
Abstract
Non-typeable Haemophilus influenzae (NTHi) is the most common respiratory pathogen in patients with chronic obstructive disease. Limited data is available investigating the impact of NTHi infections on cellular re-differentiation processes in the bronchial mucosa. The aim of this study was to assess the effects of stimulation with NTHi on the bronchial epithelium regarding cellular re-differentiation processes using primary bronchial epithelial cells harvested from infection-free patients undergoing bronchoscopy. The cells were then cultivated using an air-liquid interface and stimulated with NTHi and TGF-β. Markers of epithelial and mesenchymal cells were analyzed using immunofluorescence, Western blot and qRT-PCR. Stimulation with both NTHi and TGF-ß led to a marked increase in the expression of the mesenchymal marker vimentin, while E-cadherin as an epithelial marker maintained a stable expression throughout the experiments. Furthermore, expression of collagen 4 and the matrix-metallopeptidases 2 and 9 were increased after stimulation, while the expression of tissue inhibitors of metallopeptidases was not affected by pathogen stimulation. In this study we show a direct pathogen-induced trans-differentiation of primary bronchial epithelial cells resulting in a co-localization of epithelial and mesenchymal markers and an up-regulation of extracellular matrix components.
Keywords
Introduction
Sustained bronchial inflammation is a hallmark in the pathogenesis of chronic obstructive pulmonary disease (COPD). The inflammatory response is maintained by repeated exposure to noxious agents, especially cigarette smoke, and leads to a persistently elevated concentration of inflammatory cells and cytokines in the airways.1,2
Infection-associated acute exacerbations accelerate lung function decline, but, as recent studies show, bacterial colonization of the lower airways is also a trigger of chronic inflammatory responses.3–5 One of the key pathogens in stable COPD patients, as well as in exacerbations, is non-typeable Haemophilus influenzae (NTHi).3,6
The goal of this study was to evaluate interactions between NTHi and the human bronchial epithelium, including possible effects of NTHi infection on airway remodeling. Special focus was put on trans-differentiation processes in the bronchial epithelium, in accordance with the concept of epithelial–mesenchymal transition (EMT). This concept proposes a loss of epithelial markers and subsequent acquisition of mesenchymal markers and has been well characterized in several studies utilizing cell culture,7–10 mainly using TGF-β as a stimulant.
Previous studies showed a gain of mesenchymal markers and, in particular, collagen expression in tissue from COPD patients11–13 compared with control subjects, but there is only limited evidence of EMT occurring in vivo so far. To the best of the authors’ knowledge, no studies have decidedly focused on pathogen-associated EMT in COPD.
To gain insight into the role of NTHi infections in cellular trans-differentiation processes ex vivo, we established a model of primary human bronchial epithelial cells (pBECs) which were stimulated with NTHi or TGF-β, a growth factor well known for its immunomodulatory and fibrogenic properties and for its central role in EMT induction.9,14
To establish experimental conditions as close to the physiological setting as possible, we cultivated the pBECs using an air–liquid interface (ALI).
Material and methods
Culture and differentiation of primary human bronchial epithelial cells
pBECs were harvested from infection-free patients (assessed by clinical parameters and laboratory results) undergoing bronchoscopy for diagnostic reasons (for patient characteristics, see Table 1).
Patient characteristics.

The local ethics committee approved the use of human cells (University of Lübeck, AZ 11-098) and each volunteer signed a written consent form beforehand. The cells were harvested using a cytobrush (Medscand Medical, Tomelilla, Sweden) and kept in RPMI 1640 Medium (Biochrom, Berlin, Germany) at 4°C until preparation, which took place 4 h (at most) after cell extraction. The cells were then purified and transferred into a petri dish containing Airway Epithelial Cell Growth Medium, to which imipenem and cilastatin were added (Promocell, Heidelberg, Germany, see manufacturer’s page for detailed content). Cells were incubated at 37°C and 5% CO2 until they reached 80-90% confluence, taking approximately 10 d. Then, they were passaged, washed and re-suspended in antibiotic-free medium (no antibiotics were added to the medium from that point). A total of 200,000 cells were layered onto a 0.4 µm pore-size cell culture membrane insert (Greiner Bio One Thincert™ 12 Well (Kremsmünster, Austria)). The insert was cultivated submerged in medium for 2 d, followed by exposure of the cell layers’ apical surface to air by removal of the upper compartments’ medium. The cells were left at the ALI for another 4 d to polarize and minimize fibroblast contamination, as assessed by Western blot analysis of vimentin expression, which was indiscernible at that point (data not shown). After this period, the cells were incubated with 500 μl medium alone or stimulated with medium containing 10 ng/ml recombinant human TGF-β (R&D Systems, Minneapolis, US) or 106 CFU/ml NTHi. The NTHi strain used in these experiments (Isolate B5144) was isolated in our university hospital from a 60-yr-old COPD patient with invasive pneumonia. Characterization as NTHi was made using biochemical methods (the requirement of factor x and V for bacterial growth and negative slide serum agglutination tests) after initial identification as H. influenzae.
Stimulants or medium were added to the apical side of the inserts and removed without additional washing steps after 24 h of stimulation. Samples were taken 24 h, 72 h and 8 d after stimulation. All experiments were carried out using four different donors.
For the extensive preliminary tests needed to establish the ALI model, we harvested cells from 19 patients in total. Of these, the culture of four patient’s cells had to be terminated, in three cases due to contamination which, in all cases, was clearly discernible after the initial 24 h of culture. In one case, there was only limited cell proliferation, probably due to low initial cell numbers.
There was no discernible cell necrosis throughout the entire length of the experiment, as assessed by LDH release assay (Cytotoxicity Detection Kit, Roche, Basel, Switzerland; data not shown).
qPCR for markers of EMT
RNA in the samples was isolated using the NucleoSpin® RNA II kit (Macherey-Nagel, Düren, Germany) and then reverse transcribed into cDNA (RevertAid Reverse Transcriptase, Fermentas, Waltham, US) according to manufacturer’s instructions. Then, qPCR was performed using a Lightcycler® 1.5 (Roche). Results were normalized to 18s-RNA and mRNA content was determined by assessing the cycle-thres hold. value using the double-delta-CT method (as shown in the results section).
Used primers were: E-cadherin: forward 5′-TGTTTCTTCGGAGGAGAGCG-3′, reverse 5′-AAGATACCGGGGGACACTCA-3′, vimentin: forward 5′-TTCCAAACTTTTCCTCCCTGA-3′, reverse 5′-AATATATTGCTGCACTGAGTGTG-3′; TGF-β1 forward 5′-GCAACAATTCCTGGCGATACC-3′, reverse 5′-CTCCACGGCTCAACCACT-3′′; MMP 2: forward 5′-CTACGATGGAGGCGCTAAT-3′, reverse 5′-GCCATAGAAGGTGTTCAGGT-3′ MMP9: forward 5′-CGACGTCTTCCAGTACCGA-3′, reverse 5′-CCCAGTGGGGATTTACATGG-3′; TIMP1 forward 5′-ATACTTCCACAGGTCCCACAA-3′, reverse 5′-GGGATGGATAAACAGGGAAAC-′3; TIMP2: 5′-AGATGCACATCACCCTCTGTG-3′, reverse 5′-TGCCCGTTGATGTTCTTCTCT-3′; collagen 1: forward 5′-TCCCCAGCCACAAAGAGTCTA-3′, reverse 5′-TTTCCACACGTCTCGGTCAT-3′, collagen 4: forward 5′-TTTCCTGGTACATCTCTGCC-3′, reverse 5′-CCTTTCTCTCCAATTTCGCC-3′; and 18s: forward 5′-TCAAGAACGAAAGTCGGAGG-3’, reverse 5′-GGACATCTAAGGGCATCACA-3′.
Immunofluorescence staining
The membrane the cells adhered to was gently removed from the insert and, after fixation with 4% paraformaldehyde, blocking and permeabilization (1% BSA, 0.3% Triton-X in PBS), stained overnight (14–16 h) using vimentin Ab (1:50, Cell Signaling, Boston, US), E-cadherin-Ab (1:100, Bio Legend, San Diego, US) and DAPI nuclear stain (10 µg/ml, DAKO, Jena, Germany) according to manufacturer’s instructions. As secondary Abs, DyLight™ 488 donkey anti-mouse IgG and DyLight™ 594 donkey anti-rabbit IgG (both BioLegend) were used in a dilution of 1:100 and incubated for 1 h. Results were documented using a Zeiss Axioscope equipped with a Zeiss Axiocam (both Zeiss, Oberkochen, Germany).
Western blot analysis
After cell lysis, denatured proteins were separated by PAGE and subsequently blotted onto nitrocellulose membranes. For staining, Abs for vimentin (1:2000, Cell Signaling) and E-cadherin (1:4000, BD Transduction Laboratories, San Jose, US) were used according to manufacturer’s instructions, incubating overnight. For normalization, anti-β-actin-Ab was used (1:2000, Cell Signaling). Pictures were documented using a chemiluminescence imaging system (Fusion Fx7, Vilber-Lourmat, Eberhardzell, Germany) and densitometry was performed using ImageJ, Version 1.52p.
Statistical analysis
Statistical analysis and visualization were performed using GraphPad Prism 7. One-way analysis of variance was performed, followed by Dunnet’s multiple comparisons test when statistical significance was reached. The level of significance was P < 0.05.
Results
Expression of E-cadherin and vimentin in pBECs in response to stimulation by NTHi or TGF-β
pBECs showed a strong E-cadherin expression that remained unchanged after stimulation throughout the entire duration of the experiment. In contrast, vimentin expression was markedly enhanced by both TGF-β and NTHi stimulation. Thus, a co-localization of E-cadherin and vimentin was observed after stimulation, as shown by immunofluorescence staining (Figure 1a–c).
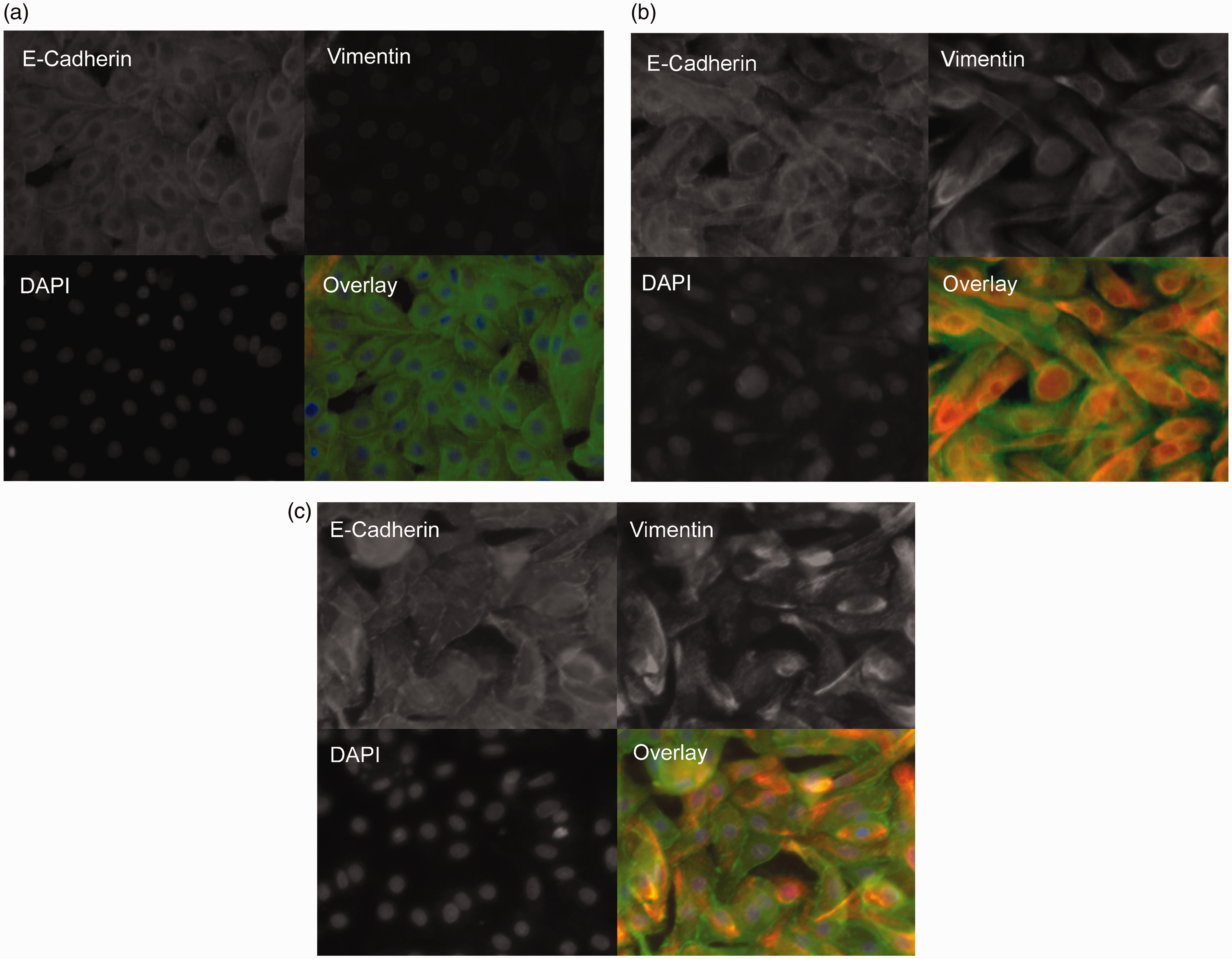
Vimentin expression is regulated by NTHi and TGF-β-stimulation in human pBECs. Immunofluorescence staining with detection of E-cadherin and vimentin in pBECs from ALI culture 8 d after stimulation with TGF-β (10 ng/ml) or NTHi (106 CFU/ml). Representative image at 400x magnification. E-Cadherin: green signals, vimentin: red signals, DAPI nuclear stain: blue signals.(a) Medium control(b) NTHi-stimulation(c) TGF-β-stimulation
In addition, the cell phenotype changed after stimulation, showing a spindle-shaped morphology compared to the medium control. Stimulation-induced effects on marker expression were confirmed by PCR (Figure 2) and by Western blot (Figure 3). The numerical increase in E-cadherin expression in the PCR after NTHi stimulation was not statistically significant and caused by one outlying value.
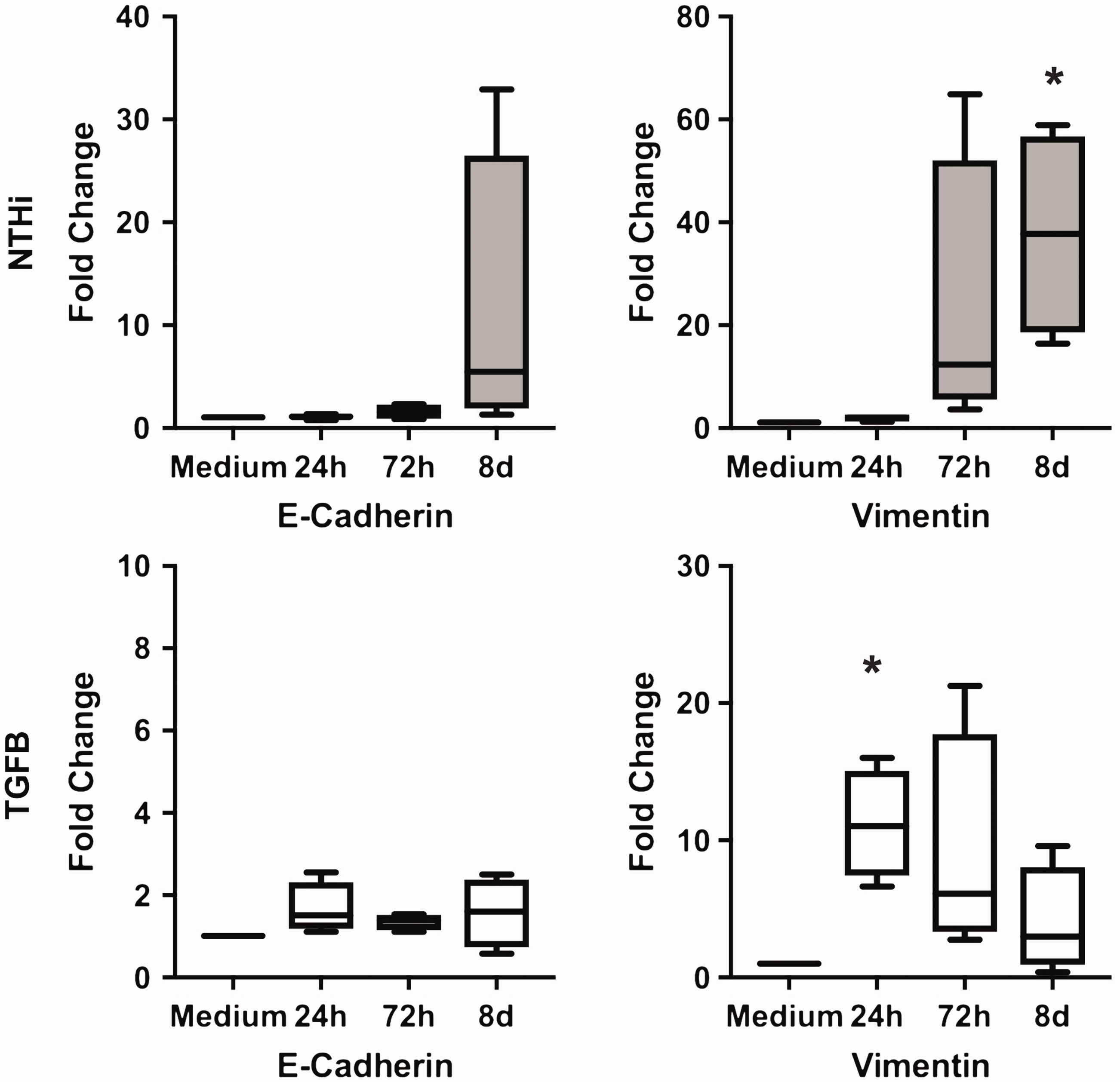
Vimentin expression is enhanced by both NTHi and TGF-β. E-Cadherin and vimentin expression in pBECs (PCR), expressed as relative increase of gene expression/medium 24 h, 72 h, and 8 d after stimulation. Box extension is from 25th to 75th percentile, whiskers from minimum to maximum value of n = 4 experiments. Stimulation by NTHi (106 CFU/ml) and by TGF-β (10 ng/ml). * P < 0.05.
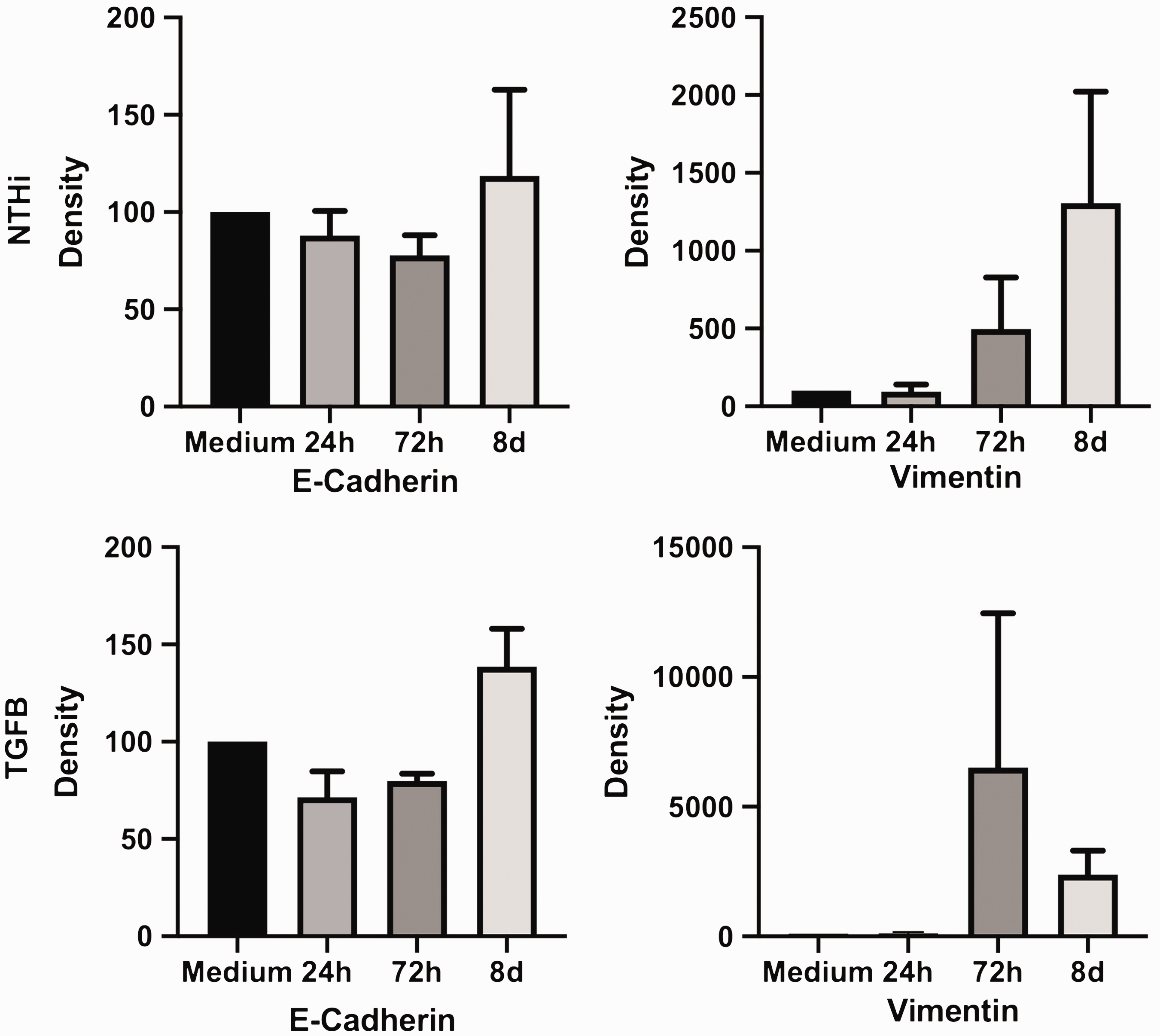
Increase in vimentin protein levels after stimulation with NTHi and TGF-β. E-Cadherin and vimentin protein levels in pBECs (Western blot), expressed as relative increase of band density/medium 24 h, 72 h, and 8 d after stimulation. Bar extension displays mean, whiskers SEM of n = 4 experiments (24 h and 8 d after stimulation) or n = 3 experiments (72 h after stimulation). Stimulation by NTHi (106 CFU/ml) and by TGF-β (10 ng/ml).
MMP and collagen induction by NTHi and TGF-β
To further investigate NTHi-induced changes of molecules relevant for ECM composition, expression of mediators and products of mesenchymal metabolism were analyzed.
MMP9 expression was markedly up-regulated in response to both NTHi and TGF-β-stimulation (Figure 4).
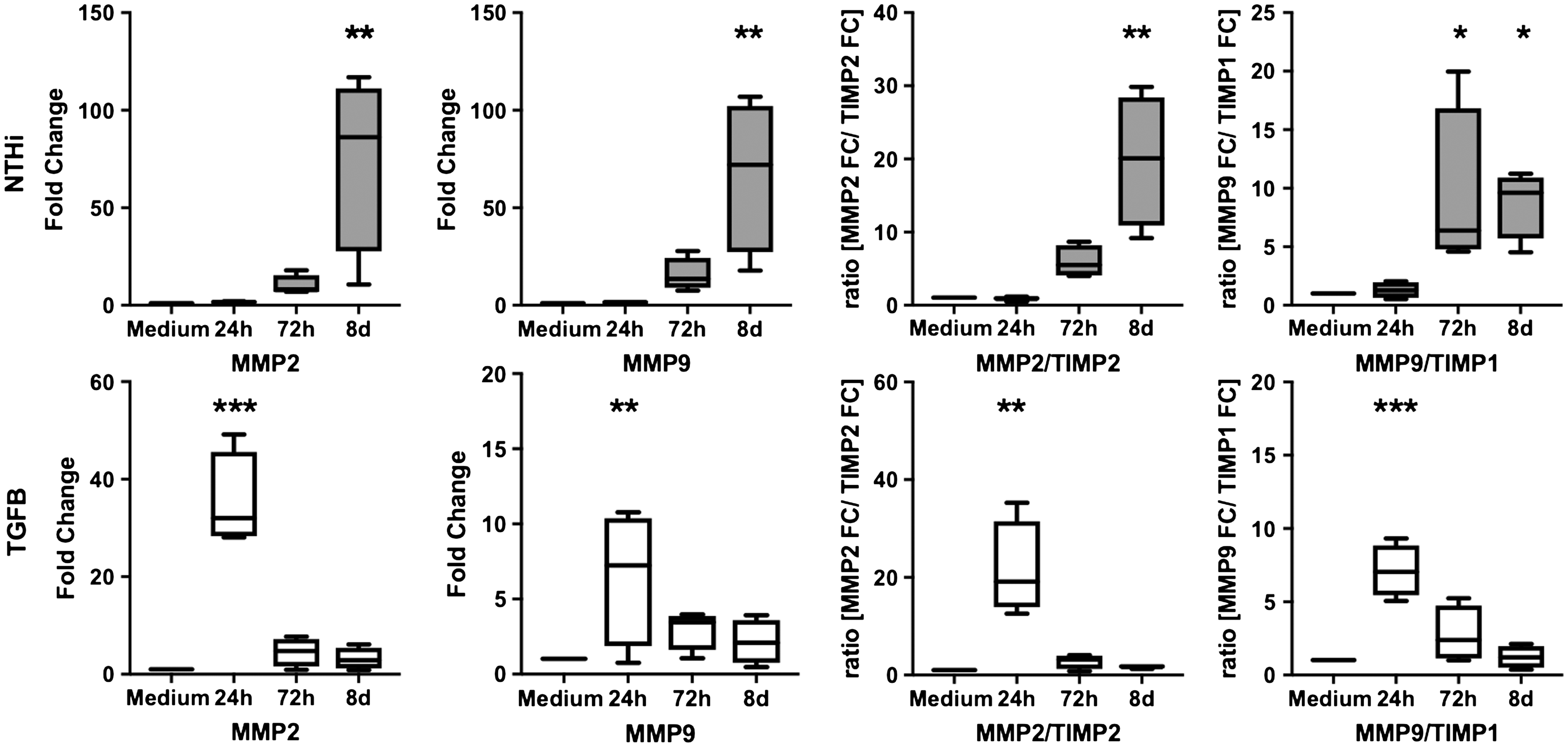
Stimulation by both NTHi and TGF-β leads to up-regulation of MMP2 and MMP9, while TIMP1 and TIMP2 expression remains stable. Expression of MMP2 and MMP9 and the expression ratio of MMP2/TIMP2 and MMP9/TIMP1 in pBECs (PCR), shown as relative increase of gene expression/medium 24 h, 72 h, and 8 d after stimulation. Box extension is from 25th to 75th percentile, whiskers from minimum to maximum value of n = 4 experiments. Stimulation by NTHi (106 CFU/ml) and by TGF-β (10 ng/ml). * P < 0.05. ** P < 0.001. *** P < 0.0001.
Stimulation of the MMP9-inhibitor TIMP1 remained far below the changes seen with MMP9, as demonstrated by a significantly elevated MMP9/TIMP1 ratio. Similar effects were seen in the expression patterns of MMP2 and its inhibitor TIMP2, resulting in a significant up-regulation of the MMP2/TIMP2 ratio (Figure 4).
Collagen type I expression numerically increased 24 h after TGF-β-stimulation and 8 d after NTHi-stimulation, respectively (Figure 5). Collagen 4 expression was significantly induced, again 24 h after TGF-β-stimulation and 8 d after NTHi stimulation (Figure 5).
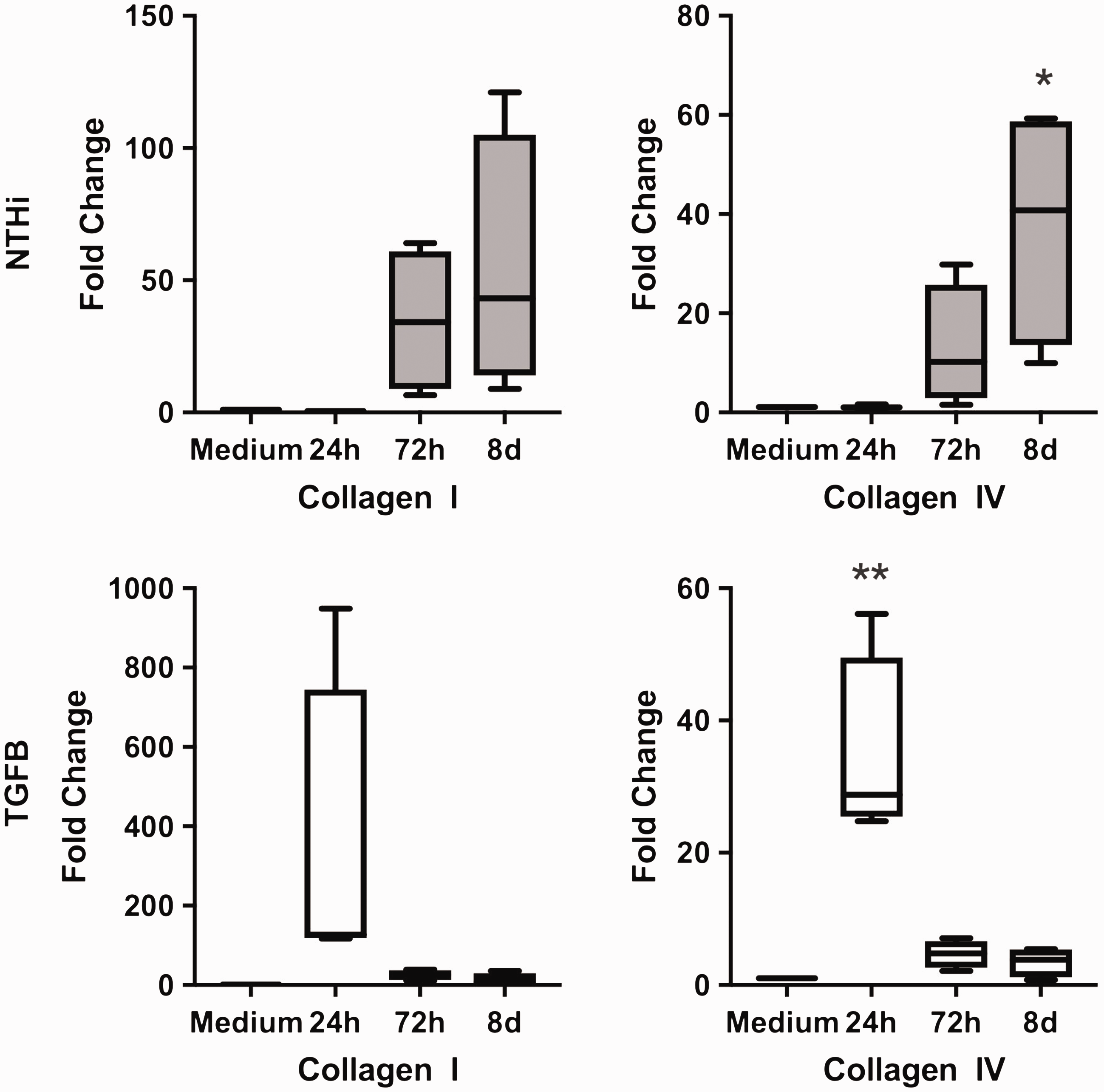
Collagen 1 and collagen 4 expression is stimulated by NTHi and TGF-β. Collagen 1 and collagen 4 expression in pBECs (PCR), expressed as relative increase of gene expression/medium 24 h, 72 h, and 8 d after stimulation. Box extension is from 25th to 75th percentile, whiskers from minimum to maximum value of n = 4 experiments. Stimulation by NTHi (106 CFU/ml) and by TGF-β (10 ng/ml). * P < 0.05. ** P < 0.001.
To elucidate possible mechanisms of the changes, we analyzed the TGF-β-expression. While no significant autoinduction was recorded (data not shown), we could show a significant up-regulation of TGF-β expression after stimulation with NTHi (Figure 6).
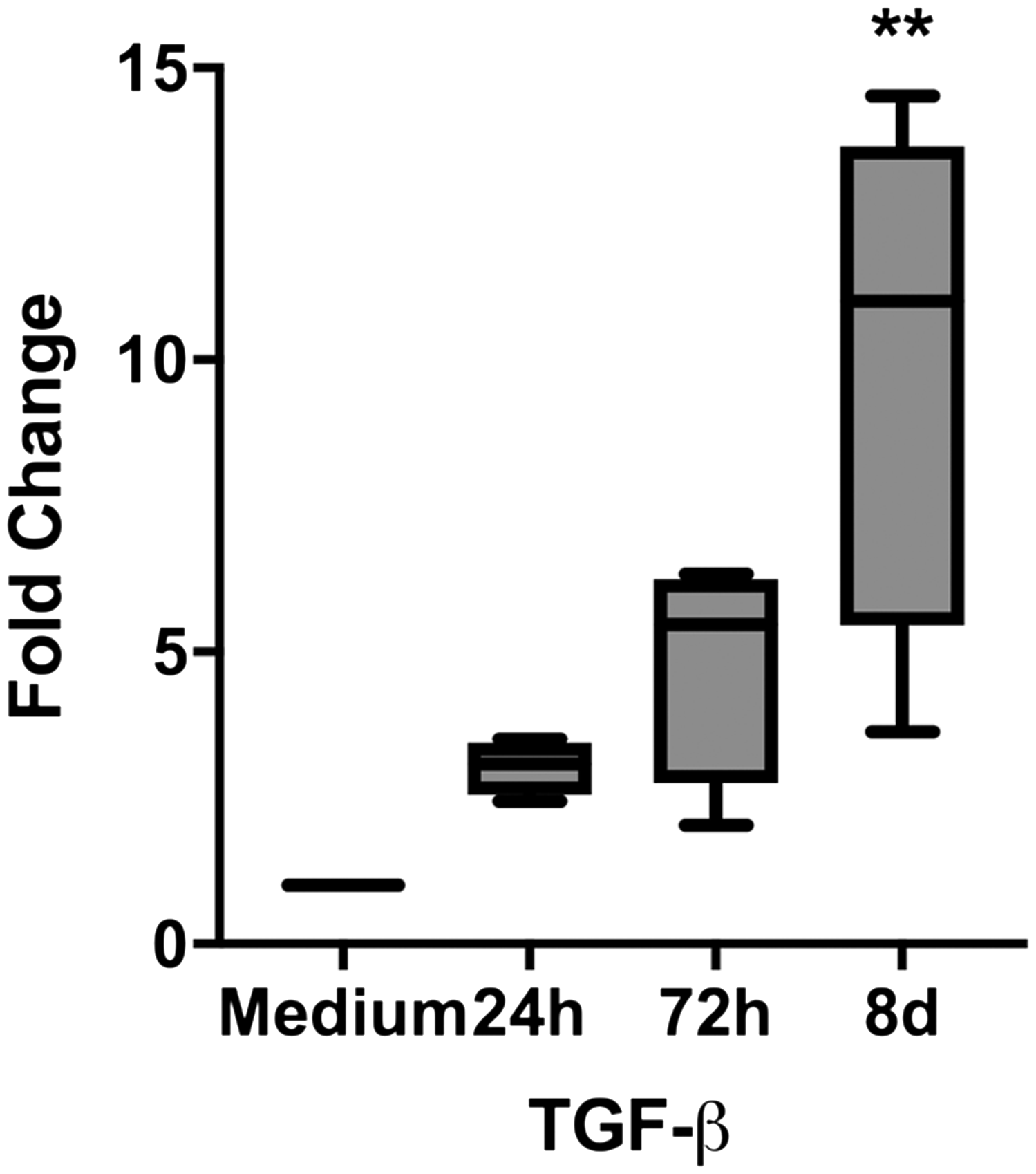
TGF-β expression is stimulated by NTHi. TGF-β expression in pBECs (PCR), expressed as relative increase of gene expression/medium 24 h, 72 h, and 8 d after stimulation. Box extension is from 25th to 75th percentile, whiskers from minimum to maximum value of n = 4 experiments. Stimulation by NTHi (106 CFU/ml). ** P < 0.001.
Discussion
In this study we demonstrated a pathogen-induced up-regulation of mesenchymal markers in pBECs. Very few studies concerning cellular trans-differentiation and EMT in the bronchial epithelium have addressed the impact of infection on these processes and, to our knowledge, none used a direct pathogen stimulation.
TGF-β, on the other hand, has been extensively studied as a stimulant in cellular trans-differentiation processes in various settings, leading to a decrease in epithelial markers, while mesenchymal markers were up-regulated to varying degrees.7,8,10 Interestingly, while in the immortalized cell line BEAS-2B we were able to reproduce these results showing a loss of E-cadherin after stimulation with both TGF-β and NTHi (data not shown), there was a different response in pBECs to both TGF-β and NTHi. We observed a strong expression of the epithelial marker E-cadherin that remained unchanged throughout the experiments. By contrast, vimentin as a mesenchymal marker was markedly up-regulated after stimulation with both TGF-β and NTHi, while being indiscernible in the medium control.
We reproduced these results using three methods: by analyzing the protein levels using Western blot and immunofluorescence staining and by analyzing the gene expression using PCR.
The apparent contradiction to EMT studies where a loss of epithelial markers was commonly seen is most likely due to methodological differences. Firstly, many EMT studies used immortalized cell lines, which, although easy to cultivate, producing reproducible results, are not easily comparable to primary cells, as demonstrated in studies showing different cellular responses to pathogen stimulation of immortalized cell lines in contrast to NHBE.22,23 In some studies focusing on EMT, commercially-available normal human bronchial epithelial cells (NHBE) were utilized,7,8,10 although they also show differences to freshly sampled cells.
Secondly, only a few EMT studies focused on the bronchial epithelium have used the ALI as a cultivation method so far. This technique is a more physiological approach to studying the bronchial epithelium since cells can polarize more easily due to the contact with ambient air, resulting in cell functions closer to the situation in vivo than in submerged cultures. When the focus is particularly on the trans-differentiation of cells, the most physiological approach should be preferred. In some recently published studies, this issue was mainly addressed in the field of in vitro toxicology.16–18 In these studies, no decreased cell viability was found in the cells cultured in the ALI when compared with liquid interface cultures. It was concluded that the ALI culture model was a viable and potentially more exact method.
With regards to cellular trans-differentiation, there are only a few studies available using the ALI cultivation method. In a study using mouse tracheal epithelial cells cultivated in the ALI, Kuroishi et al. demonstrated a TGF-β-induced co-localization of vimentin and α-SMA with the epithelial marker ZO-1. 19 Another study from Gohy et al. used pBECs from non-smokers, smokers, and COPD patients, which were cultivated in the ALI. 20 At the baseline, there was a higher vimentin expression and a lower E-cadherin expression in immunohistochemical staining in COPD patients compared with the controls. Stimulation with TGF-β led to a higher number of vimentin-staining cells, a process that was reversible with TGF-β-Ab treatment. EMT reversibility was also confirmed by Hacket et al. using pBECs obtained from donor lungs and bronchial brushings. The main finding was an elevation of vimentin and α-SMA, while expression of E-cadherin and, to a lesser degree, ZO-1 was reduced after TGF-β-stimulation. 21
Using immunofluorescence, we demonstrated a co-localization of epithelial and mesenchymal markers lasting 8 d after stimulation. The significantly induced vimentin expression after stimulation confirms previous findings using ALI cultivation in pBECs, while the co-localization of vimentin and E-cadherin is, to our knowledge, a new aspect that has not been shown so far. Our data implies that a loss of epithelial markers is not a prerequisite for a gain of mesenchymal functions, suggesting more plasticity of pBECs than previously acknowledged. The reversibility of EMT after treatment with anti-TGF-β-Abs in vitro, as previously shown, is in line with this observation.20,21
Our study also supports a broader approach to EMT in general. Kalluri and Weinberg proposed a differentiation of EMT into three subtypes: Type I EMT occurs during organogenesis, while Type II EMT as seen in response to injury, is characterized by a temporary increase in mesenchymal markers and maintains a stable expression of epithelial markers. Following this model, a loss of epithelial markers such as E-cadherin is only seen in Type III EMT, where it is a prerequisite for invasive behavior of dedifferentiated epithelial cancer cells. 24 The increased mesenchymal marker profile seen in tissue of COPD patients is most likely in concurrence with Type II EMT, as are the results demonstrated in this study.
To characterize functional implications of phenotypic changes of pBECs, we analyzed the expression of several mediators that are predominantly products of mesenchymal cells. We demonstrated that both TGF-β and NTHi significantly induce the expression of MMPs 2 and 9, as well as collagen 4, without relevant impact on TIMP expression, leading to an elevated MMP/TIMP ratio. In vivo, these changes could lead to both an enhanced breakdown of extracellular matrix and increased collagen deposition, which is a characteristic finding in the small airways of COPD patients.11,12 In a previous study, peribronchiolar fibrosis could be induced in mice by exposing them to NTHi lysates, 15 although the source of the collagen was not identified.
In conclusion, we demonstrated for the first time that NTHi infection is associated with cellular trans-differentiation processes in pBECs ex vivo and that these effects are comparable to the effects of TGF-β-stimulation. We could also show a significant induction of TGF-β-expression after NTHi exposure, which could explain the different kinetics of the expression changes, with TGF-β-induced changes being at their maximum 24 h after stimulation and NTHi-induced changes most noticeable at the latest examination point 8 d after stimulation. This observation points to an indirect response to NTHi infection via cytokine induction, although further studies are required to confirm this hypothesis, ideally using TGF-β-neutralizing antibodies.
In a follow-up study using a larger cohort, it would be interesting to see whether the changes we show in this paper differ regarding patient’s characteristics (such as smoking history, lung function, and whether COPD was previously diagnosed or not). However, in this study we had no evidence of a significant difference in behavior of the cells we used regarding the patient’s characteristics.
In the era of a more differentiated approach to the lung microbiome, in vivo stimulation with one pathogen cannot claim to portray the more complex reality. However, NTHi has been shown to be a leading microorganism orchestrating temporary changes of the respiratory microbiome during acute exacerbations. 25 The findings in this paper should lead to a broader approach regarding the impact of pathogen colonization on epithelial-mesenchymal trans-differentiation, combining clinical and in vivo methods.
The use of patient-acquired pBECs and their cultivation at the ALI as shown in this work is well suited for this purpose and should be taken into consideration more frequently in studies concerning cellular functions at the mucosal interface in the human respiratory tract.
Footnotes
Acknowledgements
The authors thank B Füssel, T Tietz, U Knuppertz and J Hofmeister for their technical assistance.
Ethical approval
This study was approved by the ethics committee of the University of Lübeck, AZ 11-098.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Deutsche Forschungsgemeinschaft of TG and DD (247915558). The funding body was not involved in any experimental decisions, work, conclusions drawn or the draft of the manuscript. In addition, this work was supported by the German Center for Lung Research (DZL) and the University of Lübeck.