
Abstract
Olanzapine (OLZ) is an atypical antipsychotic drug that also has mood-stabilizing effects. The mechanism of action of OLZ is not fully understood. Accumulating data suggest that inflammation plays a role in the pathophysiology of mental disorders and that psychotropic drugs exhibit some anti-inflammatory properties. This study was undertaken to examine the effects of OLZ on LPS-induced inflammation in rat primary glia cells. Glia cells were extracted from newborn rat brains. OLZ (1 or 50 µM) was added to culture medium at 6 or 72 h before addition of LPS for another 18 h, and levels of IL-10, prostaglandin (PG) E2, NO and TNF-α, and expression of cyclo-oxygensase (COX)-2 and inducible NO synthase (iNOS) were determined. Treatment with 50 µM OLZ (but not 1 µM) significantly decreased LPS-induced secretion of IL-10, PGE2 and TNF-α. In contrast, 50 µM OLZ significantly increased NO levels. OLZ did not alter the expression of COX-2 or iNOS in LPS-treated cells. These results suggest that OLZ differently affects the secretion of inflammatory mediators. Most of the significant effects of OLZ were obtained when 50 µM was used, which is a high and probably therapeutically irrelevant concentration. Therefore, under the conditions used in the present study OLZ seemed to lack a potent anti-inflammatory effect.
Introduction
Antipsychotic medications are the core treatment of schizophrenia,1,2 but they are also used in the treatment of bipolar affective disorder.3,4 The blockade of dopamine D2 receptors in the brain is a unifying pharmacological property of all effective antipsychotic drugs,1,2,5 suggesting that schizophrenia may be a disease of dopaminergic over-activity. This assumption is supported by the fact that currently there are no effective antipsychotic drugs that are not dopamine (D2) receptor antagonists, i.e. drugs that act through a non-dopamine blocking mechanism. Olanzapine (OLZ) is a second-generation antipsychotic drug that is beneficial in the treatment of schizophrenia and bipolar disorder.2–4,6 Similar to other psychotropic medications, OLZ produces numerous undesirable side effects.2,4,6 Among the most troubling side effects of OLZ are mass gain and increases in measures of Glc (diabetes) and lipid metabolism.2,4,6
Accumulating evidence suggests that inflammation may contribute to the pathophysiology of several mental illnesses, including schizophrenia,7–11 bipolar disorder12,13 and depression.14–19 Moreover, a large body of data implies that inhibition of inflammation may contribute to the therapeutic effects of psychotropic drugs.20–37 However, it is important to emphasize that many studies reported opposite findings; namely, that psychotropic drugs also exhibit pro-inflammatory effects.10,31,38–41 Furthermore, the mechanism underlying the anti-inflammatory properties of psychotropic drugs is not fully understood and it is not known whether their influence on inflammation contributes to their therapeutic and toxic effects.
Neuro-inflammation occurs as a response to pathologic processes in the brain such as infection, injury, degeneration or ischemia. Inflammation allows the killing of invading organisms and phagocytosis of damaged cells. 42 An acute inflammatory response to an insult in the brain differs dramatically from that of other tissues. The parenchyma of the brain is protected by both the cerebrospinal fluid and the blood–brain barrier against diffusion of molecules and microorganisms from the blood stream to the brain. Those barriers also block immune cells entry into the central nervous system (CNS). 43 Fast recruitment of neutrophils is nearly absent and monocytes are only recruited after several days. These phagocytes kill invading microorganisms, remove cell debris and facilitate tissue repair and return to homeostasis.
Brain tissue is composed of two types of cells: neurons and glia cells. Among glia cells there are three types: oligodendrocytes, microglia and astrocytes.44–46 Oligodendrocytes produce myelin, which covers neuron axons and facilitates the conduction of electrical impulses.44–46 Microglia are the brain equivalent of macrophages.44–46 They repair tissue damage, fight invading microorganisms and help in the maintenance of brain homeostasis by removing toxic debris and dead cells. Astrocytes support neurons and allow them to function properly.45,47 Astrocytes also regulate and maintain the function of synapses, which guarantees rapid transfer of information from neurons to their target cells.48,49 Microglia and astrocytes activate the inflammatory response by secreting pro-inflammatory mediators such as IL-1β, IL-6, NO, prostaglandin (PG) E2 and TNF-α.28,45,46,48,50,51 Alternatively, microglia and astrocytes can suppress the immune system by producing anti-inflammatory mediators such as IL-10.44–46,50 These data suggest that glia cells are very relevant to the research of inflammation-associated neurological disorders.
Despite their important role in maintaining brain homeostasis, over-activity of glia cells can exacerbate brain pathologies rather than being protective.45,46,52,53 The dual role of glia cells during inflammation is unique but not necessarily perfectly balanced. The balance in the contrasting effects of glia cells (pro- vs. anti-inflammatory) determines the nature of inflammation development. An excessive immune response or overactive glia cells result in neurodegenerative and other neurological disorders. 54 For example, activated microglia cells may produce an oxidative burst involving the induction of enzymes/complexes such as NADPH oxidase, inducible NO synthase (iNOS) and sometimes myeloperoxidase. These enzymes/complexes generate superoxide (O2–), NO and hypochlorous acid, respectively. 55 Microglial activation results in elevated expression of iNOS and the synthesis of NO, which contributes to pathological inflammatory processes in the brain.56–59 Another inflammatory pathway that is frequently linked to the pathophysiology of neurological disorders is the cyclo-oxygenase (COX)–PGs cascade.20,21 PGs such as PGE2 not only have many important physiological functions, but also contribute to a number of pathological processes.60–63 The biosynthesis of PGs involves the release of arachidonic acid (AA) from membrane phospholipids by phospholipase A2 and its conversion to PGs by COX.60–62 The two major isoforms of the enzyme COX are COX-1 and COX-2. COX-1 usually produces PGs that are important for physiological functions, and is expressed constitutively in most tissues.60–62 COX-2 is an inducible enzyme, the up-regulation of which is usually associated with pathological inflammatory and mitogenic processes.60–62 In support of the notion that AA metabolites contribute to the pathology of psychiatric illnesses, several randomized clinical trials have shown that selective COX-2 inhibitors exert therapeutic efficacy.64–72 For example, Müller et al. demonstrated in two double-blind, placebo-controlled trials that the addition of celecoxib (a selective COX-2 inhibitor) to risperidone or amisulpride was beneficial in patients with schizophrenia.64,68 Similarly, in randomized double-blind placebo-controlled trials, Müller et al. and Abbasi et al. showed that celecoxib add-on therapy was useful in patients with major depression.66,71 Moreover, in a double-blind placebo-controlled trial, Nery at al. found that treatment with celecoxib improved affective symptoms of patients with bipolar disorder. 67 Interestingly, aspirin (a non-selective COX inhibitor and a widely used anti-aggregant) has also been shown to possess therapeutic benefits in bipolar disorder.69,70,72
Several studies have examined the effects of OLZ on inflammation. Sugino et al. reported that OLZ exerted potent anti-inflammatory properties as it reduced production of the pro-inflammatory cytokines TNF-α and IL-6 and increased the levels of the anti-inflammatory cytokine IL-10 in the serum of LPS-treated mice. 26 Cheon et al. found that chronic treatment with OLZ significantly decreased COX activity and PGE2 concentration in rat brain. 73 Moreover, OLZ inhibited translocation of the transcription factor NF-κB to the nucleus and decreased IL-1β protein levels in astrocytes. 32 In contrast, other studies have shown contrasting results, i.e. that OLZ has prominent pro-inflammatory effects. For example, Meyer et al. found that treatment with OLZ significantly increased levels of inflammatory markers (particularly C-reactive protein) in schizophrenic patients. 10 Furthermore, chronic OLZ treatment in rats led to a significant increase in white adipose tissue TNF-α level. 38 Similarly, OLZ elevated the levels of IL-1β, monocyte chemoattractant protein-1 and NF-κB in primary human adipose-derived stem cells. 40 Moreover, OLZ enhanced the infiltration of macrophages to adipose tissue, and increased the levels of TNFα, IL-1β and IL-6 in adipose tissue and hypothalamus of female rats. 39 Taken together, these data indicate that the evidence regarding the effects of OLZ on inflammation is not conclusive. It is thus important to understand the influence of OLZ on brain inflammation. The present study was undertaken to examine the effects of OLZ on LPS-induced inflammation in rat primary glia cells.
Materials and methods
Primary rat glia culture
Primary cultures of mixed glia cells were extracted from Wistar newborn (0–24 h) rat brains. The procedures were used in accordance with the guidelines of the Faculty of Health Sciences Committee on the Use and Care of Laboratory Animals (Authorization # IL-50-10-2010). Glia cell extraction was performed as previously described.28,74 Briefly, brains were removed under sterile conditions and washed three times with a sterile 0.9% NaCl solution. Then, meninges and blood vessels were removed. The cells were dissociated through a sterile strainer with 280 -µm diameter clefts. Then, the cell suspension was filtered through a sterilized nylon mesh (120-mm pore size) to obtain single cells. The dissociated cells were pelleted twice by centrifugation at 1200 g and washed with saline. The final cell suspension was in a high Glc (4.5%) DMEM containing 10% inactivated FCS, antibiotics (penicillin 100 unit/ml and streptomycin 100 µg/ml), 2 mM l-glutamine and 0.2 U/ml regular insulin. After a cell count with a Coulter counter, 106 cells/ml were seeded in 24-well plates that were coated with 0.01 mg/ml poly-l-lysine. The plates were incubated at 37℃ in an atmosphere of 5% CO2 in air. The medium was replaced after 1–2 d and once a wk thereafter. After 2–3 wk, when the cultures were almost free of neurons and contained mainly glia cells, experiments were conducted. For instance, at this stage, cultures usually contain > 95% glia cells (∼80% astrocytes, ∼15% microglia) and less than 5% neurons, as described previously. 74
Experimental procedure
Escherichia coli LPS (Sigma, St. Louis, MO, USA) was used to induce inflammation in glia cultures. OLZ was used at two concentrations: 1 µM and 50 µM. In preliminary experiments we observed that these concentrations are safe and do not alter cell growth and viability (data not shown). We examined the effects of OLZ on LPS-induced production of IL-10, NO, PGE2 and TNF-α in primary glia cultures. Moreover, we determined the effects of OLZ on expression of the enzymes iNOS and COX-2, which are induced during inflammatory processes in the brain. Cells were treated with OLZ (alone) for 6 h or 72 h. Then, LPS was added to the culture medium for another 18 h. At the end of exposure to LPS medium samples were collected and stored at −20℃ for further determination.
Determination of TNFα, PGE2 and IL-10 levels
Levels of IL-10, PGE2 and TNF-α in samples were determined by specific ELISA kits, according to the manufacturer's protocols (R&D Systems, Minneapolis, MN, USA) and similar to our previous study. 28 The OD of standard and unknown samples was determined by using a spectrophotometer set to 450 nm. The detection limit of IL-10 and TNF-α assays was 62.5 pg/ml. The detection limit of the PGE2 assay was 39 pg/ml. In the samples where the level of the examined inflammatory mediator was below the detection limit, results were expressed as ‘undetectable’ and assigned a result of zero.
Determination of NO levels
Levels of NO in culture medium were determined by measuring the concentrations of nitrite (NO2–), a stable metabolite of NO, using the Griess reaction assay and as previously described.28,75 Samples (100 µl) and standard solution of known concentrations of NaNO2 (0.39–12.5 µM) were transferred into a 96-well plate. These were mixed with an equal volume of Griess reagent (Sigma), which contains 1% sulfanilamide and 0.1% N-1-naphthylethylendiamine dihydrochloride. The plate was incubated in the dark at 20–24℃ for 15 min. The absorbance of the reaction products was measured at 540 nm using a spectrophotometer.
Western blot analysis
Glia cells were grown in small, poly-
Presentation of the data and statistical analysis
Each figure represents one experiment out of several independent experiments in which similar results were obtained. In Figures 1–4, each bar represents 6–18 samples (wells) of the same treatment group. In Figures 5 and 6, each bar represents three independent experiments in which each immunoblot included two flasks of the same treatment group. Results are expressed as mean ± SD, unless otherwise indicated. Statistical evaluation of the results was carried out using a two-tailed student’s t-test. Values of P < 0.05 were considered statistically significant.
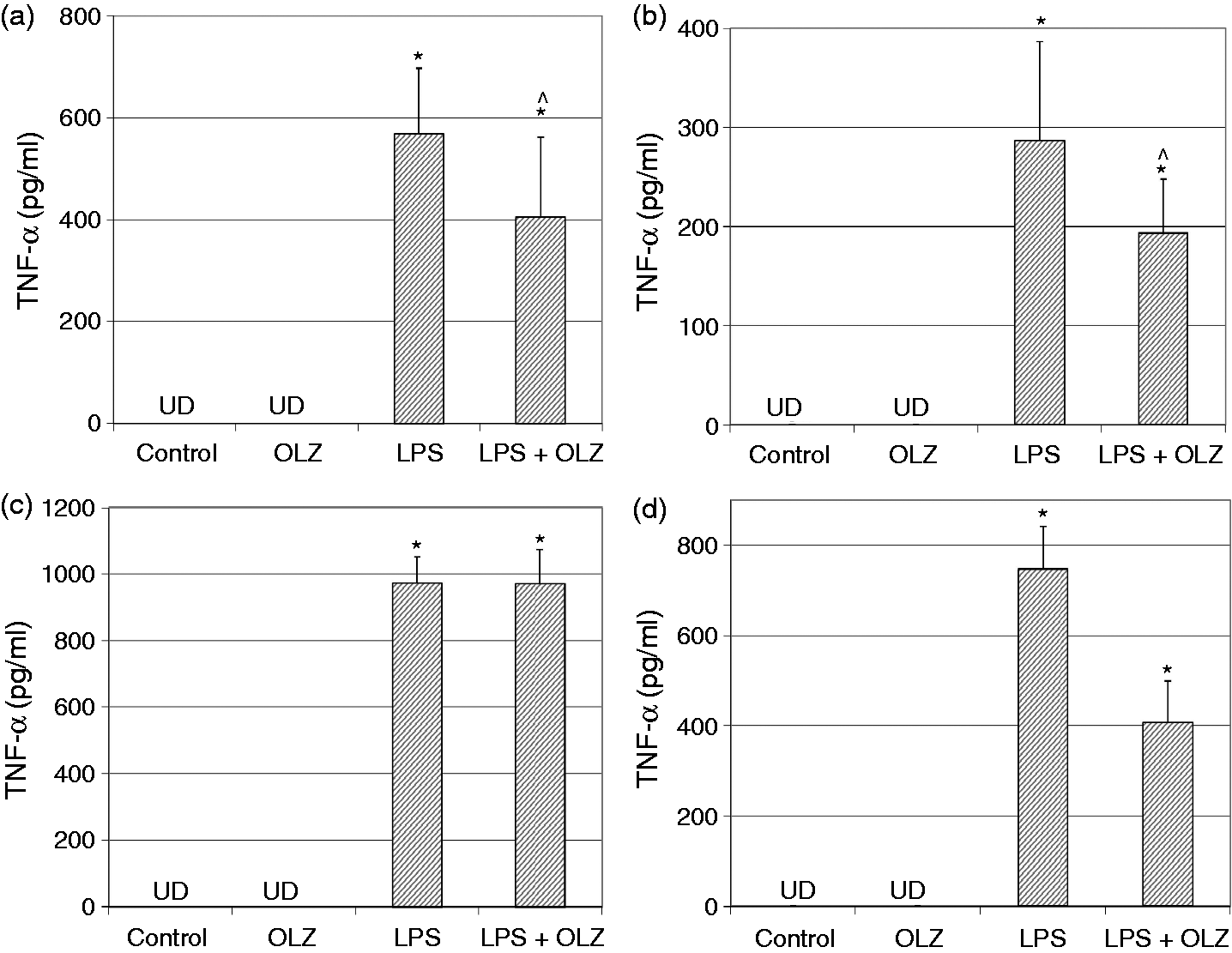
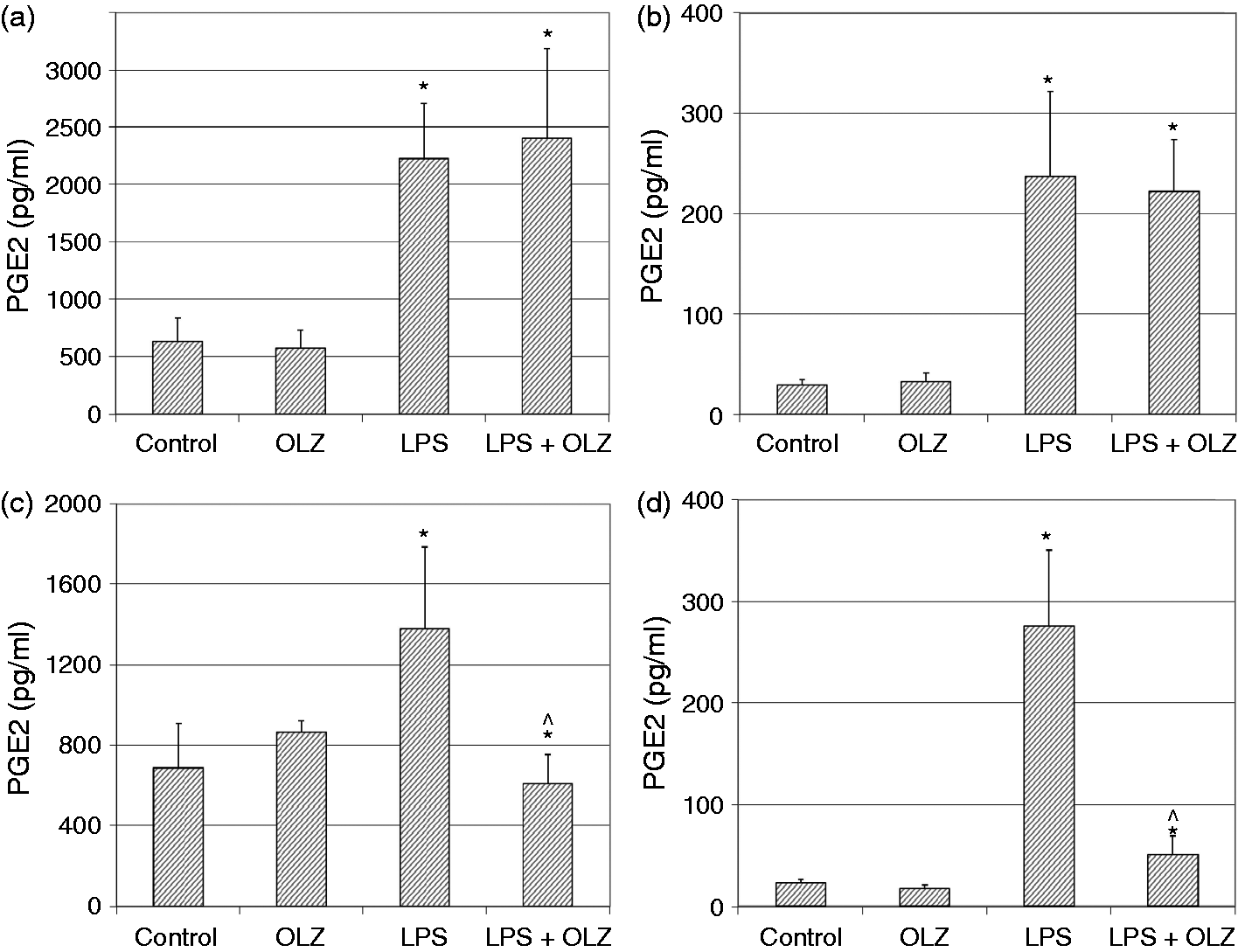
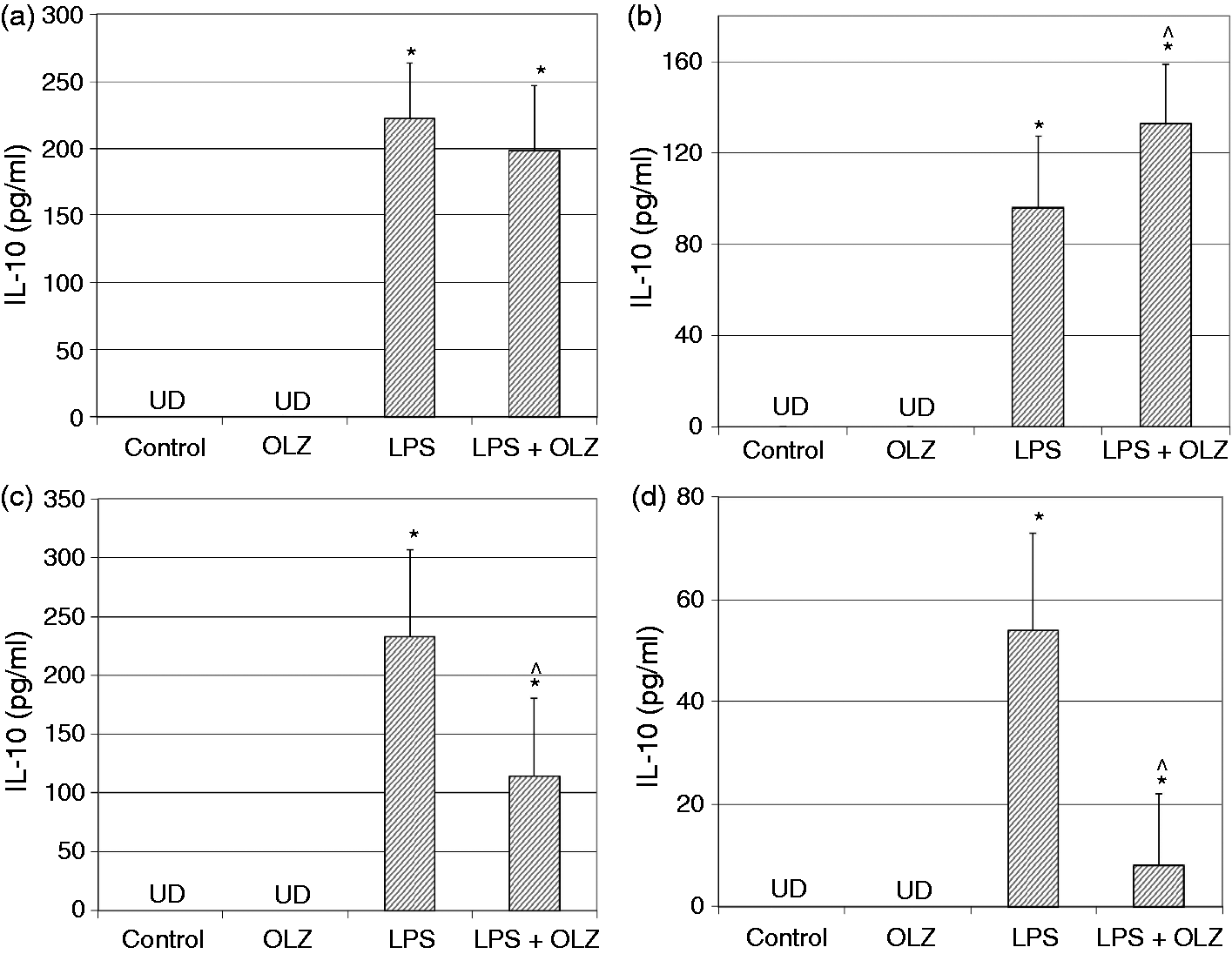
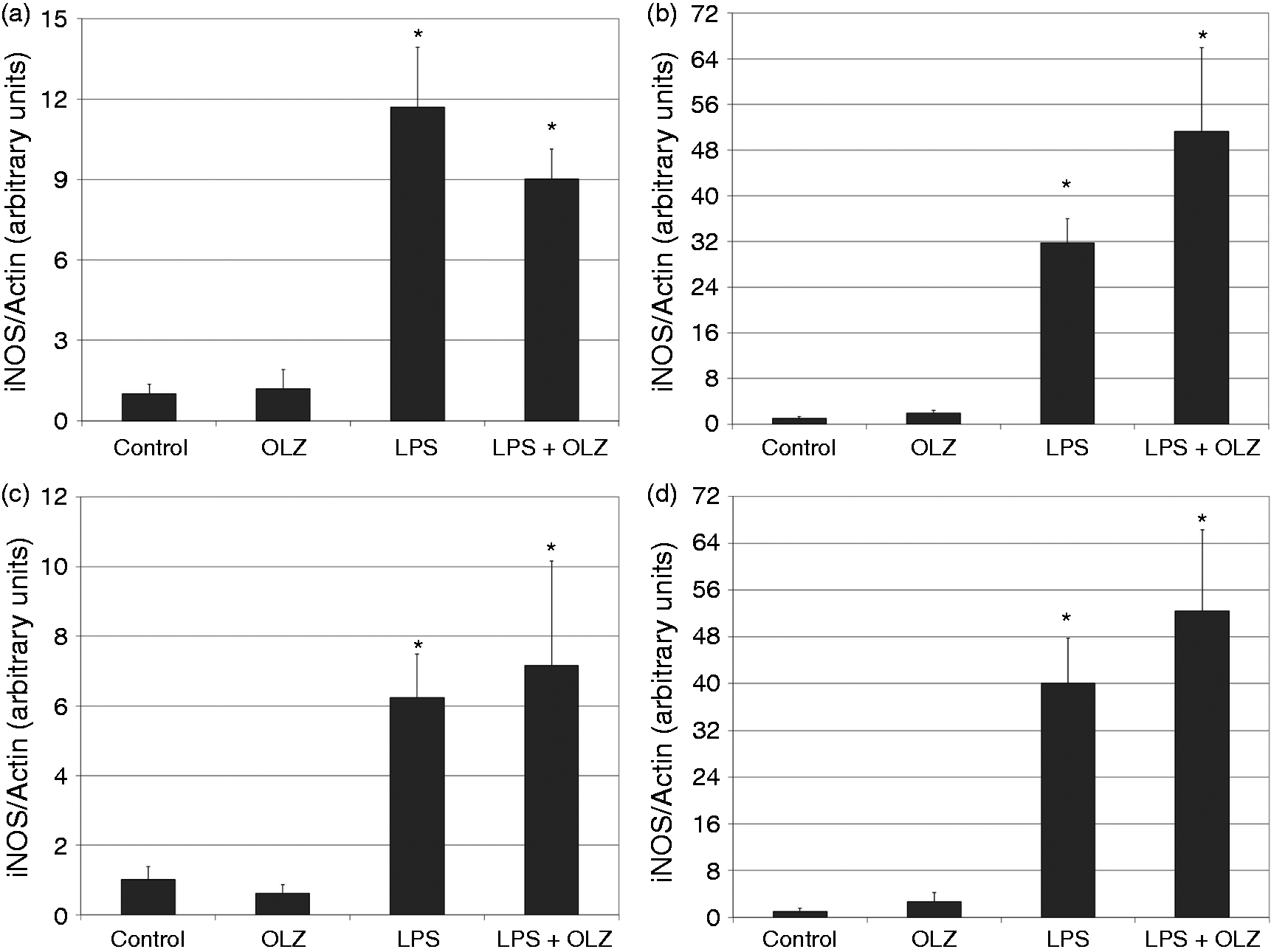
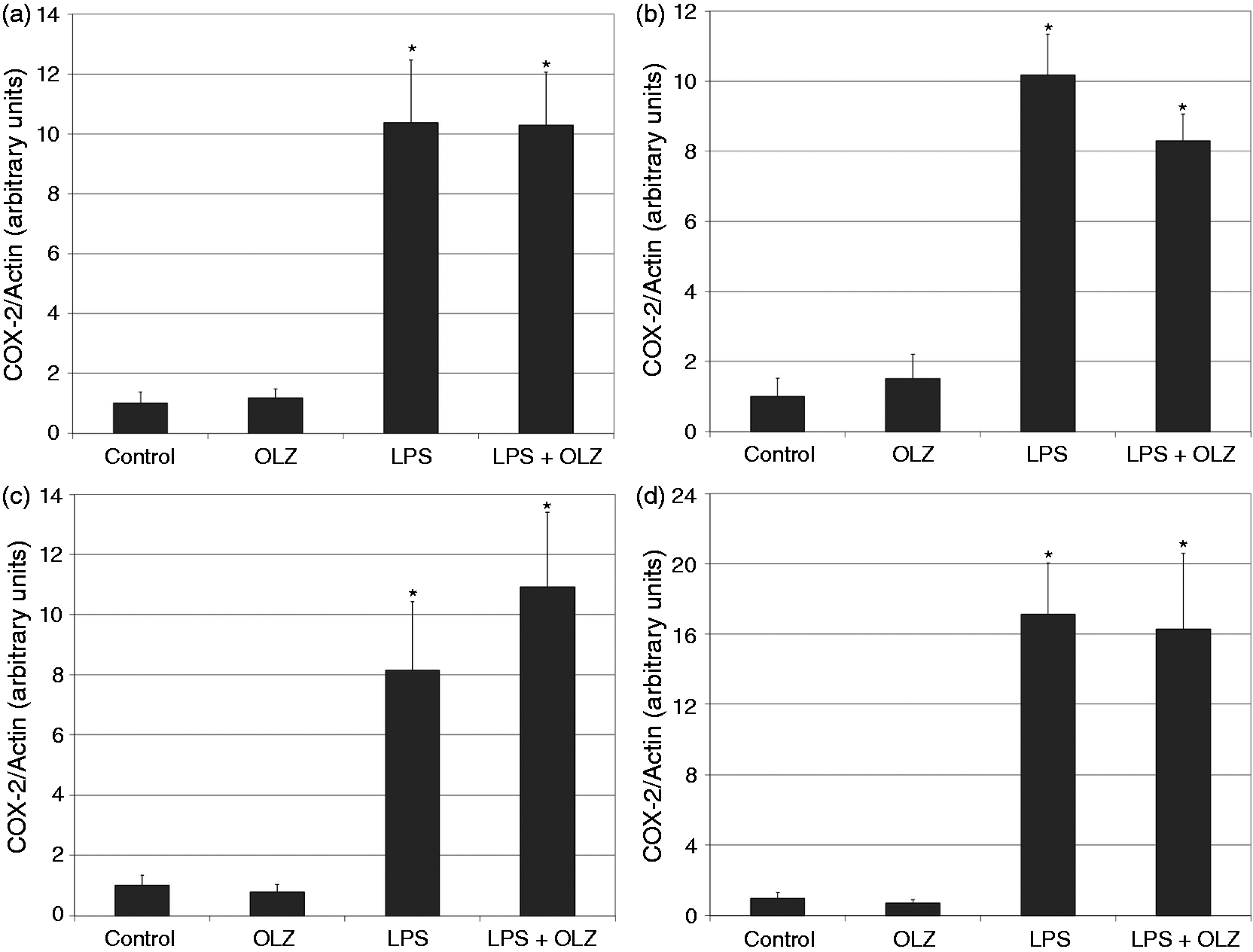
Effects of OLZ on LPS-induced secretion of NO. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (100 ng/ml) was added to the culture medium for another 18 h. NO levels in the culture medium were determined by the Griess reaction assay. Values are presented as mean ± SD of 6–18 samples in each group. *P < 0.05 vs. control; ^P < 0.05 vs. LPS. Effects of OLZ on LPS-induced secretion of TNF-α. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (100 ng/ml) was added to the culture medium for another 18 h. TNF-α levels in the culture medium were determined by ELISA. Values are presented as mean ± SD of 6–18 samples in each group. UD: undetectable. *P < 0.05 vs. control; ^P < 0.05 vs. LPS. Effects of OLZ on LPS-induced secretion of PGE2. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (5 µg/ml) was added to the culture medium for another 18 h. PGE2 levels in the culture medium were determined by ELISA. Values are presented as mean ± SD of 6–18 samples in each group. *P < 0.05 vs. control; ^P < 0.05 vs. LPS. Effects of OLZ on LPS-induced secretion of IL-10. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (100 ng/ml) was added to the culture medium for another 18 h. IL-10 levels in the culture medium were determined by ELISA. Values are presented as mean ± SD of 6–18 samples in each group. UD: undetectable. *P < 0.05 vs. control; ^P < 0.05 vs. LPS. Effects of OLZ on LPS-induced expression of iNOS. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (5 µg/ml) was added to the culture medium for another 18 h. Thereafter, cells were lysed as described in the ‘Materials and methods’ and aliquots of total cell protein (20 µg per lane) were separated on 10% SDS/polyacrylamide gel. iNOS protein was identified with a specific anti-iNOS Ab. Actin was used as an internal control to normalize for loading variations. Illustrations represent quantitative analyses of three immunoblots. (A–D) In each illustration bars represent means ± SEM of relative iNOS expression quantified densitometrically from three independent experiments in which each treatment group included two samples (flasks). Owing to variations in LPS induction of iNOS expression in different experiments, expression is presented relative to its level in the control group, which is arbitrarily expressed as 1. *P < 0.05 vs. control. Effects of OLZ on LPS-induced expression of COX-2. Primary rat glia cells were pretreated with OLZ (A, B) 1 µM or (C, D) 50 µM for (A, C) 6 h or (B, D) 72 h. Then, LPS (5 µg/ml) was added to the culture medium for another 18 h. Thereafter, cells were lysed as described in the ‘Materials and methods’ and aliquots of total cell protein (20 µg per lane) were separated on 10% SDS/polyacrylamide gel. COX-2 protein was identified with a specific anti-COX-2 Ab. Actin was used as an internal control to normalize for loading variations. Illustrations represent quantitative analyses of three immunoblots. (A–D) In each illustration bars represent means ± SEM of relative COX-2 expression quantified densitometrically from three independent experiments in which each treatment group included two samples (flasks). Owing to variations in LPS induction of COX-2 expression in different experiments, expression is presented relative to its level in the control group, which is arbitrarily expressed as 1. *P < 0.05 vs. control.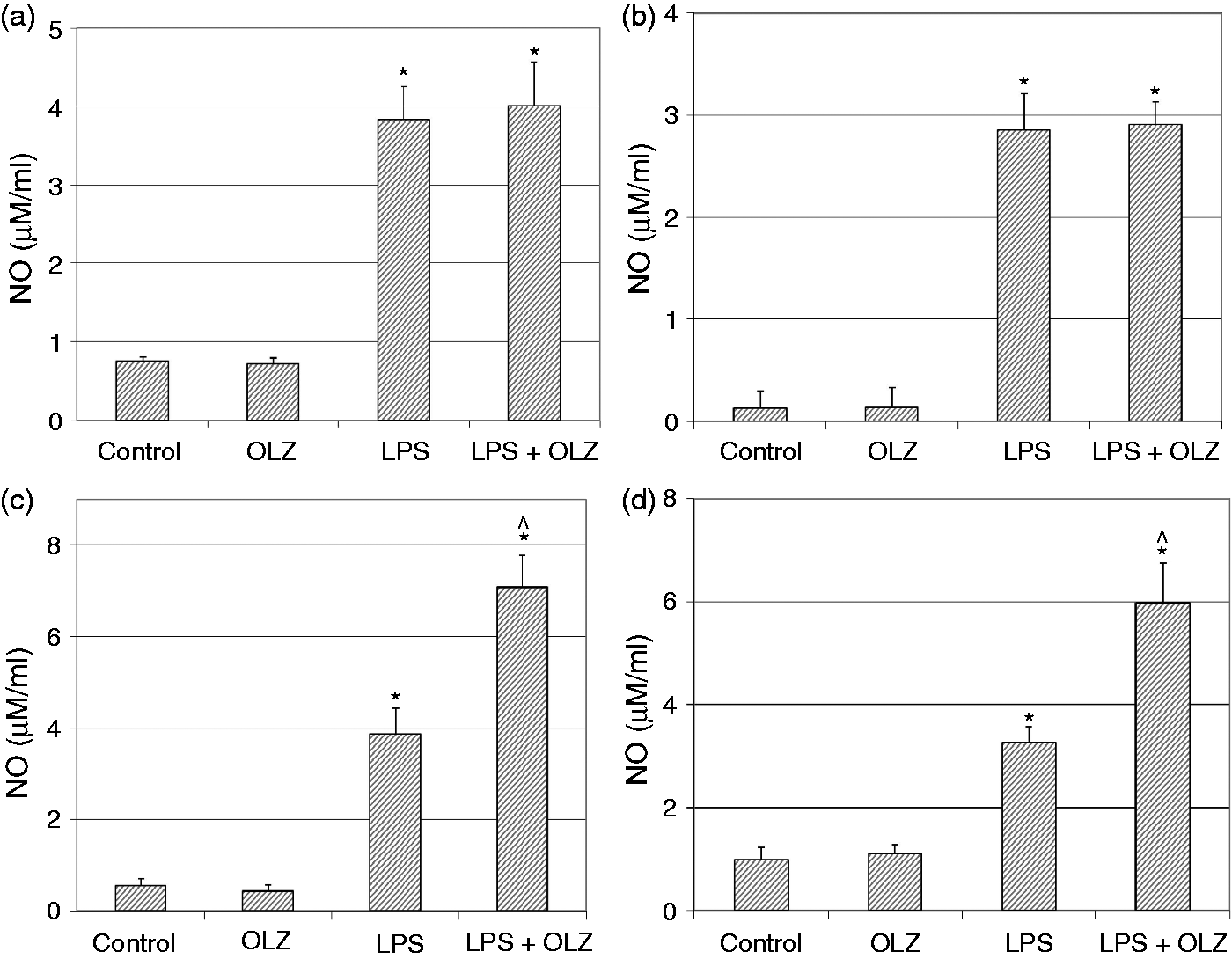
Results
Effects of OLZ on LPS-induced secretion of NO, TNF-α, PGE2 and IL-10
Treatment with LPS (100 ng/ml) significantly increased NO secretion in all the experiments performed (Figure 1). Pretreatment with OLZ 1 µM for 6 and 72 h did not alter LPS-induced secretion of NO (Figure 1A and B, respectively). However, pretreatment with OLZ 50 µM for 6 and 72 h significantly increased LPS-induced secretion of NO (Figure 1C and D, respectively). These results indicate that pretreatment with the higher concentration of OLZ enhanced NO production irrespective of the duration of exposure to the drug.
Treatment with LPS (100 ng/ml) significantly increased TNF-α secretion in all the experiments performed (Figure 2). Pretreatment with OLZ 1 µM for 6 and 72 h significantly decreased LPS-induced secretion of TNF-α (Figure 2A and B, respectively). Similarly, pretreatment with OLZ 50 µM for 72 h (but not for 6 h) significantly reduced LPS-induced secretion of TNF-α (Figure 2D). These results generally suggest that pretreatment with OLZ decreases TNF-α production.
Treatment with LPS (5 µg/ml) significantly increased PGE2 secretion in all the experiments performed (Figure 3). It is important to emphasize that basal and induced levels of PGE2 were markedly higher in the short-term (6 h) treatment protocol (Figure 3A, C). Pretreatment with OLZ 1 µM for 6 and 72 h did not alter LPS-induced secretion of PGE2 (Figure 3A and B, respectively). In contrast, pretreatment with OLZ 50 µM for 6 and 72 h significantly decreased LPS-induced secretion of PGE2 (Figure 3C and D, respectively). These results indicate that only the higher concentration of OLZ reduced PGE2 production.
Treatment with LPS (100 ng/ml) significantly increased IL-10 secretion in all the experiments performed (Figure 4). Pretreatment with OLZ 1 µM for 6 h did not alter IL-10 levels, while pretreatment for 72 h significantly increased LPS-induced secretion of IL-10 (Figure 4A and B, respectively). However, pretreatment with OLZ 50 µM for 6 and 72 h significantly decreased LPS-induced secretion of IL-10 (Figure 4C, D). These results indicate that pretreatment with the higher concentration of OLZ reduced IL-10 production.
Effects of OLZ on LPS-induced expression of iNOS and COX-2
As seen in Figure 5, treatment with LPS (5 µg/ml) significantly enhanced the expression level of iNOS. Pretreatment with OLZ 1 and 50 µM did not significantly alter LPS-induced expression of iNOS irrespective of the duration of exposure to the drug (Figure 5). However, it is worth noting that OLZ, particularly at 50 µM, led to a trend of enhancing iNOS expression (Figure 5B–D). This may partially explain the significant increase in NO secretion under treatment with OLZ 50 µM (Figure 1).
As seen in Figure 6, treatment with LPS (5 µg/ml) significantly increased the expression level of COX-2. Pretreatment with OLZ 1 and 50 µM did not significantly alter LPS-induced expression of COX-2 irrespective of the duration of exposure to the drug (Figure 6).
Discussion
The present study found that OLZ altered LPS-induced inflammation in rat primary glia cells and differently affected the secretion of various inflammatory mediators. Under certain conditions OLZ exhibited some anti-inflammatory effects including reduction of TNF-α and PGE2 secretion. However, it also exerted pro-inflammatory effects such as enhancement of NO production and reduction of IL-10 secretion. These findings are in line with previous reports in which OLZ was found to exert both anti- and pro-inflammatory effects.10,26,32,38–40,73
In this study we used two concentrations of OLZ: 1 and 50 µM. Unlike most of the drugs used for prophylactic treatment in schizophrenia and bipolar disorder, OLZ is a drug whose plasma concentration is not routinely monitored in treated patients. Clinical data indicate that plasma concentrations of 20 ng/ml (equivalent to 65 nM) and higher are associated with a positive clinical response.77–79 Such plasma concentrations are usually obtained when adult patients are treated with a daily dose of 10–20 mg/d.77–79 It is worth noting that most of the studies that examined the pharmacological effects of OLZ in vitro used much higher concentrations of the drug. For example, Kim et al. tested the anti-apoptotic effects of OLZ in human neuroblastoma SH-SY5Y cells using OLZ concentrations between 10 and 100 µM. 80 Similarly, Lu and Dwyer examined the effects of OLZ on neurite growth in PC12 pheochromocytoma cells using 1, 10 and 40 µM of the drug. 81 Nevertheless, those studies do not disprove the fact that 50 µM OLZ is a high and probably therapeutically irrelevant concentration of the drug. In our preliminary experiments we examined two other concentrations of OLZ: 100 nM, a therapeutically relevant concentration; and, 10 µM, a relatively intermediate concentration between 1 and 50 µM. We found that 100 nM OLZ did not alter the inflammatory mediators examined in this study, whereas 10 µM OLZ led to similar results as obtained with 1 µM OLZ (data not shown).
Activation of glia cells is the hallmark of CNS inflammation. Activated glia cells initiate and facilitate the secretion of a diverse array of pro-inflammatory mediators including cytokines, PGs and free radicals such as NO.28,44–46,50,51,82 TNF-α is a prominent pro-inflammatory cytokine, the secretion of which is associated with activation of other inflammatory mediators.45,83,84 TNF-α has both deleterious and protective effects on neurons and it activates the expression of many NF-κB-regulated genes that are important for cell survival. 85 Few studies examined the effects of OLZ on TNF-α secretion, most of which used peripheral tissues rather than brain cells as a model system. For example, Victoriano et al. found that chronic OLZ treatment (46 d) in rats significantly increased white adipose tissue TNF-α levels. 38 In the present study we found that, overall, OLZ significantly reduced LPS-induced secretion of TNF-α (Figure 3). The reduction in TNF-α was more profound when OLZ was added to the culture medium at 72 h (as compared with 6 h) before LPS induction. Thus, a longer pretreatment duration enhanced the TNF-α inhibitory effect of OLZ. The therapeutic effects of OLZ and other psychotropic drugs take 2–6 wk to emerge. Therefore, a chronic treatment protocol is more relevant to the pharmacology of mental illnesses. However, owing to a technical limitation, we could not conduct a typical chronic treatment protocol: primary glia cells become ready for use 2–3 wk after extraction, and they remain viable for another 1–2 wk. This limitation does not allow the conduction of a long-term (4–6 wk) treatment protocol. Nonetheless, we aimed to prolong as much as possible the exposure time to OLZ in order to examine whether a time-dependent effect exists. Indeed, our results showed that the duration of pretreatment with OLZ influenced the effects of the drug on secretion of inflammatory mediators such as IL-10 and TNF-α.
In humans, NO is recognized as a relaxant of smooth muscles in blood vessels and airways but is also linked to inflammation. NO is produced in the brain by three enzymes: endothelial NOS, neuronal NOS and iNOS. 82 Glia cells secrete NO mainly via iNOS, which is expressed after exposure to inflammatory stimuli such as TNF-α, IL-1β, IFN-γ and bacterial endotoxins. 85 NO participates in neuronal transduction, apoptosis, blood vessels relaxation, immune system activity and pain, among other functions. In the present study we found that pretreatment with OLZ 1 µM did not alter LPS-induced secretion of NO, whereas OLZ 50 µM led to a significant increase in NO levels (Figure 1). Our Western blot analysis revealed that pretreatment with OLZ did not significantly alter iNOS expression (Figure 5). However, pretreatment with OLZ 1 µM for 72 h and with 50 µM for 6 and 72 h was associated with a clear trend for increase in iNOS expression (Figure 5).
PGE2 is another mediator that was tested. PGE2 is involved in many physiological, as well as pathophysiological conditions such as vascular vasodilatation, female reproduction, gastric mucosal protection, normal kidney function, fever, induction of pain, activation of inflammation and tumorigenesis.60–63 We found that pretreatment with OLZ 1 µM did not affect LPS-induced secretion of PGE2, whereas OLZ 50 µM significantly decreased PGE2 levels (Figure 3). These findings are in line with the results of a previous study in which chronic treatment with OLZ led to a significant reduction in PGE2 levels in rat brain. 73 Interestingly, pretreatment with OLZ 50 µM did not significantly alter the expression of COX-2 in LPS-treated cells (Figure 6). It is possible that the reduction in PGE2 levels under treatment with OLZ 50 µM derive from a post-translational modification of COX-2 activity rather than a decrease in COX-2 expression.
The next cytokine that we examined was IL-10. IL-10 is a recognized anti-inflammatory cytokine that has been shown to suppress a broad range of inflammatory responses.86,87 Pretreatment with OLZ 1 µM for 72 h significantly increased LPS-induced secretion of IL-10 (Figure 4), attesting to a possible anti-inflammatory effect of the drug under these conditions. On the other hand, pretreatment with OLZ 50 µM for 6 and 72 h significantly decreased IL-10 levels in LPS-treated cells (Figure 4), suggesting that an increased concentration of the drug accentuates its pro-inflammatory effects. However, as mentioned above, 50 µM OLZ is a high concentration of the drug. Thus, an enhancing (anti-inflammatory) effect of OLZ on IL-10 is a more reasonable occurrence when the drug is given at a therapeutic concentration. In line with this assumption, in preliminary experiments we observed that 10 µM OLZ significantly increased IL-10 levels in LPS-treated glia cells (data not shown).
Inflammatory mediators are known to influence neurotransmission, cognition, behavior and other crucial functions in the brain.15,88–92 Therefore, the impacts of different psychotropic drugs on levels of inflammatory mediators in the brain most likely affect their therapeutic as well as toxic, effects. In the present study we found that OLZ exhibited both anti- and pro-inflammatory effects. However, under the experimental conditions used, OLZ lacked a prominent effect on LPS-induced inflammation. In this regard, our study has two limitations: (i) we used an extra-therapeutic concentration of OLZ (50 µM), and (ii) we examined the effects of OLZ on LPS-induced inflammation only at two time points; perhaps, determining the effects of the drug on more time points would have provided a more scientifically sound insight into the actual effects of OLZ on LPS-activated glia cells. Therefore, it is difficult to speculate how OLZ will affect brain inflammation when it is given chronically at therapeutic concentrations to psychiatric patients. More research is necessary to understand the kinetic and mechanism underlying the effects of OLZ on brain inflammation.
Footnotes
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.