rho, p38 stress kinase, and NF-κB signaling, which are the hallmarks of endothelial barrier dysfunction [Kratzer et al. 2012]. ROS-induced arterial dysfunction in patients with OSA and exacerbated the effect of CPAP treatment [Del Ben et al. 2012]. Treatment with H2S markedly prolonged the survival of mice under oxygen exposure and reduced malondialdehyde (MDA) and peroxynitrite formation [Li et al. 2013]. The overexpression of O2– was demonstrated in allergic nasal mucosa [Moon et al. 2009]. The serum ROS levels in active and chronic lung TB cases were significantly higher than those in the control group [Ceylan et al. 2005]. The levels of the lipid peroxidation end product, 9,11-linoleic acid, and free radical activity were higher in the active pulmonary TB cases than in the inactive pulmonary TB cases [Jack et al. 1994]. H2O2 levels were significantly higher in subjects with COPD and in smokers than in nonsmokers [Inonu et al. 2012]. Plasma total antioxidant status and total oxidant status levels are elevated in children with allergic rhinitis compared with the control group [Emin et al. 2012], suggesting that the high level of total oxidant indicates that these patients are exposed to severe oxidative stress. OSA patients showed increased protein oxidative damage and impaired antioxidant defenses [Mancuso et al. 2012]. OSA and COPD patients showed nocturnal hypoxemia, with a minimum oxygen saturation ranging from 53–92% and 50–87%, respectively. The mean levels of thiobarbituric acid reactive substances were 15.7 ± 3.6 and 15.3 ± 3.4 nmol MDA/ml in OSA and COPD patients, respectively, whereas the mean level in the control group was 4.1 ± 0.2 nmol MDA/ml. Nocturnal desaturated COPD patients showed increased levels of lipid peroxidation [Okur et al. 2013]. Similarly, calcitriol has been reported to exert a receptor-mediated effect on the secretion of H2O2 by human monocytes [Gluck and Weinberg, 1987]. Human monocytes in culture gradually lose their capacity to produce O2– when stimulated. The addition of calcitriol, LPS, or lipoteichoic acid restored the capacity of stimulated monocytes to produce O2– and increased their oxidative capacity compared with that of unstimulated monocytes [Cohen et al. 1986]. Calcitriol can also protect nonmalignant prostate cells from oxidative stress-induced cell death by eliminating ROS-induced cellular injuries [Levy and Malech, 1991]. VD metabolites and analogs have been reported to induce lipoxygenase mRNA expression, lipoxygenase activity, and ROS production in a human bone cell line [Bao et al. 2008]. In another study, the VD analog (24R)-1,24-dihydroxycholecalciferol prevented neuronal damage caused by H2O2-induced toxicity in the SH-SY5Y cell line [Tetich et al. 2004]. VD can also reduce the extent of lipid peroxidation and can induce SOD activity in a hepatic anti-oxidant system in rats [Somjen et al. 2011]. Astrocytes play a pivotal role in the central nervous system (CNS) detoxification pathways in which glutathione (GSH) is involved in eliminating oxygen and nitrogen reactive species, such as nitric oxide (NO). Calcitriol affects this pathway by enhancing intracellular GSH pools and significantly reduces the nitrite production that is induced by LPS [Garcion et al. 1999]. These findings suggest that VD modulates oxidative stress in respiratory diseases.
The role of the nitric oxide synthase in lung diseases
Nitric oxide synthase (NOS) is an enzyme that is involved in the synthesis of nitric oxide (NO), which regulates a variety of important physiological responses, including cell migration, the immune response, and apoptosis. In most murine models, iNOS is essential for the killing of intracellular mycobacteria [Chan et al. 1992]. iNOS-/- mice proved highly susceptible to M. tuberculosis, which exhibited faster replication in the lungs of these mice than in the lungs of hosts deficient in other genes [Macmicking et al. 1997]. iNOS inhibitors also render similar deleterious effects on TB infection in mice, as assessed by mortality, bacterial burden, and pathological tissue damage [Chan et al. 1995]. eNOS-/- mice developed increased pulmonary arterial pressure after 6 months of cigarette smoke exposure, which was associated with vascular remodeling and increased vascular nitrotyrosine staining [Wright et al. 2012]. Enhanced levels of iNOS in influenza-infected lungs were observed during increased pulmonary neutrophilia [Wheeler et al. 2013]. iNOS was significantly upregulated in the lung at day 4 after LPS exposure. When iNOS-/- mice were exposed to intratracheal LPS, early lung injury was attenuated [D’Alessio et al. 2012]. iNOS inhibition attenuates physical stress-induced lung hyper-responsiveness and oxidative stress in animals with lung inflammation [Marques et al. 2012]. Conversely, the activation of 1α-hydroxylase in macrophages increases the level of calcitriol, which inhibits iNOS expression and reduces NO production within LPS-stimulated macrophages [Chang et al. 2004]. Thus, calcitriol production by macrophages may provide protection against the oxidative injuries that are caused by the NO burst. Calcitriol is known to inhibit LPS-induced immune activation in human endothelial cells [Garcion et al. 1997]. In experimental allergic encephalomyelitis, calcitriol inhibits the expression of iNOS in the rat CNS [Equils et al. 2006]. The arteries from VD-deficient offspring have an impaired ability to relax due to deficiencies in the production of two important factors, namely NO and endothelium-derived hyperpolarizing factor. VD-deficient female offspring have an additional impairment in the NO signaling pathway in the arterial muscle [Tare et al. 2011].
Figure 1 illustrates the role of VD in various lung diseases.
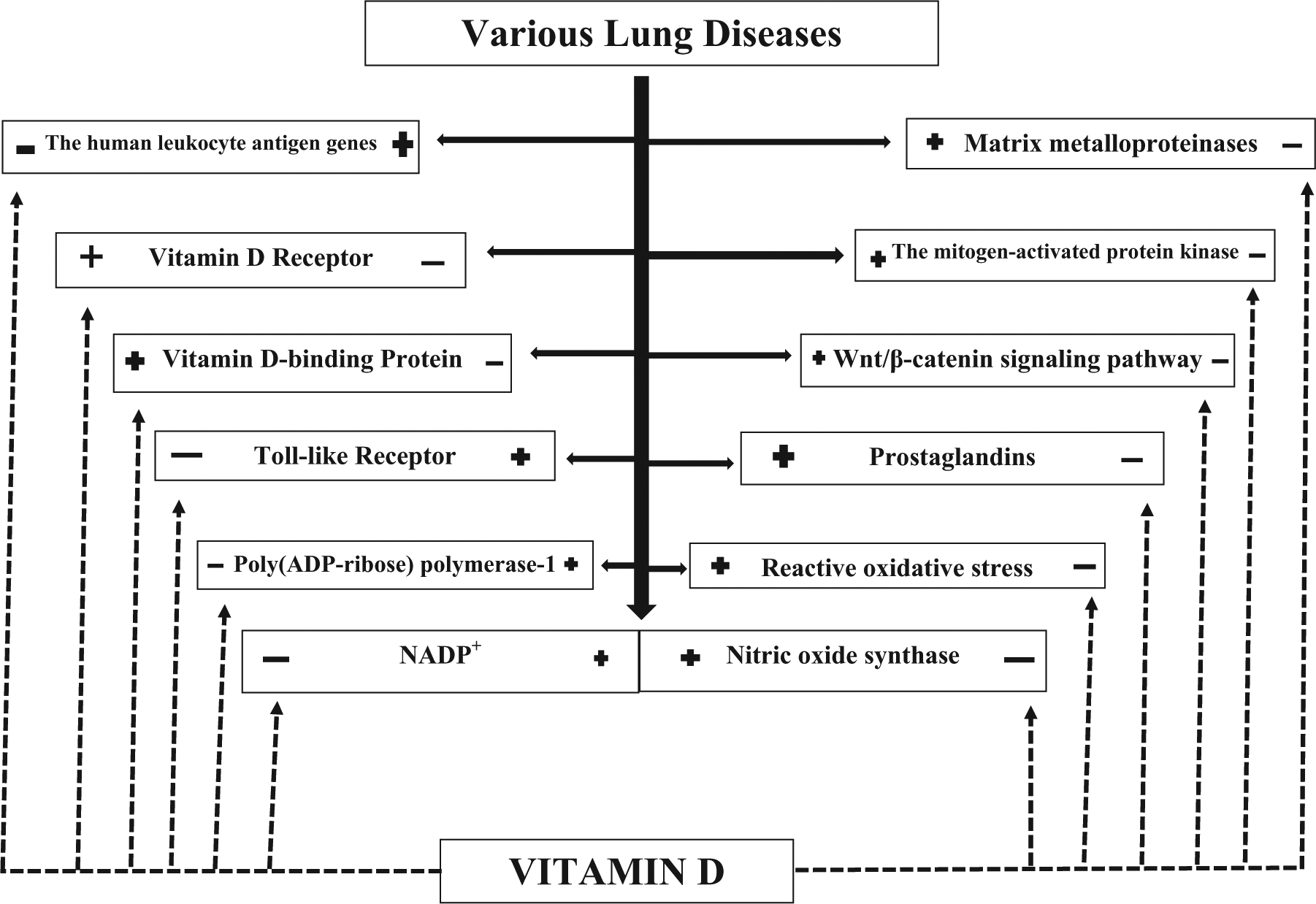
The role of vitamin D in various lung diseases.
Conclusions
In this review, the most recent data are overviewed on the role of VD insufficiency in the pathogenesis of respiratory diseases. A lot of clinical and experimental data indicates that in many respiratory diseases lack of sufficient VD production underlies the pathology. Respiratory diseases linked to insufficient VD production are overviewed. Specific genetic dispositions making the patients prone to respiratory diseases are also reviewed. Finally, the intercellular and intracellular mediators setting in motion various pathological cascades in the respiratory organs are explained, where VD exerts its protective effect by inhibiting them. These include among others inhibition of the PARP-1, the MAPK, the Wnt/ß-catenin, and the NOS-signaling systems. Reduced production of ROS, various cytokines, PGs also contribute to the beneficial actions of VD in various lung diseases.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of interest statement
The authors declare no conflicts of interest in preparing this article.