
Abstract
Survivin is an anti-apoptosis protein that may be associated with the development of eosinophilia; the latter is associated with the pathogenesis of many immune disorders. Here we report that less apoptotic eosinophils (Eos) were induced in those isolated from mice suffering from food allergy (FA) than those from naive mice after treating with cisplatin in vitro. Exposure to cisplatin induced more Fas ligand (FasL) expression in Eos isolated from naive mice than in those of FA mouse. Survivin was detected in the intestinal tissue extracts in much higher amounts in the FA group than in the naive group. Immunohistochemistry showed that epithelial cells were the major source of survivin in the intestine. Exposure to IL-4 or IL-13 up-regulated the expression of survivin in intestinal epithelial cells. Survivin interfered with the expression of FasL in Eos. Inhibition of survivin attenuated the eosinophilia-related inflammation in the intestine. In conclusion, intestinal epithelial cell-produced survivin induced defects in apoptosis in Eos to contribute to eosinophilia in the intestine. Inhibition of survivin can suppress the eosinophilia-related intestinal inflammation. The data suggest that survivin may be a novel target for the treatment of FA.
Introduction
Eosinophilia indicates a condition that the number of eosinophils (Eos) is unusually increased in the local tissue. In the cytoplasm of Eos, there are a large number of granules containing chemical mediators, including major basic protein (MBP), eosinophil cationic protein (ECP), eosinophil peroxidase (EPX), eosinophil-derived neurotoxin (EDN). 1 Upon activation, Eos release the contents of granules into local tissues to induce tissue damage and inflammation. 1 Eosinophilia is associated with the pathogenesis of a large number of inflammatory diseases, such as allergic asthma, food allergy (FA), rheumatic arthritis, autoimmune diseases, eosinophilic enteropathy, and eosinophilic leukemia. 2 It is believed that the mechanism of eosinophilia formation is overdifferentiation of Eos. 3 The Eos differentiation has been extensively studied and published data indicate that IL-5 plays a critical role here. 3 While a number of reports indicated that the induction of Eos apoptosis can ameliorate allergic diseases, 4 the regulation of the apoptotic machinery in Eos remains largely unknown.
Apoptosis is a physiological and non-inflammatory process to eliminate the unwanted cells to maintain the homeostasis in the body. Defects in apoptosis in cells may result in an overproliferation of the cell type, such as cancer cells that usually have defects in apoptosis which results in cancer growth. 5 Fas and Fas ligand (FasL) play an important role in the regulation of immune cell lifespan via the apoptosis pathway. Deregulation of apoptosis may result in autoimmune diseases in the body. 6 Although the apoptotic machinery is present in Eos, 7 it remains to be further elucidated whether it is compromised in allergic diseases.
Published data indicate that survivin is associated with defects in apoptosis. 8 Survivin belongs to a family of apoptosis inhibitors. Most studies focus on its role in the pathogenesis of cancer. 9 Other cells also express survivin, such as epithelial cells, 10 dendritic cells, 11 and lymphocytes. 12 Survivin is also expressed by Eos which contribute to the pathogenesis of allergic diseases. 13 Thus, we hypothesized that the apoptosis machinery in Eos in an allergic environment may be compromised by survivin. To test the hypothesis, we isolated Eos from mice suffering from FA, and their apoptosis status was assessed. The results of this study showed that survivin expression was significantly higher in Eos of FA mice than in those of naïve control mice. Inhibition of survivin resulted to the induction of apoptosis.
Materials and methods
Reagents
The Abs against survivin, Fas, FasL, MBP, IL-4 receptor α (IL-4Rα), c-Myc, and IL-4α RNAi kit were purchased from Santa Cruz Biotech (Santa Cruz, CA). The YM-155, survivin, IL-4, IL-5, IL-13, and survivin ELISA kit were purchased from BioMart (Beijing, China). The annexin v kit, propidium iodide (PI), FITC-dextran, reagents, and materials for immunoprecipitation (IP) and chromatin IP (ChIP) were purchased from Sigma Aldrich (St. Louis, MO). The reagents and materials for RT-qPCR and Western blotting were purchased from Invitrogen (Carlsbad, CA).
Mice
Male BALB/c mice (6–8 wk old) were purchased from the Guangzhou Experimental Animal Center (Guangzhou, China). The mice were maintained in a specific pathogen-free facility at Shenzhen University and allowed to access food and water freely. The present study was approved by the Animal Ethics Committee at Shenzhen University (SZUa20170015).
Establishment of eosinophilia in the mouse intestine
Eosinophilia is one of the pathological signs in FA intestines. Thus, following our established procedures, 14 we developed eosinophilia in the mouse intestine by gavage feeding with OVA (1 mg/mouse) and cholera toxin (20 μg/mouse) in 0.3 mL saline, once a wk for four consecutive wk. The mice were challenged with 5 mg OVA/mouse in wk 5 and sacrificed 1 d after. The FA symptoms and pathological signs were assessed as previously reported. 14 The procedures are referred as “the OVA/CT procedures” hereafter.
Isolation of mouse intestinal Eos
Intestinal segments were excised and cut into small pieces. The samples were incubated with collagenase IV (1 mg/mL) for 2 h at 37°C with mild agitation, and then filtered through a cell strainer (70 μm first, then 40 μm). The lamina propria mononuclear cells (LPMC) were collected by Percoll gradient density centrifugation. Eos were purified from LPMCs, following published procedures, 15 with an Eo enrichment kit (BioMart, Beijing, China) containing anti-CD2 (to deplete T cells), anti-CD14 (to deplete monocytes), anti-CD16 (to deplete neutrophils), anti-CD19 (to deplete B-cells), anti-CD56 (to deplete NK cells), anti-Ly6G (to deplete neutrophils), anti-FcɛRI, and anti-c-kit (to deplete mast cells). The remaining cells were then positively selected by anti-CD35. 16 The purity of Eos was greater than 98% as checked by flow cytometry. The viability of Eos was higher than 99% as checked by Trypan blue exclusion assay.
Cell culture
The isolated Eos were cultured in RPMI1640 medium supplemented with 10% FCS, 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 2 mM glutamine. The medium was changed after 2–3 d. The cell viability was greater than 99% as checked by Trypan blue exclusion assay.
Assessment of apoptosis of Eos
Eos were prepared and activated in the culture as described above. The Eos were then stained with annexin v and PI following the manufacturer’s instructions. Cells were analyzed with a flow cytometer (FACSCanto II, BD Bioscience), and cells stained annexin v+ or PI+ annexin v+ were regarded as apoptotic Eos.
Assessment of Eos in the intestine by flow cytometry
LPMCs were prepared as described above. In surface staining, cells were incubated with Abs (labeled with fluorescence) of interest or isotype IgG at 4°C for 30 min. In intracellular staining, cells were treated with fixative and permeable reagents first, then incubated with Abs (labeled with fluorescence) of interest or isotype IgG at 4°C for 30 min. After washing with PBS thoroughly, cells were analyzed with a flow cytometer (FACSCanto II, BD Bioscience). Data were analyzed utilizing the software package flowjo. Data from isotype IgG staining were used as gating references.
Real-time quantitative RT-PCR (RT-qPCR)
Total RNA was extracted from Eos and converted to cDNA with a reverse transcription kit following the manufacturer’s instructions. The samples were amplified in a qPCR device with the SYBR Green Master Mix and the presence of relevant primers, including survivin (tggcagctgtacctcaagaa and tctatcgggttgtcatcggg), Fas (acctccagtcgtgaaaccat and ctcagctgtgtcttggatgc), and FasL (atagccaaccccagtacacc and gctggttgttgcaagactga). Results are presented as fold change against the housekeeping gene β-actin.
Preparation of protein extracts from Eos
Eos were collected from relevant experiments and lysed with a lysing buffer. The lysates were centrifuged for 10 min at 10,000 g. Supernatants were collected and used as cytosolic extracts. Pellets were incubated with a nuclear lysing buffer for 30 min and centrifuged at 10,000 g for 10 min, and supernatants were collected and used as nuclear extracts. All procedures were performed at 4°C.
Western blotting
Protein extracts were fractioned by SDS-PAGE and transferred onto a PVDF membrane. After blocking with 5% skim milk for 30 min, the membrane was incubated with the primary Abs (1:500 diluted) of interest or isotype IgG overnight (16–18 h) at 4°C, followed by incubating with the second Abs (labeled with peroxidase; 1:2000 diluted) for 1 h at room temperature (22–24°C). The membrane was washed with TBS Tween 20 after each time of incubation. The immunoblots on the membrane were developed by enhanced chemiluminescence and photographed with an imaging device.
Immunoprecipitation
The protein samples were prepared and precleared by incubating with protein G agarose beads for 2 h. The beads were removed by centrifugation. The supernatant was collected and incubated with Abs of interest or isotype IgG overnight, and the beads were collected by centrifugation. The protein complexes on the beads were eluted with an eluting buffer and analyzed by Western blotting. All procedures were performed at 4°C.
Chromatin IP
Eos were collected from relevant experiments and fixed with 1% formalin for 15 min to crosslink the DNA and surrounding proteins. The cells were lysed with a lysing buffer, and lysates were sonicated to shear the DNA into small pieces. The samples were then subjected to the IP procedures. After elution from beads, the DNA was recovered from the samples with a reagent kit following the manufacturer’s instructions. The DNA was analyzed by qPCR in the presence of primers of the FasL promoter (aggcagagtggtcggtttta and cctatccatcccacttcccc). Results are presented as fold change against the input.
ELISA
Cytokine levels were determined by ELISA using commercial reagent kits following the manufacturer’s instructions. For assessing the serum levels of OVA-specific IgE, plates were coated with OVA (20 µg/mL; at 4°C overnight; BSA was used as a negative control). After blocking with isotype IgG for 30 min, serum samples (100 ml/well) or IgE standard (double dilution from 100 ng/mL to 0 ng/mL) were added to the plates and incubated at 4°C overnight. The plates were washed with TBS (containing 0.1% Tween 20) three times and incubated with a peroxidase-labeled second Ab for 1 h at room temperature. The plates were washed three times again. Tetramethylbenzidine (TMB) was added to each well to develop the immune reaction products; the reaction was terminated by adding 2 M H2SO4. The plates were read with a microplate reader, and readout values from negative control wells were subtracted from those of each sample well. A standard curve was generated first. Results were calculated applying a standard curve.
Immunohistochemistry
Jejunal segments were excised from mice after relevant treatment. The samples were snap frozen in liquid nitrogen. Cryosections were prepared and dried at room temperature overnight. After blocking with 1% BSA for 30 min, the sections were incubated with the primary Abs (1:200 diluted) of interest or isotype IgG overnight at 4°C, followed by incubating with the second Abs (labeled with fluorescence; 1:300 diluted) for 2 h at room temperature. The sections were washed with PBS three times after each time of incubation. The nuclei in sections were stained with PI to show the tissue structure. The slides were mounted with cover slips and observed with a fluorescent microscope. The sections were coded; the observers were not aware of the code to avoid the observer bias.
Histology
Jejunal segments were excised from mice and fixed with 4% formalin overnight. The tissue was embedded in paraffin following the routine procedures. Paraffin sections were prepared (4 µm) and stained with hematoxylin and eosin. The sections were observed with a light microscope.
RNA interference (RNAi)
RNAi reagent kits of target genes were purchased from Santa Cruz Biotech. Eos were treated with the reagent kits following the manufacturer’s instructions. The effects of RNAi were assessed by Western blotting 48 h after RNAi.
Inhibition of survivin in mice
Mice were received YM155 (an inhibitor of survivin; 2 mg/kg/d in 0.3 mL saline) by gavage-feeding concurrently with the administration of OVA/CT in the course of sensitization. The control mice were fed with saline instead of YM155.
Statistics
The data are presented as mean ± SEM. The difference between two groups was determined by Student’s t test or ANOVA followed by Dunnett’s t test or Student–Newman–Keuls test if more than two groups. P < 0.05 was considered statistically significant.
Results
Defects in apoptosis in Eos in the intestine of mice with FA
To elucidate if defects in apoptosis in Eos contribute to eosinophilia in the intestine, eosinophilia was established in the mouse intestine of an FA mouse model applying OVA/CT procedures. The mice showed FA-like response, including higher levels of OVA-specific IgE, MBP, and IL-4 in the sera (Figure 1a to c). LPMCs were prepared from the intestinal tissues of FA and control mice and analyzed by flow cytometry. The results showed that the frequency of Eos was much higher in the FA group than in the control group (Figure 1d and e). In addition, Eos were negatively selected from LPMCs by MACS. The Eos were stained with annexin v and PI and analyzed with a flow cytometer. The results showed that about 1% Eos were apoptotic in both, FA mice and naive mice. After exposure to cisplatin, an apoptosis inducer, the frequency of apoptotic Eos was significantly increased in the control group, while the number of apoptotic Eos in the FA group was only slightly increased (Figure 1f and g). The results indicated that apoptosis can be induced in Eos under physiological conditions, which is significantly compromised in the allergic environment.
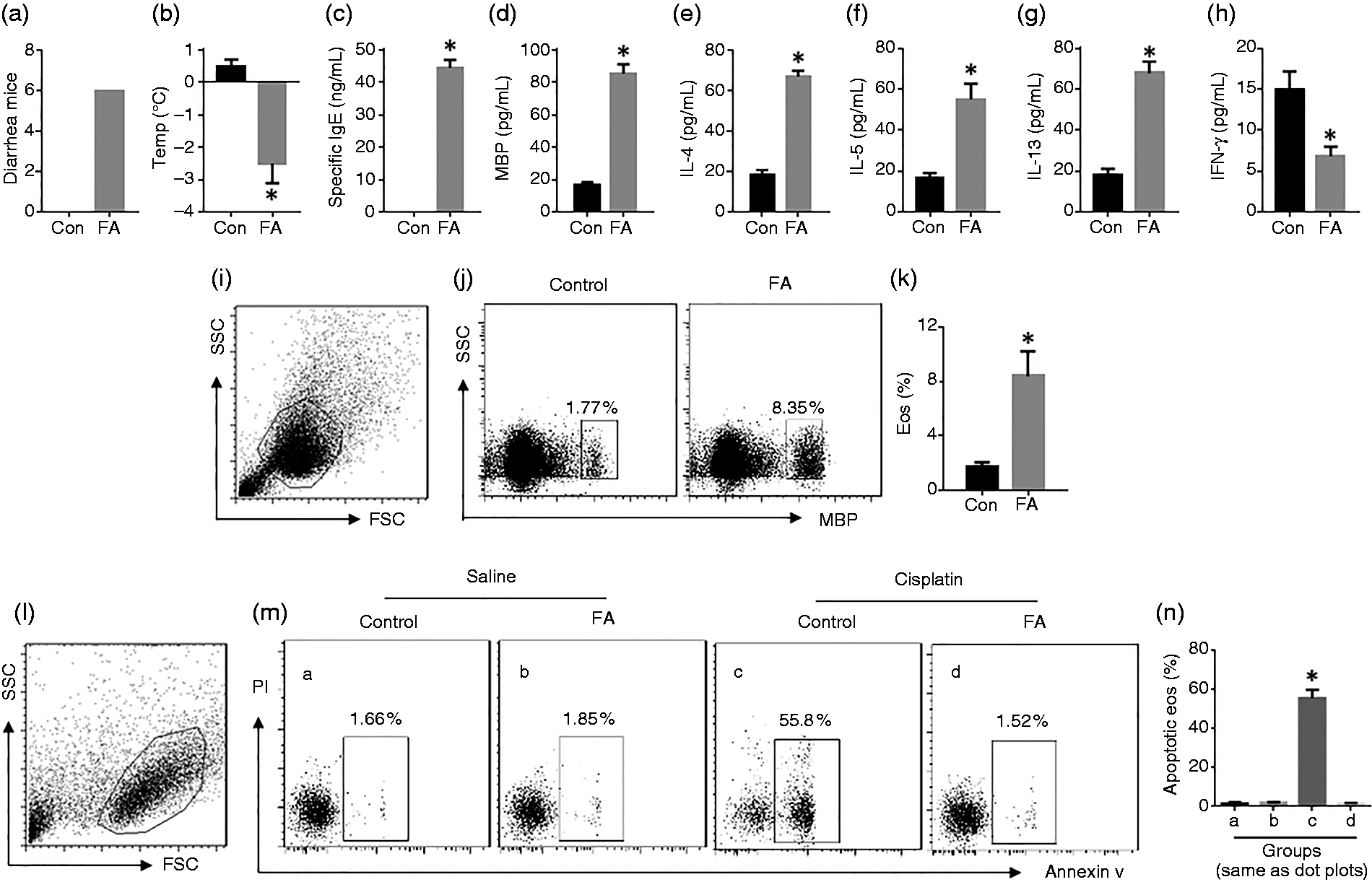
Induction of apoptosis in intestinal Eos. BALB/c mice were treated with the OVA/CT procedures to develop FA. (a) The number of mice that had diarrhea during 2 h after OVA challenge. (b) Core temperature changes 30 min after OVA challenge. Serum levels of specific IgE (c), MBP (d), Th2 cytokines (e–g), and IFN-γ (h). (i) Plots of FSC and SSC for Eo gating in (j). (j) Gated dot plots show Eos in LPMCs. (k) Summarized data of Eos in LPMCs. (l) Plots of FSC and SSC for Eo gating in (m). (n) Summarized data of the gated dot plots in (m). The data indicated by the bars are presented as mean ± SEM. *P < 0.01, compared with the control (Con) group. Each group consists of 10 mice.
Survivin is positively correlated with the frequency of Eos in the intestine
Survivin is an inhibitor of apoptosis. 8 To elucidate if survivin is associated with the defects in apoptosis in Eos of FA mice, the jejunal segment was excised and processed for cryosections. The sections were stained by immunohistochemistry and Eos were counted under a light microscope. The frequency of Eos was significantly higher in FA mice than in control mice (Figure 2a and b). Protein extracts were prepared from intestinal tissue and analyzed by ELISA. The results showed that protein levels of survivin were higher in the FA group than in the control group (Figure 2c). A correlation assay was performed with the data of survivin and Eo frequency. A positive correlation was identified between the expression of survivin and the Eo frequency in the intestine (Figure 2d). The results imply that survivin may be associated with eosinophilia in the intestine of FA mice. Next, we isolated Eos from the intestine. Survivin protein was also detected in Eos, which was significantly higher in Eos of the FA group than in the control group (Figure 2e). However, the mRNA of survivin was merely detected in Eos of both, the FA group and the control group (Figure 2f). The results suggested that Eos do not actively express survivin. The survivin detected in FA Eos (Figure 2e) may be absorbed from that released by epithelial cells. To test the inference, Eos were isolated from the naïve control mouse intestine and cultured in the presence of exogenous recombinant survivin (labeled with FITC). As observed with a confocal microscope, Eos were capable of absorbing exogenous survivin (Figure 2g). To localize the sources of survivin in the intestine, immunohistochemistry was performed with the intestinal sections. The results showed that intestinal epithelial cells were the main source of survivin in the intestine (Figure 2h).
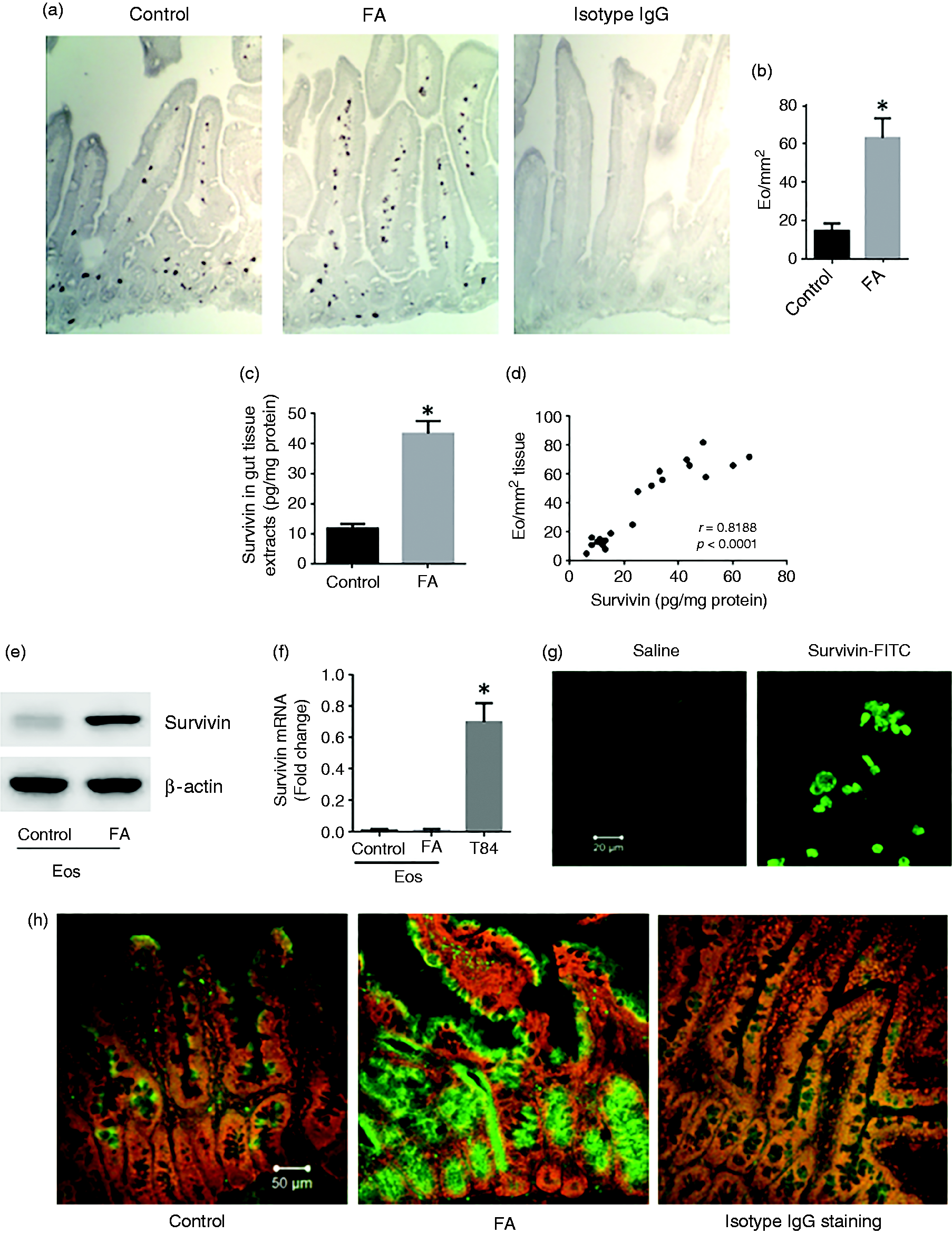
Survivin positively correlates with Eo number in the intestine. BALB/c mice were treated with saline (control; n = 10) or OVA/CT (to develop intestinal eosinophilia; n = 10). (a) Representative immunohistochemistry images (×200) show Eos (stained in dark brown) in the intestinal tissue. (b) Summarized data of (a). (c) Protein levels of survivin in intestinal tissue. (d) The scatter dot plots show correlation between Eo frequency and survivin in intestinal mucosa. (e, f) Eos were isolated from the intestine. The immunoblots show the levels of survivin in Eos (e); the bars show the levels of survivin mRNA in Eos (f). The samples of T84 cells were used as a positive control. (g) The confocal images show Eos absorbed recombinant survivin (labeled with FITC) in the culture. (h) The green stain on the representative confocal images show the positive stain of survivin in the intestinal sections (by immunohistochemistry; ×200). The data of bars are presented as mean ± SEM. *P < 0.01 (t test), compared with control group.
Th2 cytokines induce expression of survivin in intestinal epithelial cells
To elucidate the role of allergic condition on the expression of survivin in epithelial cells, IEC cells, an intestinal epithelial cell line, were exposed to Th2 cytokines, including IL-4, IL-5, and IL-13, for 48 h. The exposure to IL-4 or IL-13, but not IL-5, markedly increased the expression of survivin in IEC cells (Figure 3a and b). The results implied that IL-4 and IL-13 activate the IL-4 receptor (IL-4R)α on IEC cells to induce the expression of survivin. To test the inference, IEC cells were treated with IL-4Rα shRNA to knock down the expression of IL-4Rα (Figure 3c). The IL-4Rα-deficient IEC cells were exposed to IL-4 in the culture. Indeed, the expression of survivin was not increased in these cells. To corroborate the results IEC cells were treated with IL-4 at gradient concentrations in the culture. The expression of survivin by IEC cells was induced in an IL-4 concentration-dependent manner as shown in the IEC cell extracts and in culture supernatant (Figure 3d to f). The results demonstrated that IL-4 or/and IL-13 activated IL-4Rα to induce the expression of survivin in intestinal epithelial cells; the survivin can be released into the microenvironment.
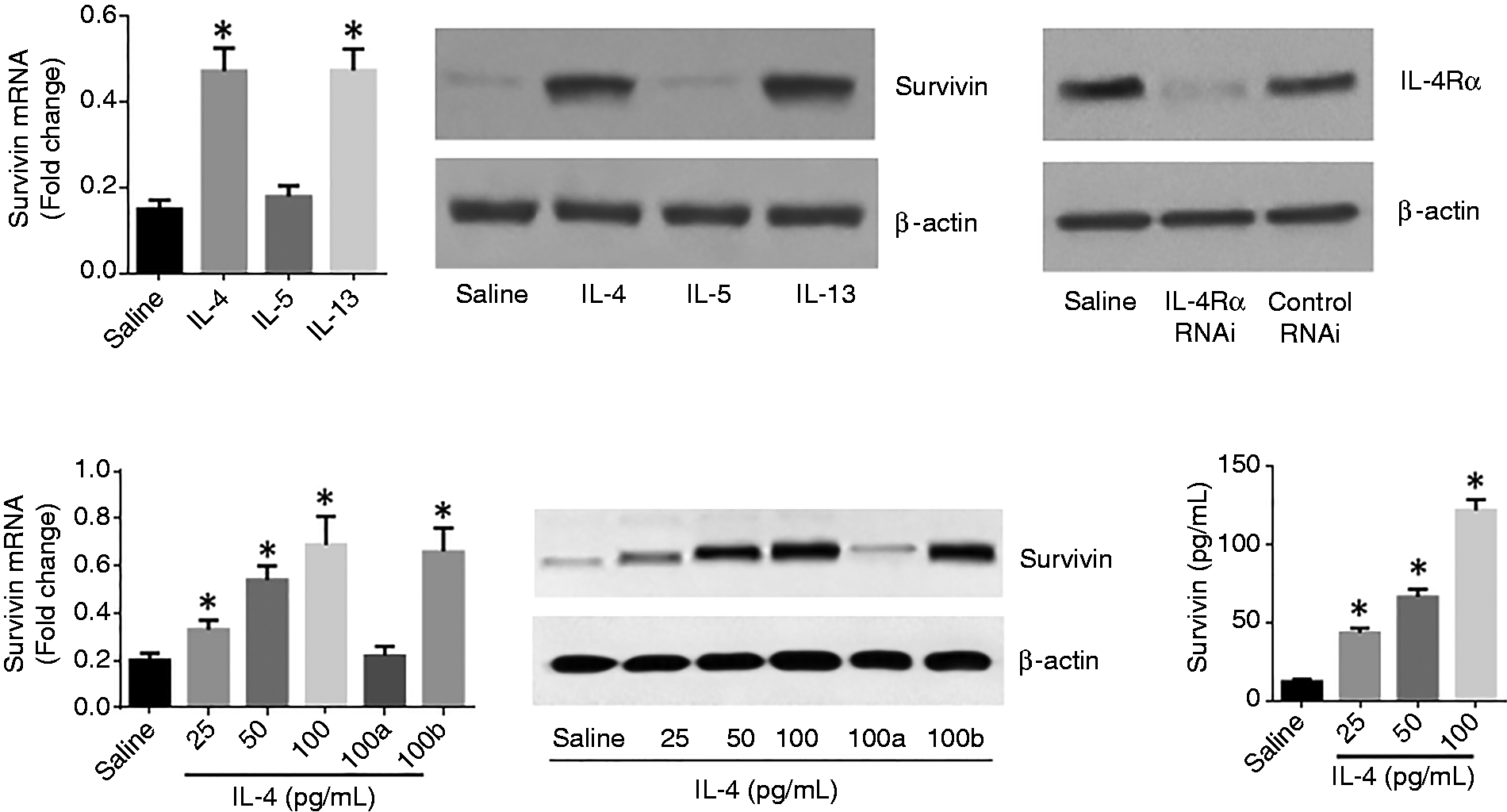
Th2 cytokines induce survivin expression by IEC cells. (a, b) IEC cells were exposed to reagents (100 pg/mL for each cytokine) as denoted on the x-axis of (a) for 48 h. (c) The results of IL-4Rα RNAi. (d, e) IEC cells were exposed to survivin at gradient concentrations in the culture for 48 h. The bars of (a) and (d) show the mRNA levels of survivin. The immunoblots in (b) and (e) show the protein levels of survivin. (f) The levels of survivin in the culture supernatant (by ELISA). *P < 0.01, compared with the saline group (t test for (a); ANOVA for (d) and (f)). (a) IEC cells were treated with IL-4Rα RNAi to knock down the expression of IL-4Rα. (b) IEC cells were treated with control RNAi used as controls.
Survivin suppresses gene transcription of FasL in Eos
Since Fas and FasL play a central role in the induction of apoptosis, the expression of Fas and FasL in Eos was assessed. The results showed that the levels of Fas in Eos were not disturbed by the activation (Figure 4a and b). The expression of FasL in Eos was markedly increased in the saline group after activation, which did not occur in the FA group (Figure 4c and d). The results suggested that survivin may disturb the expression of FasL in Eos. To test this, Eos were isolated from the intestine of naïve mice. The Eos were cultured in the presence of survivin and activators for 48 h. Indeed, exposure to survivin suppressed the expression of FasL in Eos in a dose-dependent manner (Figure 4e and f). Activation of Eos by cisplatin did not alter the expression of p53 (Figure 4g and h). By co-IP, a complex of survivin and c-Myc, the transcription factor of FasL, was detected in the cell extracts of Eos isolated from the FA group (Figure 4i). The results implied that the epithelial cell-derived survivin can be absorbed by Eos and forms a complex with c-Myc in Eos. To test such inference, a Flag-c-Myc-expressing plasmid was constructed and transfected into EoL-1 cells (Figure 4j). The cells were then cultured in the presence of recombinant survivin (with a His label) for 12 h and analyzed by co-IP. A complex of survivin and recombinant c-Myc was detected in the cell extracts (Figure 4k). To understand the physiological role of the physical contact between survivin and c-Myc, a ChIP assay was performed with the cell extracts. The gene transcription activities, including the levels of c-Myc and Pol II (RNA polymerase II) at the FasL promoter locus was lower in Eos collected from FA mice as compared to the control mice (Figure 4l and m). The results indicated that survivin physically contacted c-Myc to restrict the c-Myc to bind the FasL promoter, thus, to restrict FasL gene transcription in Eos. On the other hand, exposure to exogenous survivin in the culture suppressed the expression and induced defects of apoptosis in naive Eos (Figure 5).
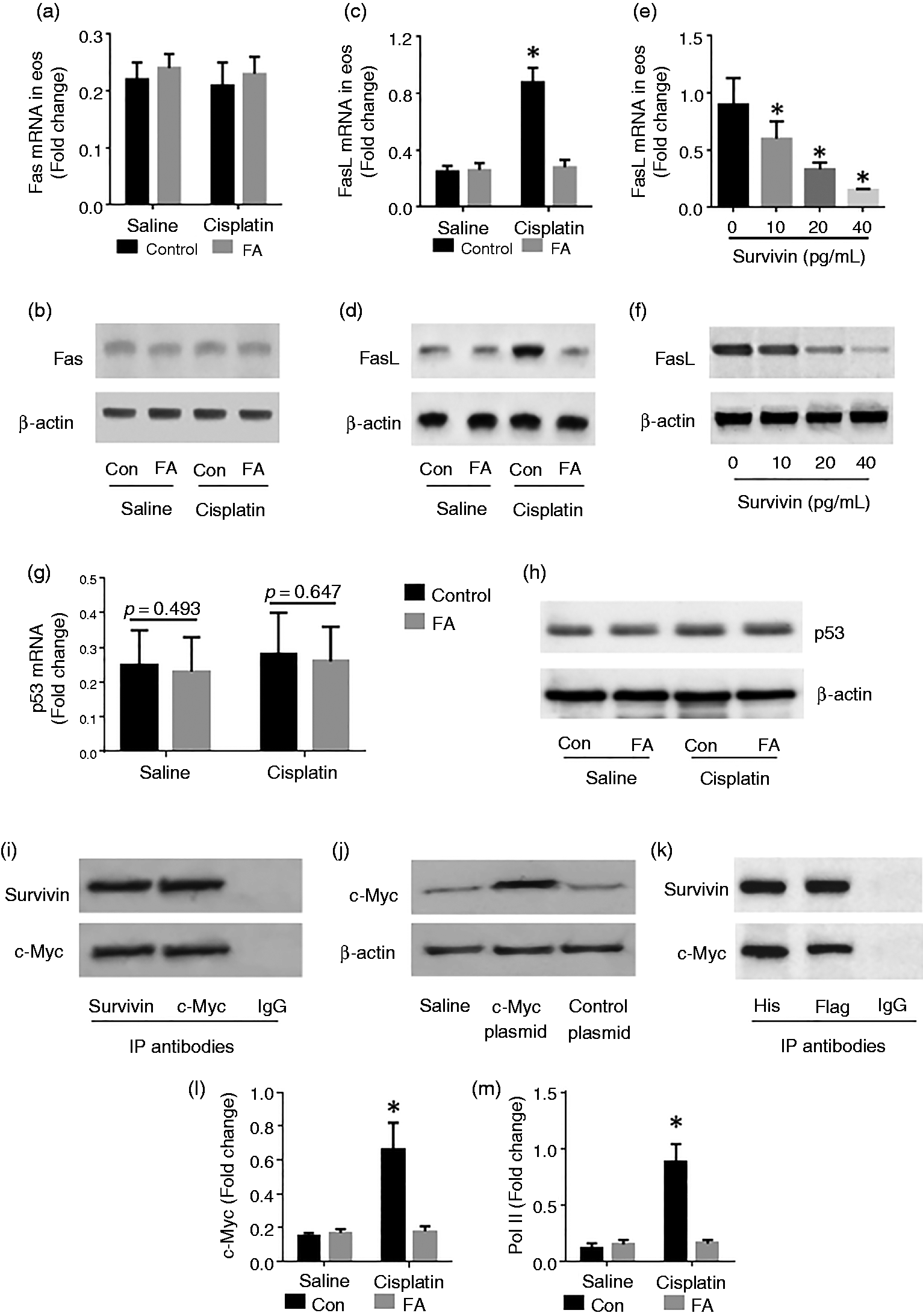
Assessment of Fas and FasL in intestinal Eos. (a–f) LPMCs were prepared from naive control (Con) mice (n = 10) and FA mice (n = 10). Eos were purified from LPMCs by MACS and exposed to cisplatin (25 µM) for 48 h. The Eo extracts were analyzed by RT-qPCR and Western blotting. The bars indicate the mRNA levels of Fas and FasL; the immunoblots indicate the protein levels of Fas and FasL. The data of bars are presented as mean ± SEM. *P < 0.01, compared with the saline group. (g, h) The expression of p53 in Eos. (i) Eos were treated with cisplatin in the culture. The immunoblots show a complex of c-Myc and survivin in Eos. (j) c-Myc-expressing (labeled with Flag) plasmids were transfected into EoL-1 cells (an Eo cell line). The immunoblots show the expression of c-Myc in the EoL-1 cells. (k) EoL-1 cells with c-Myc expression were exposed to recombinant survivin (labeled with His) in the culture for 12 h. The immunoblots show that a complex of c-Myc and survivin formed in EoL-1 cells. (l, m) The levels of c-Myc (L) and Pol II (M) at the FasL promoter locus of Eos after activation. The data represent three independent experiments.
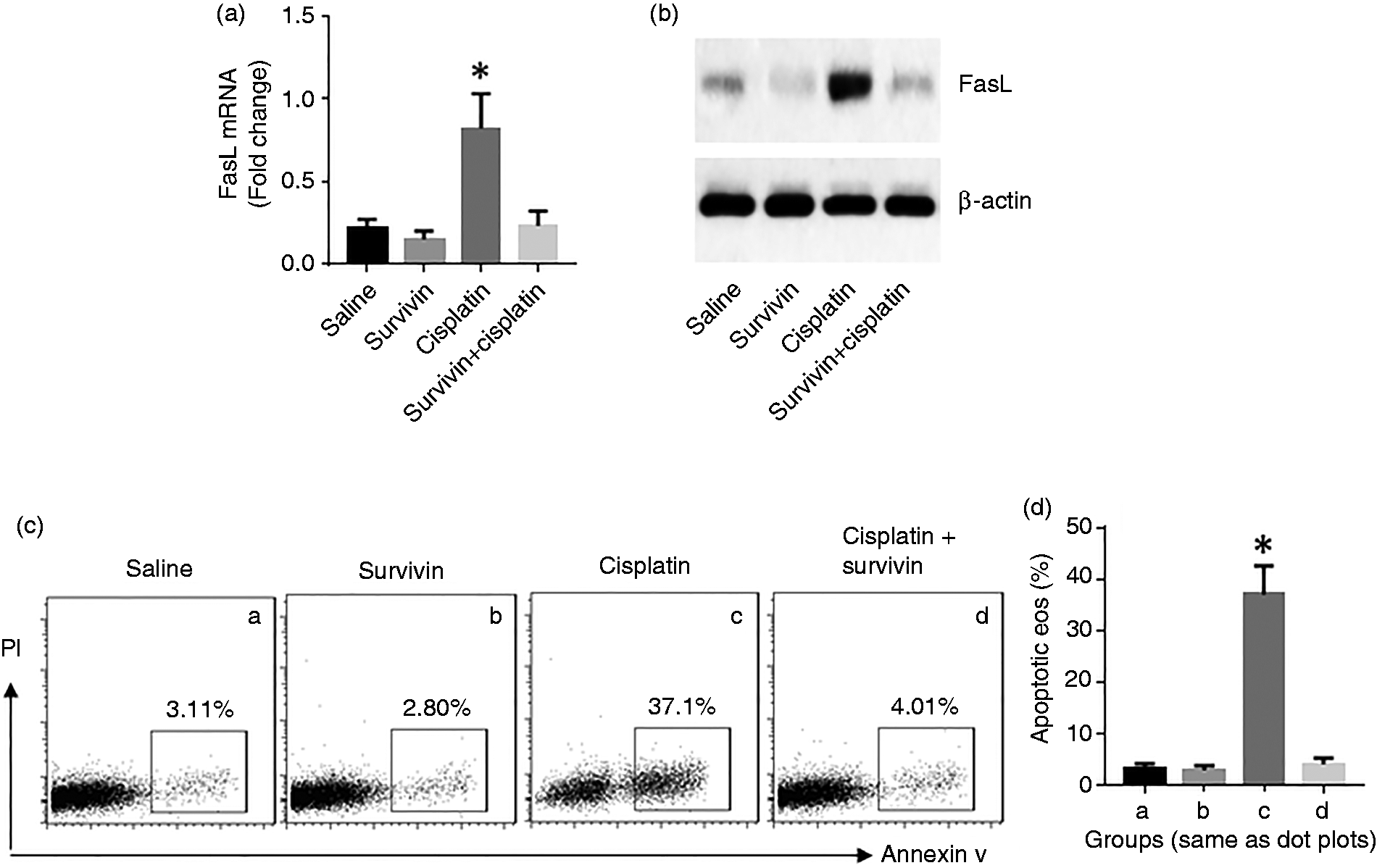
Exogenous survivin suppresses apoptosis in Eos. Eos were isolated from LPMC of naïve mice and cultured with the conditions denoted on the x-axis of (a). (a, b) Expression of FasL in Eos. (c) Gated dot plots indicate apoptotic Eos. (d) Summarized data of (c). Data of bars are presented as mean ± SEM. *P < 0.01, compared with the saline group. The data represent three independent experiments. Cisplatin = 25 µM. Survivin = 40 pg/mL.
Inhibition of survivin attenuates Eo-related intestinal inflammation in mice
The data reported above demonstrated that survivin plays an important role in the pathogenesis of eosinophilia and that inhibition of survivin may attenuate the Eo-related intestinal inflammation. To test this, we established eosinophilia in the intestinal mucosa of mice following the FA-inducing procedures with or without administration with YM-155 (an inhibitor of survivin). As compared to control mice, the FA group possessed much more Eos in the intestinal mucosa, indicating that the eosinophilia was successfully established. The number of mononuclear cells in the intestinal mucosa was also markedly higher in the FA group than that in the control group. The epithelial permeability was significantly higher in the FA group than that in the control group. Intestinal tissue destruction was observed in the FA group, but not in the control group. The data demonstrated that inflammation was induced in the intestinal mucosa, which was significantly attenuated in those FA mice that had received YM-155 (Figure 6). These results demonstrated that inhibition of survivin can attenuate eosinophilia and the related inflammation in the intestine.
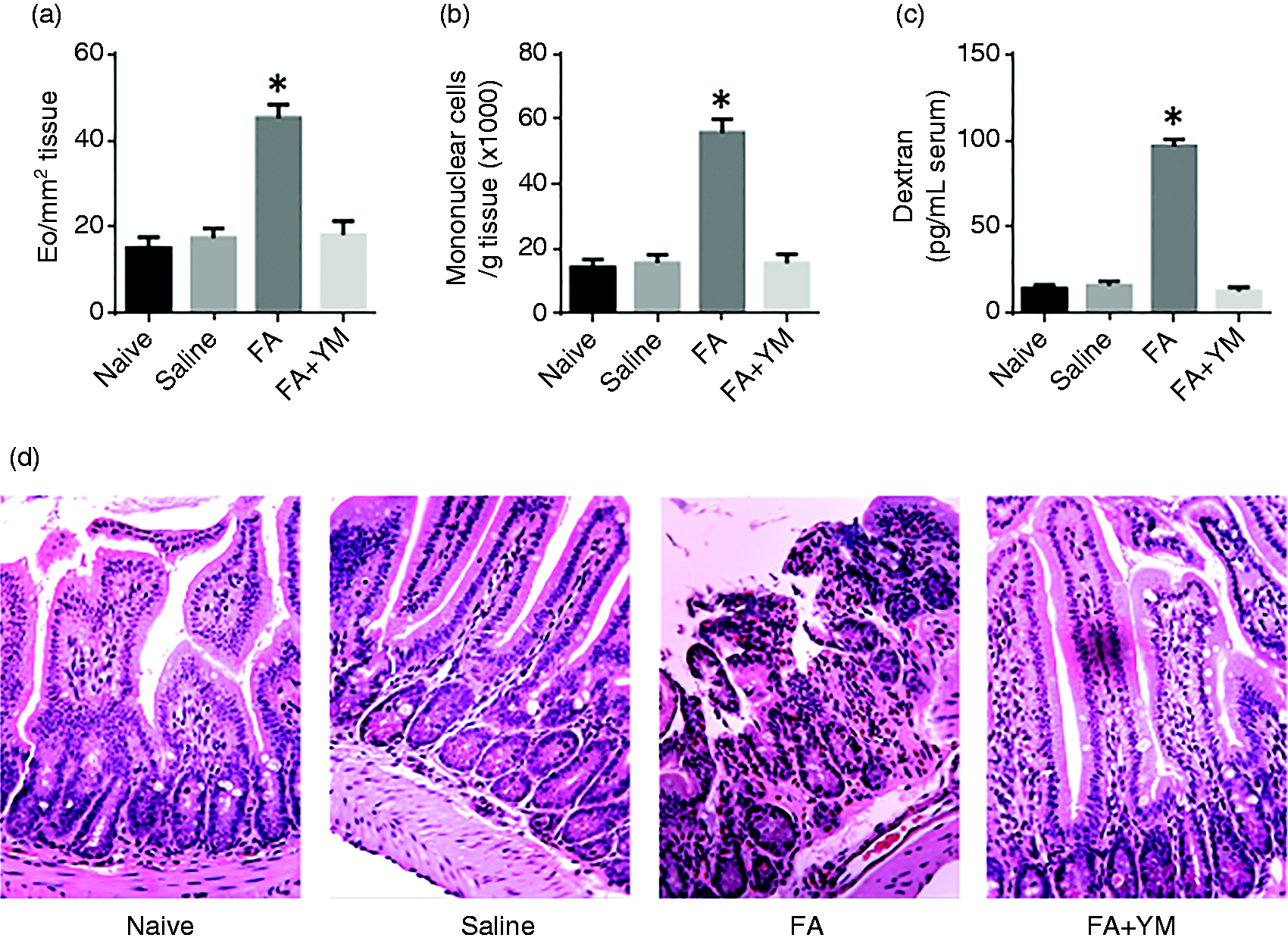
Inhibition of survivin inhibits eosinophilia-related inflammation in the intestine. BALB/c mice were treated with the OVA/CT procedures to establish eosinophilia in the intestine. (a) Frequency of Eos in the intestine; the Eos in immunohistochemistry stained sections were counted in 20 randomly selected fields (×400) of each sample. (b) Mononuclear cell number per gram tissue processed by flow cytometry. (c) Serum levels of dextran used as an indicator of intestinal epithelial barrier permeability. (d) Representative images (×200) showing histology of the small intestine of mice. Naïve: naïve mice. Saline: mice were gavage-fed with saline used as controls. FA: mice were treated with the OVA/CT procedures to establish the FA condition and eosinophilia in the intestine. YM: YM-155 (an inhibitor of survivin). Each group consisted of 10 mice. The data indicated by the bars are presented as mean ± SEM. *P < 0.01 (t test), compared with the naïve group.
Discussion
The presented data demonstrated that intestinal epithelial cells produce survivin, which can be up-regulated by Th2 cytokines. The survivin can be released into the intestinal tissue to be absorbed by Eos. The survivin binds to c-Myc to form a complex to restrict the c-Myc from binding the FasL promoter in Eos, thus, to restrict the expression of FasL and induce the defects in apoptosis in Eos. Eos with defects in apoptosis are resistant to the induction of apoptosis, which may be a causative factor in the development of eosinophilia.
Eosinophilia is characteristic for various inflammatory disorders, including allergic reactions, parasite infections and some chronic inflammatory diseases such as eosinophilic esophagitis, eosinophilic enteropathy, or eosinophilic myocarditis. 17 Based on released toxic mediators upon Eo activation, eosinophilia contributes a lot to the pathogenesis of such inflammatory diseases. It is recognized that in allergic responses, parasite infections are the causative factors of eosinophilia. 17 IL-5, one of the signature Th2 cytokines, is required for the development of Eos, 18 and IL-33 can promote the Eo development. 19 The presented data provided a novel factor contributing to eosinophilia. The data showed defects in apoptosis in Eos that prevents the apoptosis induction in Eos isolated from the FA mouse intestine, suggesting that the defects in apoptosis in Eos contributes to the eosinophilia in the intestine of FA mice, which was demonstrated by the results that administration of survivin inhibitor blocked the induction of eosinophilia and eosinophilic inflammation in the intestine as shown by the present data.
We observed the defects in apoptosis in Eos isolated from the intestine of FA mice that had eosinophilia. The data show that apoptosis can be induced in Eos isolated from naïve mice, suggesting apoptosis can be induced in Eos under physiological condition. The data also show that apoptosis cannot be induced in Eos isolated from FA mice, suggesting that the apoptosis machinery is compromised in these Eos. The role of apoptosis is to eliminate the unwanted cells to maintain the homeostasis in the body. 20 The defects in apoptosis may result in certain cell type over accumulation in the body to induce disorders such as inflammation in the body. 21
In this study, we observed that survivin was over-expressed in the intestine of FA mice. Survivin is an inhibitor of apoptosis. The data show that survivin plays a role in the defects in apoptosis in Eos. We did observe survivin in Eos isolated from the FA mouse intestine, but those Eos did not show the mRNA of survivin, indicating that these Eos do not express survivin. This is in line with the notion that survivin is not produced by terminally differentiated cell types. 22 Eos are a terminally differentiated cell type; the cells should not express survivin. The survivin detected in Eos can be absorbed from external sources. The data revealed such an external source that intestinal epithelial cells produce survivin, which was up-regulated by Th2 cytokines.
Most studies on survivin are about its role in the pathogenesis of cancer. Survivin inhibits caspase 9, 23 or/and caspase 3 and caspase 7, 24 or interferes with the function of p53, 23 to abrogate the process of apoptosis, so as to promote cancer cell growth. Recent reports indicate that survivin is also involved in the pathogenesis of other disorders; survivin transgenic mice show the tendency to suffer from Th2-biased inflammation such as airway allergy. 25 Survivin is also involved in the pathogenesis of rheumatoid arthritis by promoting joint damage and resulting in poor treatment response. Inhibition of survivin attenuates the inflammation of rheumatoid arthritis in mice. 26 Our data are in line with the recent reports by showing that survivin is associated with the pathogenesis of experimental immune inflammation in the intestine.
Eosinophilia is a sign of a number of immune inflammations. Because of releasing toxic mediators upon activation, Eos directly contribute to the development of inflammation in the intestine. 27 Current therapeutic remedies for the eosinophilia-related inflammation in the intestine are not satisfactory. 27 In the present study, we induced eosinophilia and intestinal inflammation in mice. Importantly, both eosinophilia and inflammation in the intestine were significantly attenuated by the inhibition of survivin. The results mirror the importance of the role of survivin in the pathogenesis of eosinophilia-related disorders in the intestine, which may be a novel therapeutic remedy for the treatment of the eosinophilia-related inflammation in the intestine.
In summary, intestinal epithelial cells produce survivin in response to Th2 cytokine stimulation. The survivin can be absorbed by Eos to interfere with the apoptotic machinery to compromise the apoptotic machinery in Eos. Inhibition of survivin can attenuate the eosinophilia-related inflammation in the intestine. The data suggest that survivin may be a novel target for the treatment of FA.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National “13th-5” key program, Precision medicine research project (grant numbers 2016YFC0905802 and 2016YFC0903700), Guangdong provincial scientific technological research project (grant number 2016A020216029), and Shenzhen scientific technological basic research project (grant numbers JCYJ20160429114659119, JCYJ20160328144536436, and KQTD20170331145453160).