
Abstract
Dendritic cell (DC) subsets form a remarkable cellular network that regulate innate and adaptive immune responses. Although pigs are the most approximate model to humans, little is known about the regulation of monocyte-derived DCs (moDCs) and splenic DCs (SDCs) in the initiation of immune responses under inflammatory conditions. We investigated the activation and maturation of porcine moDC and SDC subpopulations following LPS stimulation. Porcine monocytes that would differentiate into moDCs were isolated. SDCs were isolated directly from the porcine spleen. Following LPS stimulation, phagocytosis activity, TLR4/MyD88-dependent gene expression, co-stimulatory molecule, and pro-inflammatory cytokine (TNF-α, IL-1β) and chemokine (IL-8) expressions were increased in both cell subsets. Furthermore, moDCs showed higher levels of gene and protein expression compared with SDCs. Interestingly, moDCs were found to be more responsive via the TLR4/TRAF-dependent signalling pathway of activation. Only SDCs expressed higher level of IL-12p40 gene and protein, whereas, IFN-γ gene and protein expression were likely to be unchanged after LPS stimulation in both cell subtypes. These data demonstrate that porcine moDCs display a greater ability to initiate innate immune responses, and could be used as a model to investigate immune responses against Ags.
Introduction
Dendritic cells (DCs) are highly specialised APCs, and play a critical role in bridging the innate and adaptive immune systems by recognising Ags via complex mechanisms that involve the decoding and integration of various signals received in a receptor-dependent manner.1,2 According their stage of functional development, DCs can be divided in two major categories: immature and mature. Immature DCs are characterised by active up-take and processing of Ags through high expression of cell surface pattern recognition receptors (PRRs) and low levels of co-stimulatory molecules (CD80 and CD86), 3 which lead to phagocytosis of the latter. Upon capturing foreign Ags, immature DCs become mature and migrate to secondary lymphoid organs, where they activate T-cell and initiate adaptive immunity. 4 Spleen-resident DCs have been categorised into two major subsets on the basis of their phenotype: conventional/myeloid DCs are characterised by CD11c+HLA-DR+CD123−, and express TLRs 2 and 4; plasmacytoid DCs exhibit CD11c−HLA-DR+CD123+ in human and mice, and express TLRs 7 and 9. 5 An appropriate porcine DCs model is important to investigate their immune responses in vitro, ignoring the involvement of microenvironments in secondary lymphoid tissues that might be important in determining their functions. Notably, the culture of precursor monocytes with granulocyte-macrophage colony-stimulation factor (GM-CSF) and IL-4 leads to the development of immature monocyte-derived DCs (moDCs) models in vitro.6,7 When mouse and human splenic CD11c+ immature DCs (SDCs) capture LPS or other pathogens, they can further differentiate into mature DCs, and up-regulate MHC-II, CD40, CD80, CD83 and CD86 on their surface. 8 Presentation of a pathogen by different DC subtypes is assumed to have an impact on the innate immune response. However, the relative efficacy of the different CD11c+ SDCs and moDCs has not been compared in pigs. Additionally, there is a dearth of knowledge about the generation, activation and the pattern of immune response of moDCs in pigs.
Notably, the unique immune function of DCs is that immature DCs become mature DCs by recognising invasive pathogens through their ability to detect PAMPs using PRRs. 9 TLRs are the best-characterised PRRs, and are linked to the control of bacterial and viral infection. 10 TLR4 is stimulated by the PAMP LPS or endotoxin from Gram-negative bacteria. LPS is one of the best studied immunostimulatory bacterial components that can induce systemic inflammation. LPS interacts with DCs via the TLR4/MD2/CD14 receptor complex, and induces the release of the essential pro-inflammatory cytokines and chemokines to activate a potent immune response. 11 Two signalling pathways—MyD88-dependent and MyD88-independent (TRIF-dependent) associated with TRAF6—have been described for TLR4 activation following LPS stimulation. Activation of the MyD88-dependent pathway drives a rapid activation of NF-κB and produces pro-inflammatory cytokines, including TNF-α, IL-1β, IL-12p40 and IL-6, and chemokines, such as monocyte chemoattractant protein 1, macrophage inflammatory protein 3α and IL-8.12,13 However, LPS activation of the MyD88-independent signalling pathway results in rapid activation and nuclear translocation of IFN regulatory factor 3 (IRF3), which leads to activation of the type 1 IFN signalling pathway. 14 It has been reported that IFN-β—a type 1 IFN—plays an important role in cell survival, protein synthesis and control of inflammatory responses against Ags. 15 However, there has been no study to distinguish the relevance of the immune response of moDCs and SDCs in pigs with regard to the TLR signalling pathway. Therefore, it is important to compare the MyD88-dependent and MyD88-independent signalling pathways of immune responses between moDCs and SDCs.
We therefore evaluated the immune responses of LPS-induced SDCs and moDCs in vitro for use as a model. We analysed the impact of various concentrations of LPS on phagocytosis and cell viability response, and investigated MyD88-dependent and MyD88-independent related gene expression in LPS-stimulated porcine moDCs and SDCs in a time-dependent manner. Furthermore, the production of co-stimulatory molecules, pro-inflammatory cytokines and chemokines were measured.
Materials and methods
Animals
Three 35-d-old Pietrain female piglets were used for this study. The piglets were clinical healthy and no respiratory disease was found in the clinical history or upon physical examination. Animals were housed in isolation rooms at the teaching and research station of Frankenforst, University of Bonn, Germany. The feeding, housing and husbandry practices of the animals followed the standard guidelines. 16 The experiment was approved by the Veterinary and Food Inspection Office, Siegburg, Germany. All the piglets were euthanased by i.v. injection of T61 (Intervet) by a registered veterinarian after sedation using ketamine (Intervet, Unterschleißheim, Germany), according to German animal protection law. 16
Generation of moDC from porcine PBMC
PBMCs were isolated from peripheral blood. Blood samples were collected from the vena cava cranialis in sterilised tubes with EDTA. The PBMCs were isolated by Ficoll-Paque density gradient (cat. #10771; Sigma; Seelze, Germany) centrifugation, as described previously.17,18 PBMCs were washed twice in Dulbecco's phosphate-buffered saline (DPBS) (cat. #1419-169; Life Technologies; Darmstadt, Germany) and re-suspended in DMEM (cat. #41966-029; Invitrogen; Darmstadt, Germany) supplemented with 2% FBS (cat. #10270; Invitrogen; Darmstadt, Germany), 500 IU/ml penicillin-streptomycin (cat. #15140; Invitrogen; Darmstadt, Germany) and 0.5% fungizone (cat. #15290-026; Invitrogen; Darmstadt, Germany). These PBMCs (5 × 106 cell/ml) were cultured in six-well plates (2 ml/well) for 4 h. The moDCs were generated following the procedure described previously.19–21 Briefly, after 4 h of incubation, non-adherent cells were removed and the adherent monocytes were washed twice with pre-warmed (37℃) DPBS in order to purify the cells. The purified monocytes were cultured in RPMI-1640 medium (cat. #21875; Invitrogen; Darmstadt, Germany) supplemented with 10% FBS, 1000 UI/ml penicillin-streptomycin, 1% fungizone, 20 ng/ml recombinant porcine (rp) GM-CSF (cat. #711-PG-010; R&D System; Wiesbaden, Germany) and 20 ng/ml rp IL-4 (cat. 654-P4-025; R&D Systems; Wiesbaden, Germany) for 7 d at 37℃ with 5% CO2. Half of the medium was replaced every third day with fresh medium. After 7 d of incubation, the adherent moDCs were counted and re-cultured in a new plate for subsequent assays.
SDC isolation
The spleen was collected under sterile conditions within 30 min of euthanasia from the same pigs used for blood sampling. Splenic tissues were cut into 5–7-cm-long sections and flushed thoroughly with 0.9% NaCl. The tissues were transported in Hanks-EDTA (cat. H4641; Sigma; Seelze, Germany) solution, on ice, to the laboratory for DC isolation. 22 Briefly, the tissue pieces were transferred into collagenase solution (collagenase 1 mg/ml (cat. #C2139-100 MG; Seelze; Germany), BSA 1 mg/ml (bovine serum albumin: cat. #CA2153; Sigma; Seelze, Germany); DNase 10 UI/ml (cat. #C 79254; Qiagen; Dusseldorf, Germany), penicillin-streptomycin 500 UI/ml and fungizone 0.25%) and incubated in a shaking (120 rpm) incubator at 37℃. After 2–4 h of incubation, the collagenase solution containing released cells was filtered through a 70-µm cell strainer (cat. #352350; BD-Falcon; Heidelberg, Germany) followed by filtration through a 40-µm cell strainer (cat. #352340; BD-Falcon; Heidelberg, Germany). Cells were washed twice in DPBS and re-suspended in RPMI-1640 medium supplemented with 10% FBS, 1000 UI /ml penicillin-streptomycin and 1% fungizone. In order to purify the SDCs, cells were seeded (1 × 107 cells/ml) into a six-well plate (2 ml per well) and incubated for 12–16 h. Finally, the non-adherent cells were removed by gently washing twice in warm RPMI-1640 medium; then, the SDCs were cultured for the further study.
Stimulation of moDCs and SDCs
moDCs and SDCs were seeded separately at 2 × 106 cells/well in six-well tissue culture plates and incubated overnight at 37℃ in a CO2 incubator. Afterwards, both cell types were treated with LPS (cat. #tlrl-3pelps; InvivoGen; Toulouse, France) at concentrations of 0.5, 1.0 and 2.0 µg/ml for 24 h for a cell viability assay. For the gene expression study and cytokine measurement, cells were stimulated with 1 µg/ml LPS and harvested at 0, 3, 6, 12 and 24 h after stimulation. Total RNA was isolated from these cells for gene expression analysis, and the culture supernatant was used to measure the cytokine levels. SDCs and moDCs stimulated or non-stimulated for 24 h with 1 µg/ml LPS were used for the expression of co-stimulatory molecules (CD40 and CD86).
Cell viability assay
Cell viability was investigated using the WST-1 cell proliferation kit (cat. #10008883; Cayman Chemical; Hamburg, Germany), following the manufacturer’s instructions. Briefly, SDCs and moDCs were cultured in 96-well plates at a density of 104–105 cells/well in 100 µl of RPMI-1640 medium for 24–48 h in a CO2 incubator at 37℃ before LPS stimulation. This allowed the cells to adhere to the surface of the microplate. Cells were cultured, untreated or treated with LPS, at concentrations of 0.5, 1.0 and 2.0 µg/ml for 24 h. Ten microlitres of reconstituted WST-1 mixture was added to each well. After 2 h of incubation in a CO2 incubator at 37℃, the absorbance of the samples was measured using a microplate reader (Thermo Max, Wesel, Germany) at a wavelength of 450 nm. Cell viability was calculated (%) following the manufacturer’s formula.
DC maturation and cell type measurement using flow cytometry
For FACS analysis, cells were harvested, washed twice with DPBS and cell number counted. The cells were incubated for 30 min with FACS staining buffer (DPBS supplement with 2% FBS, 10 mM NaN3 and 10 mM HEPES) and then washed with the same staining buffer. Cells were stained either with mouse anti-human CD11c phycoerythrin-cyanine 7 Ab (clone N418, 25-0114; BD eBioscience; Heidelberg, Germany), a mouse anti-human CD40 FITC Ab (clone G28.5, NB100-77786; Novus Biologicals; Cambridge, UK) or a mouse anti-human CD86 APC Ab (clone hB7-2, FAB141A; R&D Systems; Wiesbaden, Germany) Ab. All these Abs are of the IgG1 isotype. Cells were stained for 30 min on ice in 100 µl of staining buffer solution under light-protected conditions. The cells were washed twice using staining buffer and re-suspended in 400–500 μl staining buffer. The events were acquired on 10,000 cells using a FACSCalibur Dual Laser Flow Cytometer (BD Biosciences, Sparks, MD, USA) and analysed by FlowLogic software (BD Biosciences, Heidelberg, Germany).
Gene expression study using quantitative real-time PCR
List of primer sequences used in this study.
Phagocytosis assay
Phagocytosis activity of the cells was investigated using a Vybrant Phagocytosis Assay kit (cat. #V-6694; Molecular Probes; Goettingen, Germany). The principle of the test is based on the intracellular florescence emitted by the engulfed particles, as well as the effective fluorescence quenching of the extracellular probe by Trypan Blue. 24 The phagocytosis assay protocol uses five negative controls, five positive controls and four experimental samples. Briefly, cells were cultured in a six-well plate for 48 h with RPMI-1640. Cells were scraped from the six-well plate, washed twice with DPBS, and the cell density adjusted to 106 cells/ml. For the phagocytosis assay, 50 µl of the different concentration of LPS (0.5, 1.0 and 2 µg/ml) in RPMI-1640 were added to the experimental wells containing cells. The cells were incubated for 4 h at 37℃ with 5% CO2 to allow the cells to adhere to the microplate surface. Afterwards, the RPMI-1640 solution was removed from the microplate wells. Then, 100 µl of fluorescent BioParticle (Vybrant Phagocytosis Assay kit) suspension was added to all the negative control, positive control and experimental wells. Two h after incubation at 37℃ in a CO2 incubator, the BioParticle suspension was aspirated from all of the wells. Finally, 100 µl Trypan Blue was added to the wells and incubated for 1 min at room temperature and then the Trypan Blue was aspirated immediately. The fluorescence emission was measured in a fluorescence microplate reader (Thermo Electron, Waltham, MA, USA) using 480 nm excitation and 520 nm emission. The net phagocytosis of the cells was calculated according to the response of the phagocytosis effector agent following the manufacturer’s instructions.
Cytokine measurements by ELISA
ELISA was used to investigate the differences in cytokine secretion of moDC and SDCs at different time points. moDC and SDC (1 × 106 cells/ml) were stimulated with 1 µg/ml LPS for 0, 3, 6, 12 and 24 h, and supernatants were collected for TNF-α (cat. #PTA00; R&D Systems), IL-8 (cat. #P8000; R&D Systems), IL-1β (cat. #PLB00B; R&D Systems), IL-12p40 (cat. #P1240; R&D Systems) and IFN-γ (cat. #PIF00; R&D Systems) measurement using ELISA according to manufacturer’s instructions. The results were determined using a microplate reader (ThermoMax) set at 450 nm.
Statistical analysis
The data were analysed by SAS version 9.2 (SAS Institute, Cary, NC, USA). Pairwise comparisons were made between the treatment groups and control using the Student’s t-test and Tukey’s multiple comparison test. The data were expressed as means ± SD and P < 0.05 was determined to be statistically significant.
Results
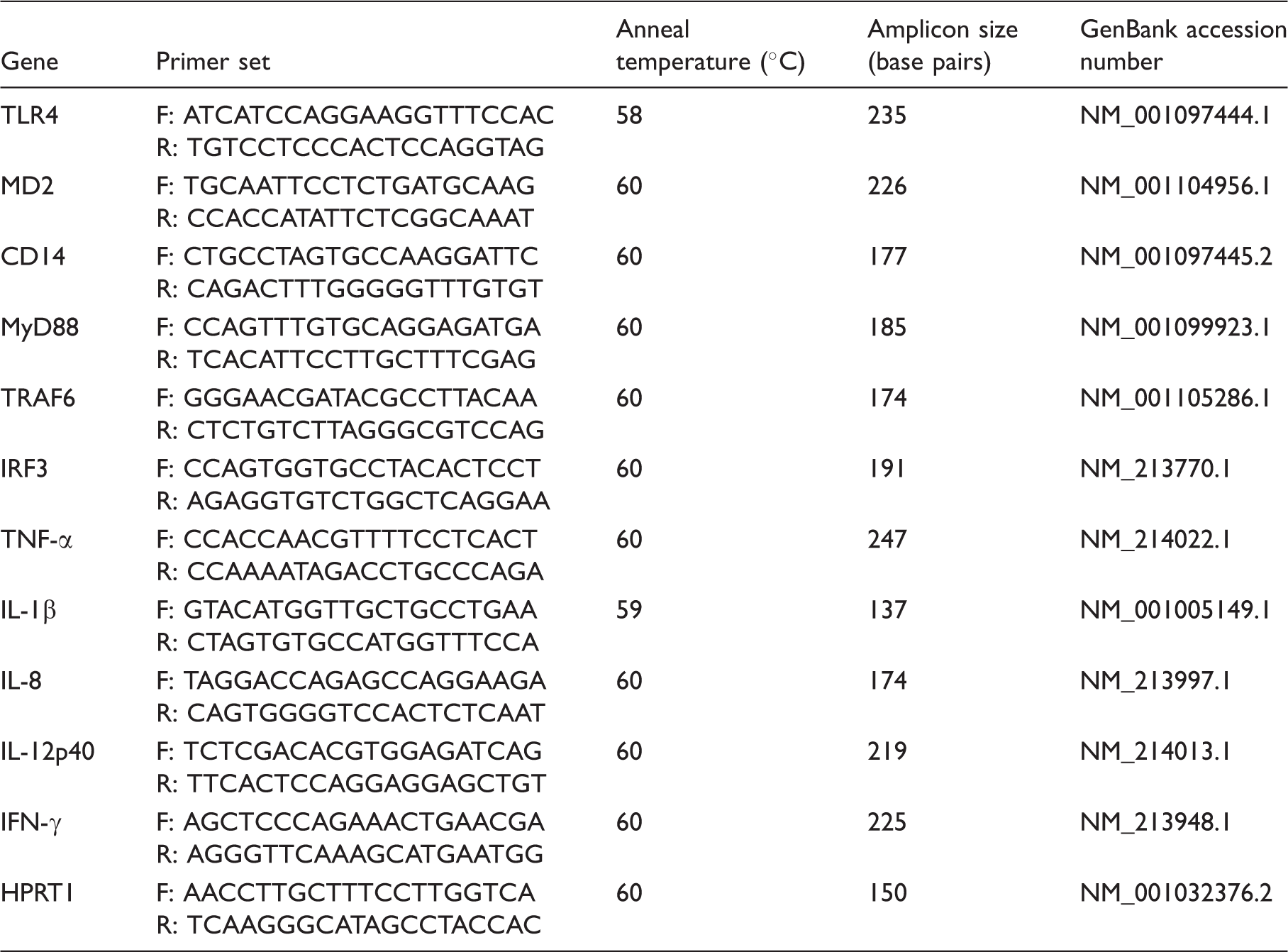
Different LPS concentrations affect moDCs and SDCs viability
To determine the oxidative stress and cytotoxicity effects of LPS on porcine DCs, we detected cell viability with different concentrations of LPS.
25
The viability of both cell types was more affected by a higher LPS concentration than a lower LPS concentration (Figure 1A). Viability was significantly decreased in moDCs treated with 1.0 and 2.0 µg/ml LPS. It was found that only in the case of treatment with 2.0 µg/ml LPS was moDC viability decreased compared with SDCs (Figure 1B). These results indicate that moDCs are more sensitive to LPS exposure; on the basis of these data, 1.0 µg/ml LPS was considered for further studies.
Effects of LPS stimulation on cell viability of moDCs and SDCs. moDCs at d 7 and SDC at d 3 were treated or not with LPS (control, 0.5, 1.0 and 2.0 µg/ml). Comparison of moDC or SDC cell viability after challenge with different concentrations of LPS (A), and cell viability of moDCs and SDCs after challenge with LPS (B) . The results are combined from three independent experiments; each experiment was performed in triplicate. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.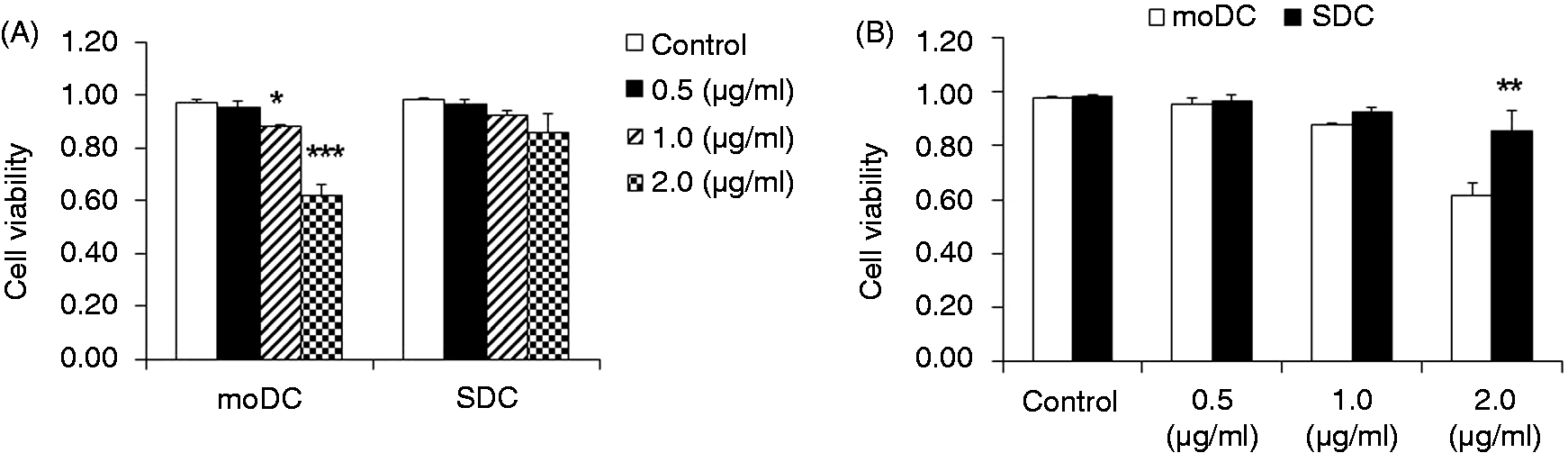
The phagocytic activity of moDCs was higher than SDCs
The phagocytic activity might vary between cell types, as well as during DC maturation; therefore, the difference in phagocytosis between moDCs and SDCs was investigated. Phagocytosis by moDCs was significantly increased in a concentration-dependent manner (Figure 2A). However, in the case of SDCs, phagocytosis activity was significantly increased following treatment with 1.0 and 2.0 µg/ml LPS compared with 0.5 µg/ml LPS (Figure 2A). When moDCs and SDCs were compared, irrespective of LPS concentration, the phagocytic activity of moDCs was significantly higher than in SDCs (Figure 2B).
Comparison of phagocytic activity between moDCs and SDCs. moDCs have stronger phagocytic activity than SDCs stimulated by LPS. moDCs at d 7 and SDC at d 3 were scraped for the phagocytosis assay. moDCs and SDCs were incubated with fluorescein-labeled Escherichia coli K-12 BioParticles after challenge with LPS (control, 0.5, 1.0 and 2.0 μ g/ml) for 4 h. Phagocytosis of moDCs and SDCs stimulated with different concentrations of LPS (A) and comparison of phagocytic activity of moDCs and SDCs under the same concentration of LPS (B). The results are combined from three independent experiments; each experiment was performed four times. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.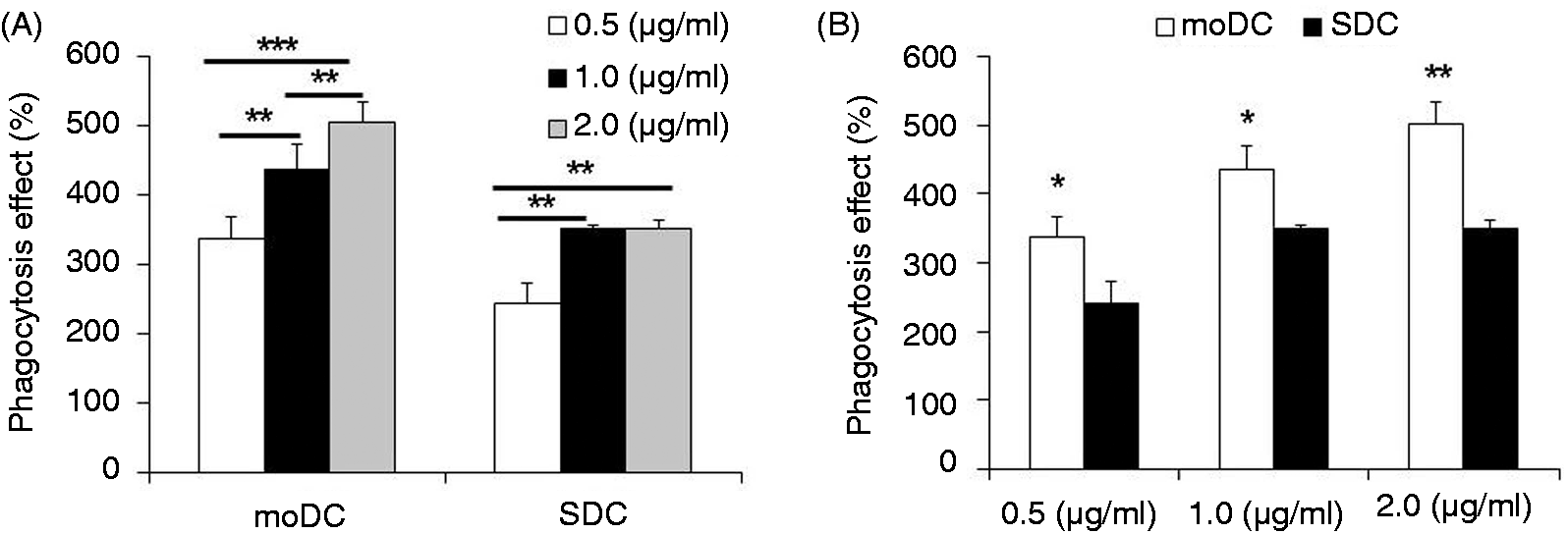
TLR4/MD2 complex-related genes were up-regulated in moDCs and SDCs
To clarify the LPS-stimulated regulation of the TLR4/MD2/LPS complex, associated gene expressions were quantified by qRT-PCR. TLR4 and MD2 mRNA was significantly (P < 0.001) up-regulated 2 h after LPS treatment in moDCs (Figure 3A,B), whereas these genes were increased at 3 h after LPS stimulation in SDCs (Figure 3A,B). When moDCs and SDCs were compared, TLR4 was significantly higher at 1 h and 12 h in moDC compared with SDC, respectively, and MD2 was significantly higher at 1 h and 6 h in moDCs compared with SDCs, respectively. We found that CD14 mRNA expression was undetectable for 12 h after LPS stimulation in moDCs (Figure 3C). The Ct value was more than 36 cycles, which was considered to be undetectable. CD14 mRNA expression was significantly increased at 6 h after LPS stimulation, and remained significantly higher (P < 0.01) in SDCs (Figure 3C). However, it was significantly increased at 12 h after LPS stimulation (P < 0.001), and remained higher in moDCs (Figure 3C). Notably, CD14 mRNA expression was higher in SDCs at 6 h compared with moDCs, whereas it was higher in moDCs at 24 h compared with SDCs (Figure 3C).
The effect of LPS stimulation on the TLR4/MD2/CD14 complex and the mRNA expression of downstream genes. The effect of LPS stimulation on the TLR4/MD2/CD14 complex and mRNA expression of downstream genes in moDCs and SDCs stimulated with LPS (1.0 μl/ml) for the indicated times (0, 1, 3, 6, 12, 24 h). moDCs at d 7 and SDCs at d 3 were treated or not LPS. The mRNA expression of TLR4, MD2, CD14, MyD88, TRAF and IRF3 was quantified by qRT-PCR after treatment. (A–C) TLR4/MD2/CD14 complex; (D, E) TLR4/MyD88-dependent; (F) MyD88-independent pathway-related gene expression between moDCs and SDCs subtypes in a cell type-dependent and time-dependent manner. The results are combined from three independent experiments; each experiment was performed in triplicate. The data are represented as the mean ± SD. The letters x (P < 0.05), y (P < 0.01) and z (P < 0.001) indicate the significant differences of a gene expression between moDCs and SDCs at a specific time point. *P < 0.05, **P < 0.01 and ***P < 0.001 indicate the time-dependent TLR4/MD2/CD14 complex and downstream mRNA expression in moDCs and SDCs between the LPS stimulation group and LPS control group.
Comparison of NF-kB pathway activation between moDCs and SDCs
In order to further clarify the pathway followed by the DCs subsets, we determined the related MyD88-dependent and MyD88-independent gene expression in moDCs and SDCs at different times. To estimate the involvement of MyD88-dependent pathways, we quantified MyD88 and TRAF6 gene expression. We found that both genes were up-regulated in both cell types, and expression of MyD88 was significantly increased from 3 h of LPS stimulation onwards (Figure 3D), whereas expression of TRAF6 was significantly increased from 6 h onwards in SDCs. The expression of TRAF6 was increased only at 12 h on moDCs (Figure 3E). Notably, the MyD88 gene was expressed more highly in moDCs than in SDCs after 3 h of LPS treatment (Figure 3D).
SDCs responded to LPS stimulation more via the TLR4/MyD88-independent pathway
Unlike the MyD88-dependent pathway, the MyD88-independent pathway is activated through IRF3 activation. We found that IRF3 mRNA was significantly increased in SDCs 6 h after LPS treatment and increased steadily up to 24 h (Figure 3F). Interestingly, IRF3 gene expression in LPS-treated moDCs was not different to the untreated control (Figure 3F).
LPS induced moDC maturation more strongly than SDCs
To study the effect of LPS on the maturation state of DC subsets, we examined the production of co-stimulatory molecules CD86 and CD40 on the porcine moDCs and SDCs. Flow cytometric analysis showed that the expression of CD86 and CD40 was significantly increased in moDCs after LPS stimulation compared with SDCs (Figure 4). However, when control group cells were compared with LPS-treated cells, the expressoion of both co-stimulatory markers was increased in LPS-stimulated DCs, indicating that both moDCs and sDCs were responding to the LPS stimulation. CD86 production was significantly up-regulated in moDCs compared with SDCs (Figure 4C). In contrast, CD86 and CD40 were expressed more in SDCs than in moDCs (Figure 4A–C), demonstrating that moDCs were more immature than SDCs.
LPS-induced moDC and SDC maturation. moDCs were harvested on d 7 as adherent immature DCs; adherent SDCs were harvested on d 3. The isotype control of moDCs were from the 7-d immature moDCs and SDCs from the 1-d adherent CD11c+ DCs (data not show). moDCs and SDCs treated or not with LPS (1.0 µg/ml) for 24 h. CD86 (A) and CD40 (B) levels were measured by flow cytometry. (C) Quantification of the parent percentage of mean. The results are combined from two independent experiments; each experiment was performed in duplicate. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.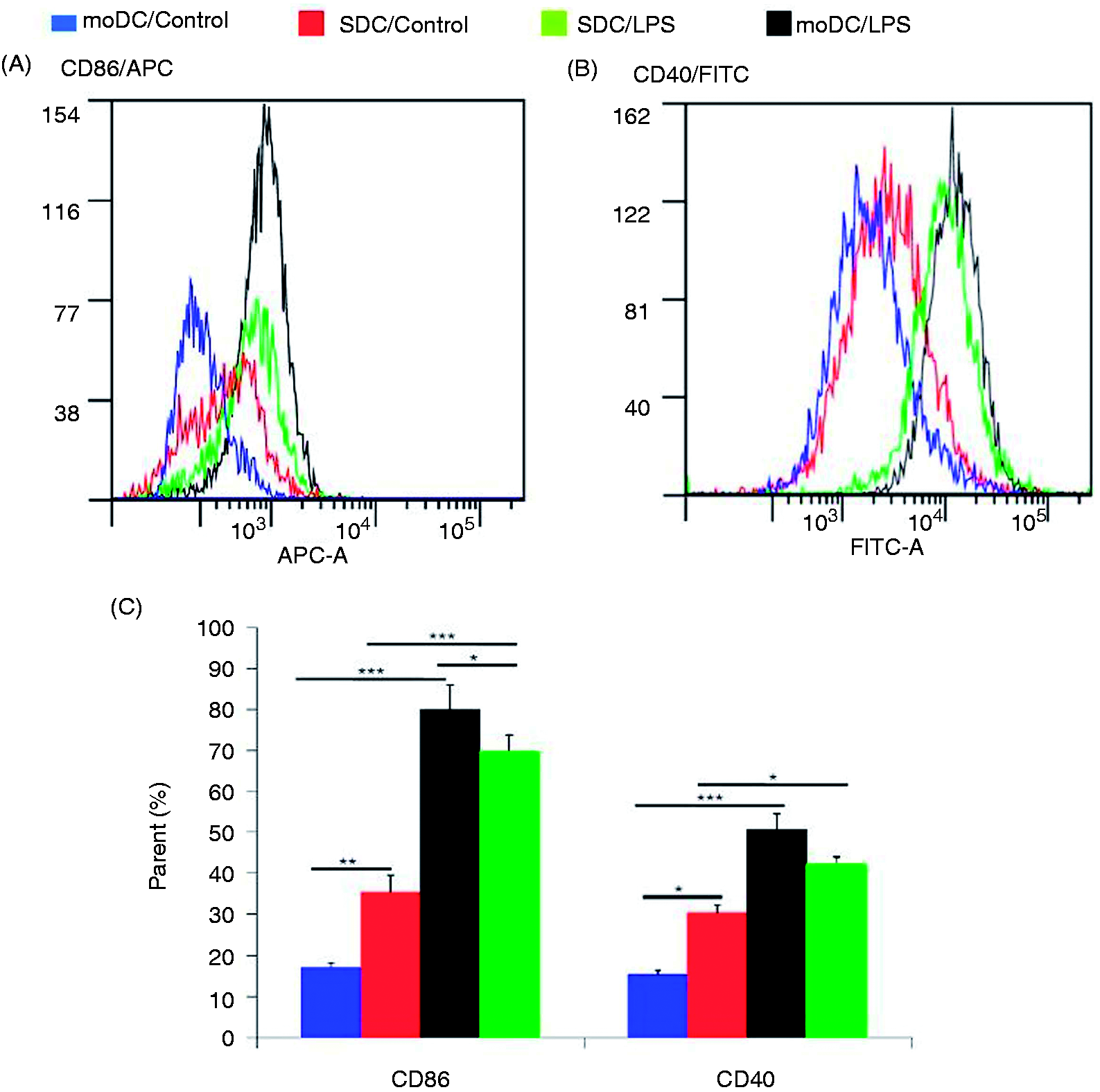
Production of pro-inflammatory cytokines was higher in moDCs than in SDCs
mRNA and protein expression of the cytokines was compared between porcine moDCs and SDCs following LPS stimulation. TNF-α mRNA expression was significantly increased in moDCs from 1 to 12 h after LPS stimulation (Figure 5A). Additionally, TNF-α protein production was remarkably higher in moDCs from 6 to 24 h after LPS stimulation compared with SDCs (Figure 5B). In contrast, IL-1β mRNA expression and protein secretion were significantly higher in SDCs at 1 and 3 h, whereas both mRNA and protein expressions were remarkably up-regulated in moDCs at 12 and 24 h (Figure 5C,D) after LPS stimulation.
Expression of the pro-inflammatory cytokines TNF-α and IL-1β, and the chemokine IL-8 in moDCs and SDCs in response to LPS. mRNA and cell culture supernatants were used from the same sample as the TLR4 pathway gene expression. (A, C, E) TNF-α, IL-1β and IL-8 mRNA expression was quantified by qRT-PCR. The results are combined from three independent experiments; each experiment was performed in triplicate. (B, D, F) The corresponding protein secretions were quantified by ELISA. The results are combined from two independent experiments; each experiment was performed in duplicate. The values show the differences between moDCs and SDCs in a (A, C, F) cell subtype-dependent mRNA and (B, D, E) protein expression manner. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.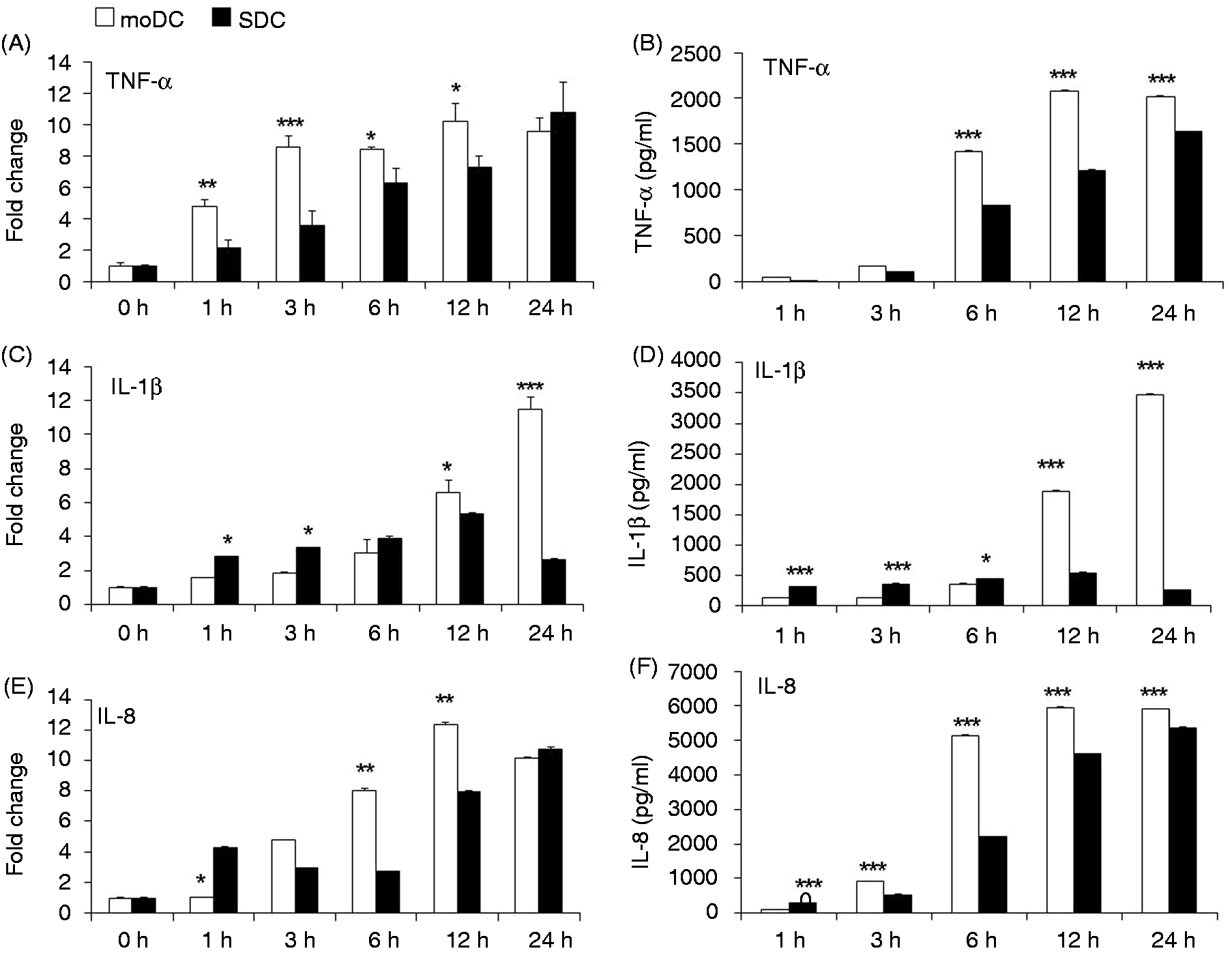
Chemokine secretion differences between moDCs and SDCs
Our data showed that IL-8 mRNA expression and protein secretion were significantly up-regulated in SDCs compared with moDCs at 1 h after LPS stimulation (Figure 5E,F). On the contrary, after 6 and 12 h, IL-8 mRNA and protein expression was significantly up-regulated in moDCs compared with SDC (Figure 5E,F). Importantly, IL-8 protein production was remarkably higher in moDCs compared with SDCs at 3 h, and remained higher until 24 h (Figure 5F) after LPS stimulation.
SDCs produce more IL12-p40 and IFN-γ than moDCs
We determined the protein expression of the auto-inflammatory cytokines IL-12p40 and IFN-γ. IL-12p40 mRNA expression was significantly up-regulated in SDCs at 6, 12 and 24 h after LPS stimulation compared with moDCs (Figure 6A). Likewise, protein secretion was also remarkably higher in SDCs at 3, 6, 12 and 24 h after LPS stimulation compared with moDCs (Figure 6B). After stimulation with LPS, mRNA and protein expression of IL-12p40 remained steady over the study period (Figure 6A,B). mRNA expression of IFN-γ was significantly higher in SDCs at 1 h compared with moDC, whereas it was significantly higher in moDCs at 3 h and 24 h compared with SDCs (Figure 6C). With regard to protein expression, IFN-γ production was higher at 6 h in moDCs than in SDCs (Figure 6D).
Expression of the auto-inflammatory cytokines IFN-γ and IL-12p40 in moDCs and SDCs in response to LPS. mRNA expression of (A) IL-12p40 and (C) IFN-γ was quantified by qRT-PCR. The results are combined from three independent experiments; each experiment was performed in triplicate. (B, D) The corresponding protein secretions were quantified by ELISA. The results are combined from two independent experiments; each experiment was performed in duplicate. The values show the differences between moDCs and SDCs in a cell subtype-dependent (A, C) mRNA and (B, D) protein expression manner. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.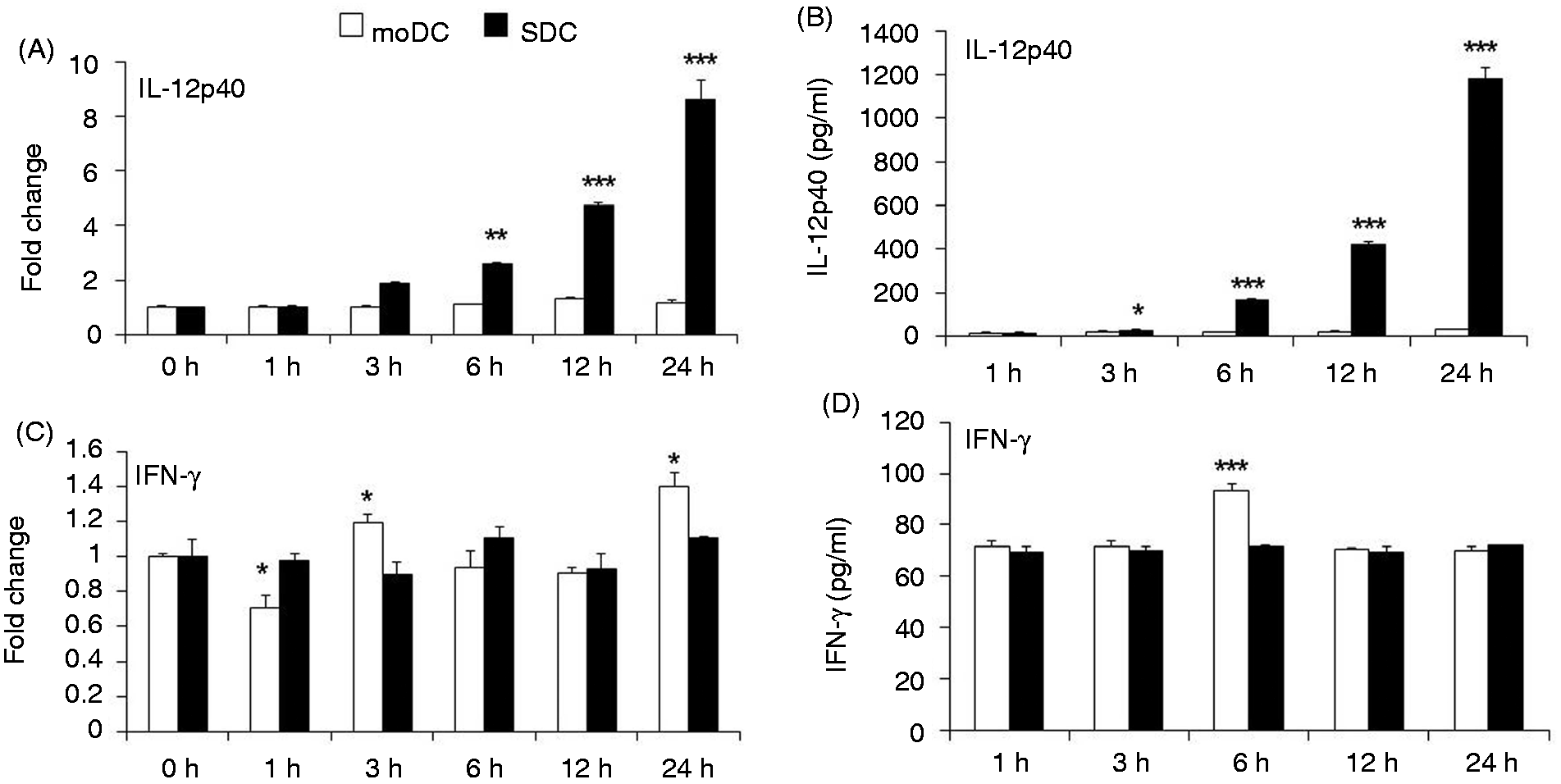
Enhanced production of pro-inflammatory cytokines by moDCs
We determined production of the pro-inflammatory cytokines TNF-α, IL-1β, IFN-γ and IL-12p40, and the chemokine IL-8 by moDCs and SDCs in response to LPS stimulation at various time points. In order to understand cytokine activity over the study period in porcine moDCs and SDCs following LPS stimulation, we used the least square mean multiple comparison test.
26
For this analysis, the protein expression of all the cytokines were used (Figures 5 and 6). We found that the secretion of TNF-α, IL-1β and IL-8 were significantly higher in moDCs than in SDCs (Figure 7). IFN-γ was found to be expressed remarkably more highly in moDCs compared with SDCs (Figure 7). Unexpectedly, IL-12p40 was remarkably increased in SDC (Figure 7), although it was not influenced by LPS stimulation in moDCs (Figure 6B).
Differences in production of cytokines and chemokines in moDCs and SDCs. Data are from the cytokine and chemokine results described in Figures 5 and 6. We used the least square mean multiple comparison test to analyse data in order to study the differences of protein secretion between moDCs and SDCs following LPS stimulation at various time points. The values show the differences between moDCs and SDCs in a cell subtype-dependent protein expression manner. The data are represented as the mean ± SD. *P < 0.05; **P < 0.01; ***P < 0.001.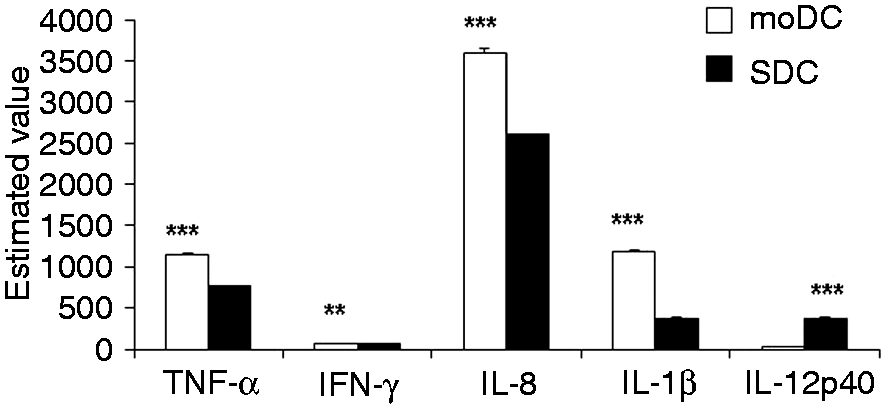
Discussion
moDCs are more susceptible to LPS and exhibit a more mature phenotype than SDCs
Phagocytosis is reported to be an important component of the immune system contributing to the first line of defence. 27 Phagocytosis involves the recognition and binding of prey receptors through PAMPs on the surface of phagocytes, in particular to TLR and phagocytic receptors. Foreign microbes, such as bacteria (LPS) or fungi, can be recognised and taken up from infection sites by professional phagocytes, such as macrophages and DCs. DCs are the key producers of phagocytic receptors, and present Ags as major APCs. 28 DCs have quite different functional outcomes, depending on their class. In this study, porcine moDCs and SDCs were exposed to various concentrations of LPS. We found that moDCs show more potential phagocytic activity after a slight increase of LPS concentration compared with SDCs. This indicates that phagocytosis is enhanced in an in vitro culture model of moDCs. Interaction with bacteria leads to the apoptotic death of phagocytes during bacterial phagocytosis and degradation stages. Indeed, there is a significant decrease in the viability of moDCs with an increasing concentration of LPS. On the basis of phagocytosis and cell viability stimulation of moDCs and SDCs stimulated with various concentrations of LPS, we assumed that 1.0 µg/ml was suitable for further study. It is well known that immature and mature DCs have a distinct morphology, phenotype and function. Importantly, the maturation of DCs is shown by up-regulation of CD40 and CD80/86 during antigenic stimulation. 29 CD28 is the most important co-stimulatory molecule on the naïve T-cell surface to clonally expand and acquire effector functions, such as control of the T-cell cycle, survival and differentiation. 30 It is well-described that CD80/86 through CD28 increase pro-inflammatory cytokine production in T-cells. 31 CD86 was found to be up-regulated on the cell surface of DCs, suggesting that it interacted with CD28, thereby promoting T-cell activation and maturation. In addition, the expression of CD40 on DCs is important for T-cell priming and T-cell-mediated effector function. 32 LPS-stimulated human or mice moDCs display a strong expression of the maturation markers CD40 and CD86,31,33 which is in agreement with this study. We identified that the co-stimulatory molecules were up-regulated in both moDCs and SDCs, whereas moDCs were induced to be more mature than SDCs following LPS stimulation. Up-regulation of the co-stimulatory molecules in response to LPS stimulation indicated that moDCs display a more mature phenotype and T-cell activation than SDCs.34 Immature DCs exhibit higher phagocytic activity than mature DCs and perform specialised tasks in Ag presentation, 35 which is in good agreement with this study. However, in humans and mice, it has been reported that immature DCs are immunologically more active with different cell functions than peripheral circulating waves of tissue-resident DCs. 36 The differences between these studies could be a result of using different DC cell subsets (moDCs, SDCs or cord blood DCs).
Comparison of TLR4 signalling pathway activation between moDCs and SDCs
DCs have high phagocytic activity and remove free microorganisms in the host innate immune system through PRRs, especially TLRs, that bind to PAMPs. 8 Importantly, one of the essential roles of DCs, like other host immune cells, is initiation of the pro-inflammatory response by the regulation of TLR activation and cytokine/chemokine secretion. 37 Mounting evidence reveals that LPS–TLR4 interaction is important in the clearance of bacteria by up-regulating the phagocytic activity of DCs via these two distinct pathways. 38 We have found that LPS up-regulates TLR4 steady-state transcripts in porcine DCs, which is in very good agreement with other studies of monocytes, macrophages and PBMCs.21,39,40 LPS has been reported to trigger mitogen-activated protein kinases, NF-κB, STAT3, IRFs and activator protein signal-transduction pathways by the activation of the TLR4 receptor complex (TLR4/MD2), which evokes the transcription and production of immune genes, including cytokines that are critical for activation of innate immunity.41,42–44 In this study, we found that increasing TLR4 and MD2 expression was detected in both types of DCs, with higher phagocytic activity, coinciding with previous reports. 45 Similar to the other studies in mice and humans, we demonstrated that the TLR4/MD2 complex genes were up-regulated in porcine moDCs and SDCs in a time-dependent manner in response to LPS stimulation. 41 TLR4 and MD2 combine into a LPS receptor molecule complex at both transcriptional and translational levels. Therefore, the differences between the expression of these two genes indicates that both DCs subtypes could respond to pathogens during different inflammatory phases. Additionally, we observed that moDCs have a more sensitive TLR4 receptor complex to recognise LPS following stimulation, and are also more efficient in the induction of T-cell differentiation and development. One of the most important characteristics of TLR4/MD2 complex activation is the up-regulation of the pro-inflammatory cytokines that activate naïve T-cells. 46
CD14 is the key cell surface protein required for the proper functioning of monocytes and DCs. 47 Unlike in SDCs, CD14 mRNA was undetectable in moDCs during the early stages of LPS stimulation. This finding is supported by a previous study, which reported that, during the differentiation of monocytes to moDCs, the cell surface expression of CD14 was silenced in vitro. 48 CD14 protein polymorphisms determine the susceptibility of individuals to microbial infection. 47 Additionally, it has been reported that LPS can induce the TLR4 pathway independently of CD14, allowing activation of the anti-inflammatory response. 49 Therefore, we postulate that moDCs are more potential immune cells and that their activation is flexible to different pathways. It is worthwhile mentioning that, after LPS stimulation, moDCs displayed considerably stronger TLR4/MyD88-dependent pathway activation, but were less likely to induce the TLR4/TRIF-dependent pathway. The application of LPS increased the potentiality of moDCs to bacterial infection and showed higher immune responses with a remarkable increase in the production of the anti-inflammatory cytokines. Moreover, we also observed that LPS not only induced the maturation of moDCs more efficiently, but also enhanced phagocytosis and migration.
moDCs are more efficient than SDCs in producing inflammatory cytokines and activating the NF-kB signalling pathway
Following LPS-induced TLR4 activation, MyD88-dependent and MyD88-independent (or TRIF-dependent) pathways are involved for the further modulation of anti-inflammatory responses. 37 Activation of MyD88 coupled to TLR4 activates TRAF6 and then rapidly activates NF-κB, resulting in the production of pro-inflammatory cytokines and chemokines, such as TNF-α, IL-1β and IL-8.50,51 This study identified that LPS, after binding to the TLR4/MD2 complex, induced pro-inflammatory cytokine expression at both the mRNA and protein levels in moDC and SDCs, and that these responses were more pronounced in moDCs than in SDCs. It has been reported that LPS-induced DC maturation is associated with the up-regulation of co-stimulatory molecules, such as MHC and CD86, which is dependent on NF-κB activation. 52 Indeed, delaying the addition of LPS to moDCs and SDCs abrogated MyD88-mediated up-regulation of LPS-induced TNF-α, IL-1β and IL-8 secretion. Activation of LPS-induced TNF-α, IL-1β and IL-8 mRNA expression by LPS requires protein synthesis, which we found in this study; specifically, expression of these molecules was more rapidly induced in moDCs than in SDCs. A vast amount of data indicate that TNF-α induces the maturation of immature DCs and, moreover, that it induces transcellular migration of DCs, which depends on the production of IL-8.53,54 The migratory DCs activate T-cells and subsequently initiate the adaptive immune response against invading bacteria and viruses. 55 However, recruitment of TRIF associated with TLR4 results in the activation and nuclear translocation of IRF3, an essential transcription factor generally involved in induction of type I IFN and pro-inflammatory cytokines, such as IL-12, IL-18 and IL-23, in wild type DCs. 14 Additionally, IRF3 is necessary for the early phase of phagocytosis. 56 We found, in this study, that LPS-induced TLR4/TRIF-dependent signalling pathway activation constitutively expressed IRF3 in SDCs, which coincides with the results of another study. 50 We found that IRF3 was uninfluenced in moDCs following the LPS stimulation. This finding indicates that the TLR4/TRIF-dependent signalling pathway might be not involved in the LPS-induced activation of moDCs.
Besides the positive features of the moDCs, we also observed some unexpected findings; that is, that moDCs did not satisfactorily release IL-12p40 or IFN-γ following 24 h of LPS stimulation. IL-12p40 plays a critical role in Th17-cell-mediated chronic inflammation by promoting Th17 cell maintenance and by the production of pro-inflammatory cytokines. 57 It also plays an important role in DCs for IFN-γ production. Expression of IL-12p40 is required for the generation of CD4 T-cells and DC migration, which, in turn, regulates the level of IFN-γ produced after microbial infection. 58 The IFN-β subunit is involved in signal transduction and interrupts the IL-12 receptor binding sites, leading to insufficient production of IFN-γ. 59 Indeed, IL-12p40 was strongly released by SDCs at different time points following LPS stimulation, which might affect the secretion of IFN-γ. Nevertheless, this increased production of IL-12p40 in SDCs could not overcome the more potential immune activation of moDCs.
Conclusions
Taken together, our results show that LPS induced activation, maturation and phagocytosis of porcine DCs, and that the level of activation of moDCs was greater than that observed in SDCs. The moDCs were also more efficient than SDCs in releasing pro-inflammatory cytokines after LPS stimulation. Higher levels of cell proliferation, differentiation and IL-1β production were observed in moDCs compared with SDCs. Pro-inflammatory cytokines were produced more abundantly in moDCs, which may result in more efficient activation of the NF-κB signalling pathway and T-cell maturation compared with SDCs. The interesting finding of this study is that LPS more strongly activated the TLR4/MyD88-dependent pathway in moDCs. These results indicate that the generation of porcine moDCs in vitro developed an appropriate DC culture model allowing the study of DC roles in the host defence system.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial or not-for-profit sectors.
Acknowledgments
We thank Ludger Buschen for animal husbandry; Birgit Koch-Fabritius and Helga Brodeßer for technical assistance; Elmar Endl, Andreas Dolf, Peter Wurst and the flow cytometry facilities for analysis (Institutes of Molecular Medicine and Experimental Immunology, University of Bonn); Nicole Krämer for phagocytosis measurement [Life and Medical Science (LIMES) Institute, Program Unit Chemical Biology and Medicinal Chemistry, University of Bonn]; Christine Große-Brinkhaus for data analysis; and Christiane Neuhoff, Rui Zhang and Mohammad Ariful Islam for sample collection. In particular, we would like to acknowledge the China Scholarship Council for supporting Xueqi Qu.
Conflict of interest
The authors do not have any potential conflicts of interest to declare.