
Abstract
TLRs represent one of the most important components of innate immunity. Currently, these receptors have been extensively studied in vertebrates and insects, but our knowledge for annelids is very limited. Therefore, the aim of our study was to identify earthworm TLR homologs by genome-wide screening, and to determine the expression of candidate genes as a response to Gram-positive and Gram-negative bacteria. Using a combination of deep pyrosequencing and quantitative PCR we found six candidate genes, for which all were expressed in Dentrobaena veneta. Two of the candidates showed significant response to bacterial exposure. In conclusion, TLRs seem to have a role in earthworm immunology.
Introduction
Invertebrates have a simple immune system, recognizing harmful or pathogenic microorganisms mainly through innate PRRs.1–3 The PRRs recognize PAMPs associated with the invading microorganisms. PRRs are generally conserved throughout evolution, with TLRs being the most widespread. 4 These have an architecture consisting of extracellular leucine-rich repeats where the extracellular domains transmit the sensed signals through a cascade of adaptor molecules to activate a range of defense mechanisms. However, we still know very little about the role of TLRs in annelids,3,5 although they have been extensively studied for vertebrates—in which several families and sub-families have been identified 6 —and insects in which the TLRs were originally detected.7,8
Earthworms are oligochaetal annelids living in soil. 9 Soil represents a microbiologically hostile environment, with > 20,000 microbial genome equivalents. 10 Despite their simple immune system, earthworms utilize the soil environment as habitat. 11 TLRs could be an important component in doing so.
Therefore, the aim of this work was (i) to identify candidate TLRs in the earthworm Dendrobaena veneta by genome-wide screening, and (ii) to determine the expression patterns of TLR homologs as a consequence of exposure towards both Gram-positive and Gram-negative bacteria.
Our strategy was to first identify TLRs in the D. veneta genome by low coverage genome sequencing. Further, we investigated the immune cell (coelomocyte) expression of these as a consequence of exposure to Staphylococcus aureus (as a candidate Gram-positive bacterium) and Escherichia coli (as a Gram-negative candidate). The rationale for investigating coelomocytes is that these are the main targets for TLR expression and immunological responses. 12
We present results showing gene expression responses of the TLR candidates to bacterial exposure, supporting their role in earthworm immunology. 5
Materials and methods
Earthworm culture
The experiments were conducted with the earthworm species D. veneta. This species was chosen because it is easy to access through the largest earthworm producer in Scandinavia (Fibe AB, Őverkalix, Sverige). Soil micro-ecosystems with a culture of the earthworm were created in specially-designed containers (36 × 51 × 18) cms with a mesh allowing free oxygen access (Magic Products, Amherst Junction, WI, USA). We used a commercial earthworm soil consisting of a mixture of an organic fiber and Canadian sphagnum peat moss that was kept at 80% moisture (Magic Products). Every week, 1 g of Magic Worm Food (Magic Products) in water slurry was added per earthworm. The food was composed of around 12% protein, 1% fat, 81% carbohydrates and 6% fiber (www.magicproducts.com).
Earthworm genome sequencing and analysis
The anterior part of a single earthworm was subjected to genome sequencing. Total DNA was isolated using the Qiagen tissue kit following the manufacturer’s recommendations (Qiagen, Hilden, Germany). The genomic DNA was prepared according to the manufacturer’s instructions for 454 sequencing of total DNA and subsequently sequenced using the Genome Sequencer FLX Titanium System (Roche, Basel, Switzerland). The genome sequencing was done at the Norwegian Sequencing Center (www.sequencing.uio.no). The sequences were deposited in the National Center for Biotechnology Information (NCBI) database with the ID SRS523963 under the BioSample accession SRP034887.
TLR homologs were identified by BLAST searches in a local database generated from the pyrosequencing reads by the CLC Genomic Workbench (CLC bio). Candidate genes used for the searches were TLR homologs identified from the Lumbricus rubellus genome database (xyala.cap.ed.ac.uk/Lumbribase/; B. Elsworth, Earthworm Genome Sequencing Consortium and M. Blaxter, ‘The Lumbricus rubellus Genome Project’, www.earthworms.org, personal communications). For the identified homologs we used the nomenclature as given to the raw sequencing reads to ease the comparison with the deposited sequencing data.
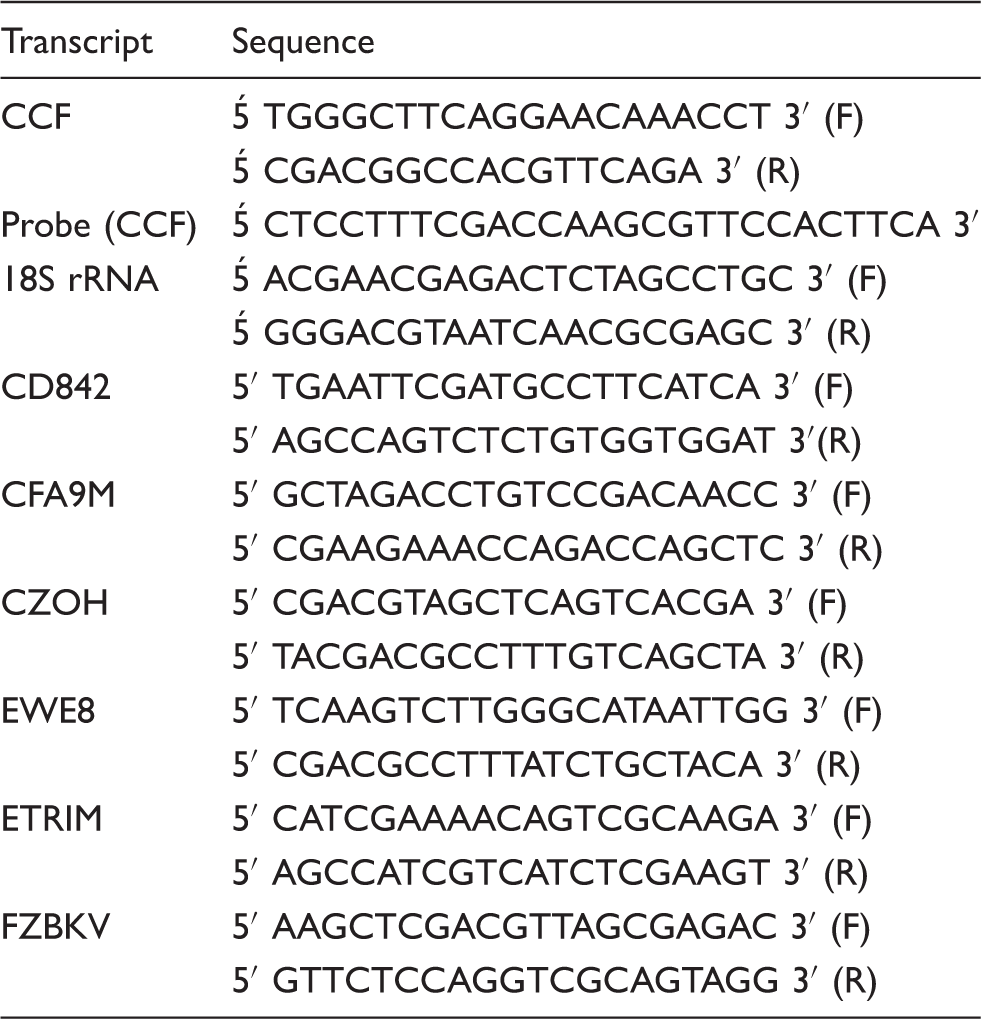
PCR primers
Primers and probes used in this study.
Gene expression experiments
Three main gene expression experiments were conducted: (i) the effect of bacterial exposure, (ii) the effect of starvation and (iii) longitudinal investigation of individual differences. The longitudinal nature of the experiments was both to identify potential confounding gene expression responses, and to investigate individual differences in expression patterns as these could influence results interpretation.
The effect of bacterial exposure was determined by soaking the earthworms in cultures containing 109 CFU/ml of E. coli K-12 or 109 CFU/ml S. aureus. These species were chosen because they represent model Gram-negative and Gram-positive bacteria, respectively. Furthermore, these bacteria have previously been used in earthworm gene expression experiments.17,19,20 Twenty non-exposed earthworms were used as controls, while 80 earthworms were exposed to bacteria (40 to each bacterium). Immune cells were harvested between 2 and 4 d. For the effect of starvation immune cells harvested from 120 earthworms were investigated, that is 20 earthworms from each of 0, 1, 2, 3, 4 and 5 d of starvation on filter paper. Finally, for evaluation of individual stability repeated analyses of 12 individually housed earthworms were conducted after 0, 14 and 28 d.
Immune cells were harvested after extraction of immune cells by electro stimulation of earthworms at 5–10 V for 5 s in a modified Lumbricus balanced salt solution (LBSS) [71.5 mM NaCl, 4.8 mM KCl, 1.1 mM MgSO4 · 7H2O, 0.4 mM KH2PO4, 0.3 mM Na2H2PO4 · H2O and 4.2 mM NaHCO3, pH 7.3 with an osmolarity of 220 milliosmols per kilogram of water (mOsmol/kg) (semi-micro osmometer K-7400, KNAUER, Berlin, Germany)] added 10 mM EDTA and 2% FBS (F7524; Sigma-Aldrich, St. Louis, MO, USA). The immune cells were then precipitated by centrifugation (500 g) for 10 min at 4℃ (Beckman Coulter Allegra 25R Centrifuge, Fullerton, CA, USA), and washed twice using modified LBSS and low-speed centrifugation.
RNA isolation was performed using Qiagen RNeasy Mini Kit on freshly harvested immune cells. Homogenized lysed samples were stored at −80℃ until all samples in the experiment were collected as described by the manufacturer. Frozen lysates was thawed at 37℃ and protocol from the manufacturer was followed for isolation of RNA.
Further, the isolated RNA was treated with DNase1, Amplification Grade (Cat. no. 18068-015; Invitrogen, Carlsbad, CA, USA) before used as template for complementary DNA (cDNA) using reverse transcriptase. cDNA was synthesized using RNase OUT, Ribonuclease Inhibitor (Recombinant), (Cat. no.10777-019; Invitrogen) and SuperScript III Reverse Transcriptase (Cat. no. 18080-044, Invitrogen). The manufacturer’s protocol was followed. A control containing reagents, but no template, was included in the experiments as a negative control.
Amplifications were conducted in a 7500 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using 20 µl reaction volumes containing 1 unit Dynazyme II HotStart DNA polymerase (Thermo Scientific, Waltham, MA, USA), 1 × Dynazyme TM II Hot start reaction buffer (Thermo Scientific), 400 µmol dNTPs (Thermo Scientific), 0.2 × Rox (Life Technologies, Invitrogen) 0.2 µmol probe (Biomers), 0.4 µmol of each primer (Biomers). All amplifications were conducted using 40 cycles with denaturation at 95℃ for 30 s and elongation at 60℃ for 1 min. The primers and probes used are listed in Table 1. LinRegPCR was used to estimate the amplification efficiency, 21 while gene expression was measured relative to the 18S rRNA gene. Statistical testing of the measured expression levels of the identified homologs was done using the Mann–Whitney U-test (SYSTAT v. 13). For all experiments, except the investigation of individual gene expression stability, all calculations were based on comparing different earthworms, while for the individual gene expression stability we performed repeated analyses of the same earthworms.
Results
Identification of TLR homologs
The genome sequencing resulted in a total of 556,684 sequencing reads with an average length of 325 base pairs (bp). The sequences represent 180 million bp. The genome size of D. veneta is about 450 million bp. 22 Our sequencing depth was thus about 0.4-fold. This corresponds to an average number of reads per gene of 1.2 (one gene is on average covered by about three reads assuming a gene size of 1000 bp, while we only analyzed 40% of these).
TLR homologs identified in D. veneta.

The L. rubellus Genome Project.
The sequencing read with the lowest e-value.
Gene expression of TLR homologs
We found relatively large gene expression effects for the TLR gene homologs ETRIM and FZBKV for the earthworms externally exposed to E. coli and S. aureus (Figure 1). Compared with the control ETRIM was down-regulated after S. aureus exposure (P < 0.0005, Mann–Whitney U-test), while FZBKV was up-regulated after E. coli exposure (P = 0.008, Mann–Whitney U-test). Similar patterns were also detected when comparing the exposure to S. aureus and E. coli (P < 0.0005 and P = 0.001, respectively, with Mann–Whitney U-test).
Expression of TLR gene homologs as response to bacterial exposure. Earthworms surface exposed to about 109 CFU/ml were analyzed 2–4 d after exposure. Error bars represent SEM between individual earthworms. **p = 0.01; ***p = 0.001; ****p < 0.001.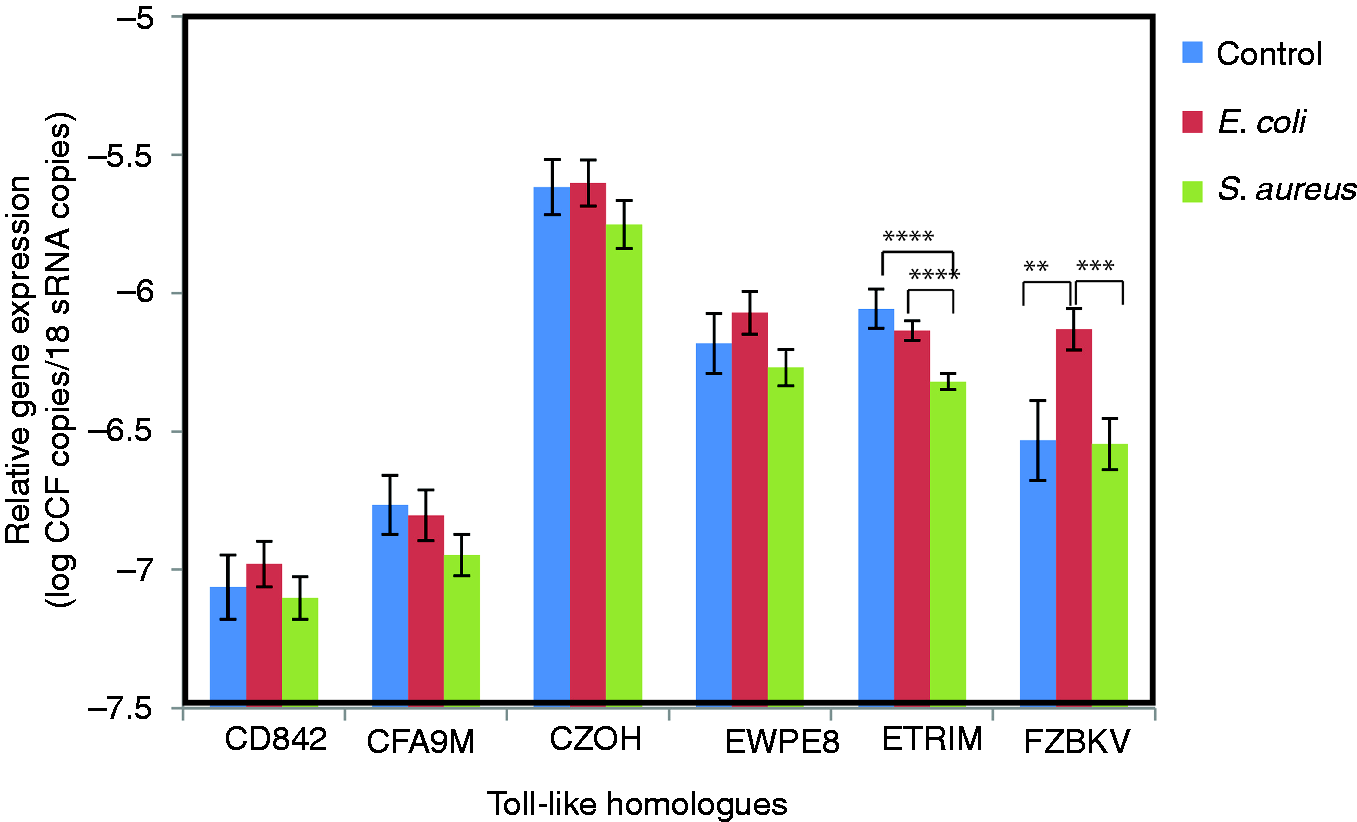
As all experiments were done on starved earthworms we conducted an experiment in which we tested the gene expression effect of starvation over 5 d. We found no effect of starvation for any of the genes investigated (Supplementary Table 1). Finally, we tested whether there were individual differences in the gene expression by analyzing the immune cell expression of the TLR homologs over a period of 3 mo. The testing was repeated three times for 12 earthworms. We did not detect any major individual differences in the expression patterns for any of the TLR homologs (Supplementary Table 2).
CCF gene expression
We could not detect any systematic differences in CCF expression for any of the experimental conditions tested (results not shown). The average CCF expression was about 5 log lower than that of 18S rRNA. What was apparent, however, were stable individual differences in the gene expression patterns over the 28 d investigated, where one earthworm showed a more than 10-fold higher expression than the others (Figure 2).
Individual differences in CCF gene expression. Longitudinal analyses were conducted for 12 earthworms after 0, 14 and 28 d. Error bars represent SEM of three measurements for each earthworm.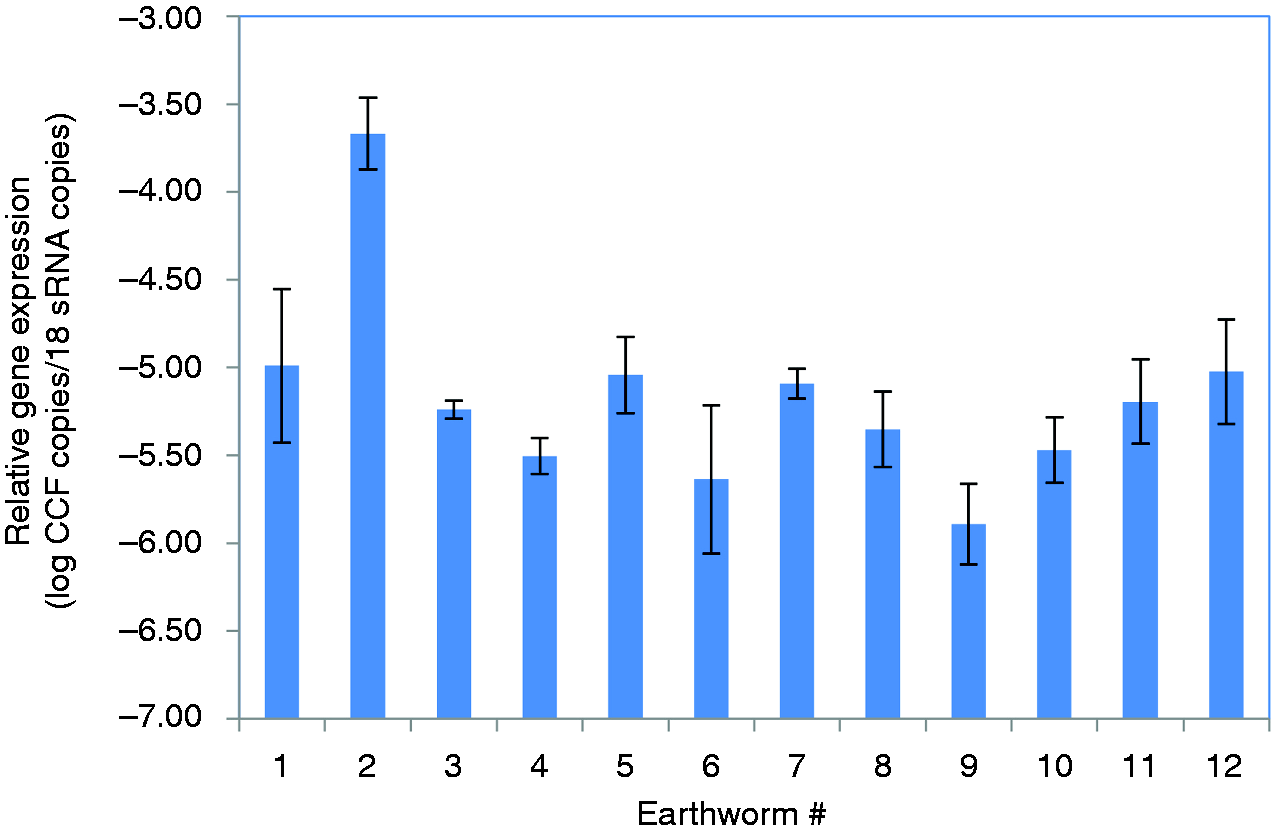
Discussion
The most prominent and systematic gene expression responses were the down-regulation of the TLR 21 homolog (ETRIM) after earthworm exposure to S. aureus, and the up-regulation of the vasorin-like homolog (FZBKV) after E. coli exposure.
TLR 21 has recently been identified in a range of non-mammalian vertebrates. 23 The receptor recognizes CpG oligonucleotides with bacterial origin, and activation requires acidification of endosomes. Staphylococcus aureus is known to escape endosomes. 24 Therefore, the down-regulation of the TLR 21 homolog could be a part of the S. aureus virulence mechanism for endosome escape. Staphylococcus aureus has previously been shown to posess several mechanisms to interfere with other TLR recognitions. 25
Vasorin expression, however, has been shown to be connected to tissue damage. 26 For vasorin homolog expression, the E. coli strain used was a non-virulent laboratory strain so that tissue damage was unlikely. However, owing to the long evolutionary distances, the earthworm homolog does not necessarily have a vasorin-like function. In fact, the closest earthworm homolog from L. rubellus is actually a TLR. Therefore, the vasorin identity may actually be a misclassification.
It has previously been suggested that TLRs play little or no role in annelid immunology. 3 The TLR homolog gene expression response detected by us, however, suggests that TLRs also have a role in annelids. This has also been confirmed by another study identifying a TLR homolog in the earthworm species Eisenia andrei. 5 The identified TLR homolog in E. andrei showed different expression patterns than those identified in D. veneta, supporting a diverse range of TLR homologs in annelids. Potential reasons for why annelid TLR have previously been overlooked could be long evolutionary distances to well-characterized model organisms.
CCF in D. veneta lacks the GlcNAc-recognition domain, which is important for the TNF-α-like properties, 27 such as potential co-induction with TLRs. 28 This may explain the lack of co-induction for the TLRs identified in our work. Interestingly, however, we detected major individual differences in CCF gene expression. The individual differences may indicate a chronic inflammation state for some earthworms, or that earthworms have an immunological bet-hedging strategy with large intrinsic individual variations.
In conclusion, we have shown gene expression responses of TLR homologs in earthworms after microbial exposure and individual variation in CCF expression. These results represent new knowledge about the innate immunology of earthworms.
Footnotes
Funding
This work was financed by Hedmark Sparebank and Hedmark University College.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.