
Abstract
Bactericidal permeability increasing protein (BPI), a 55–60 kDa protein, first reported in 1975, has gone a long way as a protein with multifunctional roles. Its classical role in neutralizing endotoxin (LPS) raised high hopes among septic shock patients. Today, BPI is not just a LPS-neutralizing protein, but a protein with diverse functions. These functions can be as varied as inhibition of endothelial cell growth and inhibition of dendritic cell maturation, or as an anti-angiogenic, chemoattractant or opsonization agent. Though the literature available is extremely limited, it is fascinating to look into how BPI is gaining major importance as a signalling molecule. In this review, we briefly summarize the recent research focused on the multiple roles of BPI and its use as a therapeutic.
Keywords
Introduction
Every living organism has developed an immune system to protect itself from diseases. Microorganisms, such as bacteria, have evolved enzymes that protect them from bacteriophage infection. Eukaryotes have evolved a defence system, collectively termed as the immune system. The basic/primitive defence mechanisms include phagocytosis, killing by antimicrobial peptides/proteins and generation of toxic moieties, such as reactive oxygen and nitrogen species. The presence of antimicrobial peptide/proteins has also been established in both prokarya1,2 and eukarya. 2 The antimicrobial peptide/proteins produced by bacteria are termed as bacteriocin and those produced by eukaryotic group are termed as antimicrobial peptides (AMP). The role of AMPs from phagocytes in the antimicrobial defence was first demonstrated by Hirsch, where the binding of phagocytin to bacteria was identified.3–5 In 1963, Spitznagel and Chi identified phagocytin as a group of cationic antibacterial proteins that are associated with the cytoplasmic granules in the phagocytes and they interact with the phagocytosed bacteria. 6 Later, Spitznagel and Zeya demonstrated that these proteins are bactericidal, damaging the bacterial permeability barrier.7–9 In 1975, Weiss et al., purified a 50–60-kDa cationic protein from granules of polymorphonuclear leucocytes (PMN) and termed it bactericidal/permeability increasing protein. 10 Later, it was revealed that AMPs are also produced by other cells, for example hepatocytes, and epithelial and fibroblast cells.
Generally, most of the AMPs carry a net positive charge, enabling them to attack the negatively-charged (outer) membranes of bacteria, especially Gram-negative bacteria. 11 Eukaryotic cell membranes do have negatively-charged phospholipids, but they are secluded in the inner leaflet, rendering AMPs ineffective against eukaryotic membrane. 12 Considering their bactericidal action they are especially secreted/produced by the cells at the sites (e.g. mucosa, skin, etc.) normally exposed to the bacteria, and by the blood cells, neutrophils, eosinophils, macrophages and platelets. 12 The role of AMPs is not restricted to bacterial killing. They act as chemoattractants, cell-growth promoters and opsonins, and bind to endotoxin (LPS), either to detoxify it or to present it to the cells to induce an inflammatory response. The AMPs that bind to LPS play a crucial role by mediating the signals from Gram-negative bacteria to the innate immune effectors. 12 The two major proteins belonging to this category are LPS binding protein (LBP) and bactericidal/permeability increasing protein (BPI). Both these proteins bind to LPS, but have opposite effects. In this review, we give a recent and brief overview of various aspects of BPI and its relative proteins, including its action, regulation and its role in various diseases.
BPI and its relatives
BPI is a member of a large family of proteins known as the lipid transfer (LT)/LBP family, whose members share an ability to bind a variety of lipid substrates. The members within the family are thought to be evolved by gene duplication and the fact that all the members have an overall similarity in the exon structure shows that they are evolutionarily related.13,14 The different members in the family include proteins that are important in innate immune responses, such as PLUNC (Palate, Lung, and Nasal epithelium Clone), LBP and proteins that act as lipid transporters, such as cholesteryl ester transfer protein (CETP) and phospholipid transfer protein (PLTP). All the members in the family except CETP, which is the most divergent, are located on chromosome 20q. 15 CETP is located on chromosome 16. 16
LBP is a plasma protein that catalyses the transfer of LPS to other LPS binding proteins on the cell surface. Even though LBP and BPI belong to the same family, they are antagonistic in their function. 17 BPI is anti-inflammatory in function, whereas LBP induces an inflammatory response. PLUNC form a group of seven proteins that are encoded by adjacent genes on chromosome 20q11. 13 Members within the family belong to either the short PLUNC (SPLUNC) group or large PLUNC (LPLUNC) group depending on their size. 13 Sequence identity within the group is less, ranging from 16% to 28%. 13 The sequence similarity of PLUNC to BPI is difficult to predict, but structurally they are similar. The amino terminal domain of LPLUNC and SPLUNC are highly variable in their secondary structure. The amino terminal of SPLUNC and both the amino and carboxy terminal domain of LPLUNC are structurally similar to that of BPI (their secondary structure).13,18,19 Like BPI, PLUNC proteins exhibit anti-inflammatory and antibacterial action.20–23 However, the direct role of PLUNC as an antimicrobial is controversial.24–26 PLUNC has been shown to act as a surfactant, reducing the surface tension of airway secretion and disrupting Pseudomonas aeruginosa biofilm formation in vitro. 27
Mode of action
BPI is a molecule of interest among the group of antimicrobial peptides because its mode of action is not just restricted to bacterial killing. BPI has been shown to have various properties, for example antibacterial, anti-inflammatory, opsonizing and anti-angiogenic (Figure 1).
Mode of action of bactericidal/permeability increasing protein (BPI).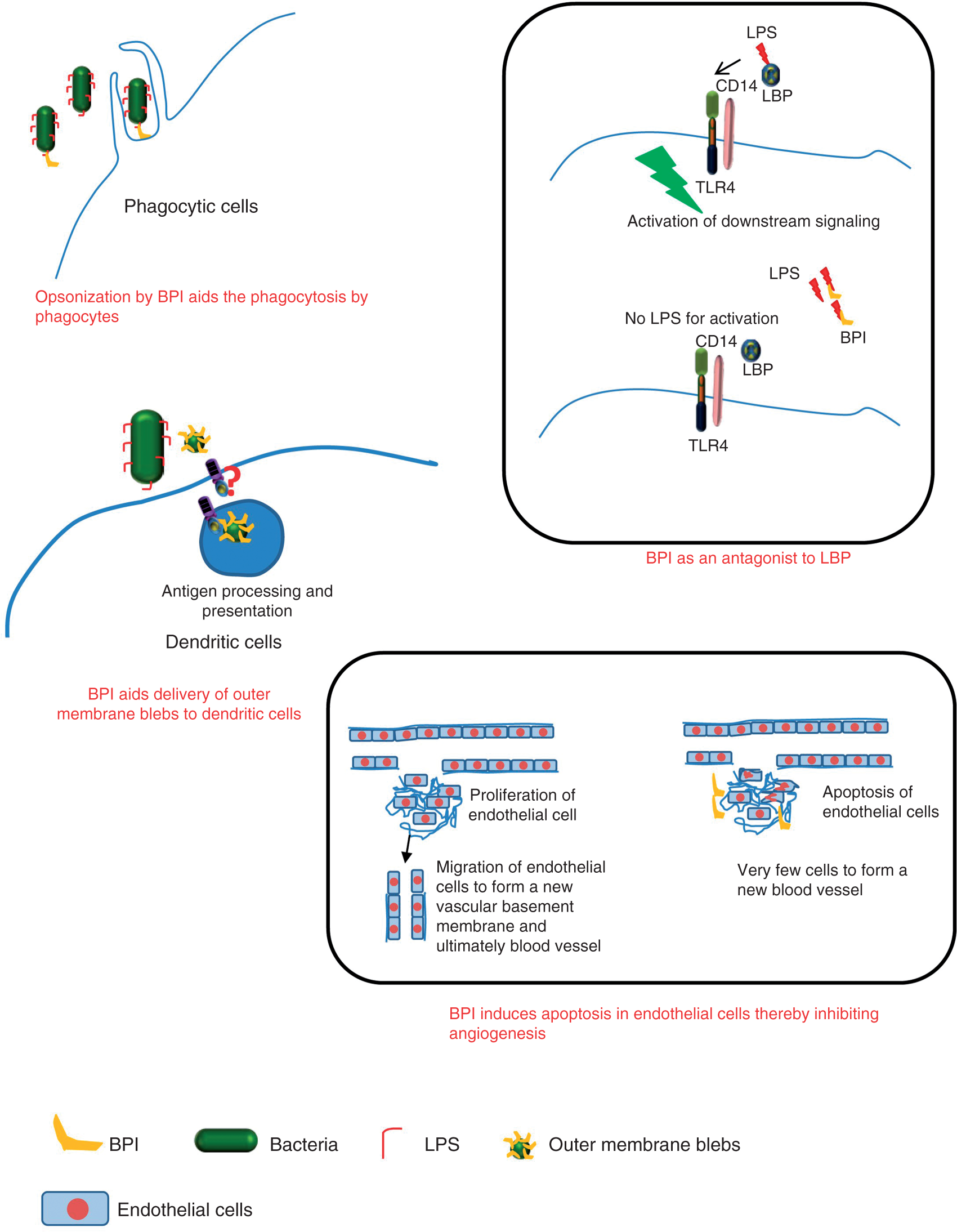
LPS from Gram-negative bacteria is the major determinant of inflammation and also of endotoxic shock/sepsis. BPI possesses the ability to neutralize the effects caused by LPS. 28 It reduces LPS-mediated neutrophil stimulation, 29 pyrogenicity and TNF-α production induced by LPS. 28 BPI neutralizes the effects of LPS by binding to the lipid A moiety in LPS.30,31 This property of BPI is also crucial for its antimicrobial activity. 32 On binding to LPS in the outer membrane, BPI penetrates into the inner membrane in a time-dependent manner, which leads to membrane damage and, ultimately, bacterial cell death. 32 Studies using recombinant BPI showed that prophylactic treatment of monocytes with BPI inhibits LBP-mediated delivery of LPS to monocytes. 31 In vitro studies on rBPI have shown that a 3-fold molar excess of BPI to LBP can prevent LPS-mediated stimulation of monocytes. 31 However, a 10-fold molar excess was essential to completely overcome the action of LBP. The role of BPI has also been demonstrated in opsonization. 33 BPI enhances the delivery of blebs derived from the outer membrane of Gram-negative bacteria to dendritic cells (DC). 34 This can aid LPS uptake and antigen (from outer membrane) presentation by DC. 34 BPI was also found to block the maturation of DCs. 35
LBP, a functional antagonist of BPI, disperses LPS aggregates and delivers the monomeric LPS to CD14 molecules.17,36 BPI acts antagonistically by stabilizing the aggregates thus inhibiting the LBP-mediated delivery of LPS to CD14 molecules. 17
All these different properties of BPI can be correlated directly to its structure. BPI is a 456-residue protein with a boomerang-shaped structure 37 consisting of two domains of comparable length separated by a proline-rich sequence. The amino and carboxy terminal regions both have a lipid binding pocket. BPI shows around ∼50% percent sequence identity to that of LBP.38–40 The amino-terminal region of BPI is comparable to that of LBP and is richer in positively-charged amino acids. 38 The positive charge at its amino terminal might be the reason for its interaction with LPS. 38 The amino terminal of BPI is important for its bactericidal and anti-inflammatory properties.41,42 The affinity of the amino-terminal domain of BPI to LPS is high compared with that of LBP. The amino terminal domain also plays a role in activating the degranulation of the phagocyte in response to the BPI-coated bacteria. 33 The carboxy terminal domain of LBP helps in delivery of LPS to CD14 and is responsible in boosting LPS-mediated signalling, 33 whereas the carboxy terminal domain of BPI plays an important role in opsonization by mediating the interaction between BPI-opsonized bacteria and the phagocyte. 33 These structural differences make LBP, an agonist and BPI, an antagonist for an LPS-mediated inflammatory response.
Recent studies have shown that BPI acts as an anti-angiogenic factor. 43 In vitro and in vivo studies have shown that BPI at a low dose inhibits growth of the endothelial cells, leads to their detachment from the extracellular matrix and induces apoptosis. 43 As there are no endothelial cells for migration the angiogenesis is inhibited. BPI binds to vascular endothelial growth factor (VEGF), through its amino terminal domain, on retinal endothelial cells thereby inhibiting its angiogenic effect. 44 Thus, BPI might be a potential candidate for treating various pathological disorders, such as cancer and atherosclerosis, 43 where angiogenesis play an important role.45,46 The anti-angiogenic property of BPI is postulated to be due to the chemical similarity of BPI with platelet factor 4. 43 It is postulated that both the amino and carboxy terminal of BPI are important in displaying the angiogenic phenomena. 43 In contrast to its angiogenic effect on retinal endothelial cells BPI exerts a mitogenic effect on retinal pericytes and epithelial cells. 44 The mitogenic effect was attributed to the ability of BPI to signal via the ERK and Akt pathways, increasing the synthesis specifically in retinal pigment epithelial cells and pericytes, and not in retinal endothelial cells. 44 This was the first report demonstrating the signalling ability of BPI. Later it was shown that BPI potentially binds to glypican4 (GPC4), a cell surface heparan sulphate proteoglycan exerting mitogenic effects. 47 However, the exact mechanism of ERK and Akt activation via GPC4 is not understood. It was proposed that the binding of BPI to GPC4 might alter its surface structures activating the ERK or PI3K/Akt pathways and modulating VEGF (a potent endothelial cell mitogen and angiogenic factor) or platelet-derived growth factor B (a decrease in which induces/enhances angiogenesis). 47
Regulation of BPI
Regulation of bactericidal/permeability increasing protein in various cell types.

While BPI is a major constituent of neutrophils, regulation of its expression is distinct from other proteins of azurophilic granules. De novo synthesis of BPI increased upon induction of human promyelocytic cell line NB4 with all-trans retinoic acid (ATRA) via binding of transcription factors CCAAT/enhancer binding protein C/EBPε and C/EBPβ to the BPI promoter, thus distinguishing it from other azurophilic granule proteins, the expression of which is regulated by the binding of C/EBPα. 56 Another report of BPI regulation in myeloid cells by Gullberg et al. showed that transcription factors acute myeloid leukaemia protein 1 (AML-1), PU.1 and Sp3 bind directly to the BPI promoter and regulate its expression. 57 While AML-1 has been known to be involved in the regulation of some azurophilic granule proteins, such as myeloperoxidase and elastase, PU.1 is a transcription factor that regulates the expression of several azurophilic and specific granule proteins. 58 BPI mRNA expression in mouse granulocytes was induced by stimulation with TLR ligands. 35 Among the TLR ligands, LPS was the most potent inducer of BPI. It induced the expression of BPI via a MyD88-independent, but TRIF/TICAM-dependent, pathway. 35
Basal expression of BPI is regulated by the binding of transcription factors C/EPBβ, a neutrophil transcription factor and Sp1/3 (ubiquitously expressed in mammalian cells) 58 to the BPI promoter in Caco-2 cells (epithelial colorectal cancer cell line). 59 Aspirin-triggered lipoxin (ATLa), an analogue of anti-inflammatory eicosanoid lipoxin A4 (LXA4) upregulated BPI expression in epithelial (Caco-2) cells. 51
Microarray studies in antrum and gastric mucosal samples of Helicobacter pylori-infected individuals have revealed that the BPI gene is upregulated (13.9-fold induction) in the antrum and corpus gastric mucosa, along with LBP, TLR 1,4,5 and 6, and IL genes IL-1, IL-2R-β, TNF-β, IL-10R-α, IL-16, IFN-γR1, GM-CSFR-β and IL-2R-γ. 60 Recently, increases in BPI expression levels upon infection have been reported in other vertebrates, as well as invertebrates. Caipang et al. showed that BPI–LBP expression in gill epithelial cells of Atlantic cod, Gadus morhua, increased significantly upon incubation with Vibrio anguillarum and atypical Aeromonas salmonicida. 61 The Cg-bpi gene was found to be induced after bacterial challenge in oyster (Crassostrea gigas) haemocytes, although it was constitutively expressed in epithelia, as demonstrated by in situ hybridization. 62 p15A, p15B and proCAP18 belonging to the cathelicidin family of innate host immune polypeptides isolated from rabbits synergize with BPI to bring about increased antibacterial activity against Escherichia coli. 63
BPI regulation by cytokines appears to have a cell type-specific response. Levels of BPI protein, which is constitutive in skin fibroblasts, increases upon stimulation with IL-4, but no increase was detected after TNF-α and IL-1β, or direct exposure to P. aeruginosa, Staphylococcus aureus and Candida albicans. 54 Nevertheless, PMN in culture showed increased release of BPI upon exposure to P. aeruginosa. 64 However, this increase was due to the release of stored BPI and not due to de novo protein synthesis. 64 Similarly, studies on BPI promoter activity in Caco-2 cells in the presence of IFN-γ, TNF-α and IL-1β showed decreased activity. 59 However, no change in BPI mRNA expression was seen in response to stimulation with IL-1β, TNF-α and IFN-γ in human ecto- and endocervical epithelial cells. 55 These observations show that the regulation of BPI is cell type-dependent.
Role of BPI in disease
Diseases associated with BPI can be characterized as either associated with mutations in BPI or due to presence of antineutrophil cytoplasmic Ab (ANCA) against BPI. ANCA are auto-Abs generated against the cytoplasmic Ags (proteins) of neutrophils. Zhao et al. first reported the presence of ANCA-BPI in the serum of patients with vasculitis. 65 BPI is a target of ANCA in a variety of diseases, such as cystic fibrosis, inflammatory bowel disease (IBD),66,67 reactive arthritis, 68 chronic obstructive pulmonary disease (COPD) 69 and transporter associated with antigen processing (TAP) deficiency. 70 The exact way in which anti-BPI Ab is generated is not understood clearly. Anti-BPI Ab can be produced in response to BPI, which may be induced on infection, or to certain epitopes on the pathogens (P. aeruginosa), which can act as molecular mimics of BPI. 71
Cystic fibrosis
Cystic fibrosis (CF) is a hereditary disease characterized by chronic pulmonary inflammation. Even though pulmonary inflammation is associated with infiltration of neutrophils, bacteria, such as P. aeruginosa, are able to colonize the pulmonary epithelium. 72 This paradoxical situation—persistence of P. aeruginosa at the inflammatory site— can be explained by the presence of ANCA against BPI in CF patients. Zhao et al. have shown BPI to be one of the major ANCA antigens in CF patients. 72 The presence of both IgA and IgG anti-BPI auto-Abs have been correlated to the severity of lung disease associated with CF. 71 Both the isotypes against BPI were directed toward its carboxy terminal domain, which is important for opsonization. 71 This might be the reason for the inhibition of BPI-mediated phagocytosis by polymorphonuclear cells by these Abs. 71 This explains the persistence of P. aeruginosa in CF patients. P. aeruginosa isolated from BPI–ANCA-positive and negative CF patients also differ in their properties, such as the ability to produce phycocyanin, the induction of an inflammatory response and apoptosis of neutrophils. 73 Paradoxically, P. aeruginosa isolated from BPI–ANCA-positive CF patients were defective in phycocyanin production and the induction of an inflammatory response. 73 P. aeruginosa isolated from BPI–ANCA-positive CF patients induces less apoptosis of PMN and epithelial cells, giving a further indication that these strains are adapted to the host environment by causing a lesser inflammatory response and slow colonization, which develops, ultimately, into persistent infection. 73
IBD
IBD is characterized by inflammation of the colon and small intestine. Two major types of IBD are Crohn's disease (CD)and ulcerative colitis (UC). The relationship between high BPI levels and IBD was first established by Manojemi et al. 74 BPI was present both extracellularly and intracellularly, and in the crypt lumen. 74 High BPI levels are found in the colonic mucosa of UC patients. 75 One of the first reports concerning the presence of BPI-ANCA in IBD patients came from a study by Stoffel et al. where they found BPI–ANCA in both ANCA-positive and ANCA-negative sera from the patients. 66 BPI was found to be a minor antigen in CD and UC. 67 They also observed a significant correlation between the presence of anti-BPI Abs and CD/UC. 67 Lys216Glu polymorphism in BPI has been associated with IBD. 76 Though the correlation between IBD and BPI is well established, the relationship between specific Lys216Glu polymorphism in the BPI gene and its involvement in IBD remains controversial. 76 Akin et al. have shown that this polymorphism is significantly high in IBD patients, 76 whereas studies by Klein et al. report that the Glu/Glu genotype frequency was significantly decreased in IBD patients. 77 The role and importance of this polymorphism is not yet well defined.
Transporter associated with Ag processing deficiency
Mutations in TAP cause a primary immune deficiency disorder that leads to recurrent Gram-negative bacterial infections78,79 TAP deficiency leads to decreased MHC Class I expression and recurrent Gram-negative bacterial infections. 79 As MHC Class II plays an important role in clearing Gram-negative bacterial infections, it was difficult to correlate TAP deficiency with recurrent Gram-negative bacterial infections.78,79 The lungs of these patients are heavily colonized with P. aeruginosa,78,79 a case similar to that of CF patients. These patients also show the presence of BPI–ANCA directed against the amino terminal region of BPI that might inhibit the bactericidal activity of BPI. 70
Alterations in BPI have been correlated with the occurrence/severity of various diseases. Pstl (T to C) polymorphism in BPI intron # 5 has been correlated with an increased risk of COPD. The levels of plasma BPI were reported to be low in COPD patients. 69 Lower levels of BPI were also observed in the synovial fluids of patients with psoriatic arthritis. 80 One of the first indications of presence of BPI–ANCA in chronic airway infection came from Kobayashi, who showed that 72.7% of patients who have chronic airway infection are BPI–ANCA positive. 81 Patients suffering from liver cirrhosis also have high levels of BPI mRNA in neutrophils and protein in the plasma. 82
BPI in therapeutics
The ability of BPI to bind and neutralize endotoxin has been exploited in therapeutics for the treatment of endotoxin-related problems. A recombinant amino terminal fragment of BPI (rBPI23) was used in a human trial to check for its efficacy in neutralizing endotoxin and its effects. 83 rBPI23 was found to significantly reduce the circulating endotoxin, release of TNF-α, p55 and p75 (soluble TNF-α receptors), Il-6, IL-8 and IL-10. 83 It also prevented endotoxin-induced leucopenia, 83 neutrophil degranulation, 83 fibrinolytic response 84 and activation of coagulation systems. 84 Another, recombinant, amino terminal fragment of BPI (rBPI21) was used as an adjunctive for the treatment of children with severe meningococcal sepsis. 85 However, it did not significantly improve the mortality rate, but had a substantial effect on morbidity. 85 There was a significant decrease in the frequency of amputation in rBPI21-treated patients, probably as a result of the decrease in stimulation of disseminated intravascular coagulation, and propagation of microvascular and macrovascular thrombosis caused by LPS in circulation. 85 rBPI21 had also been used to reduce postoperative complications, and the altered amino-acid ratio and fibrinolytic cascades associated with hemihepatectomy. 86 Although these trials have declared the use of BPI as safe and successful, the administration of large doses of recombinant BPI and the difficulty in its production by conventional methods makes it an expensive therapeutic. Alexander et al. have devised a cost-effective strategy based on an adenoviral vector for the delivery of BPI at the tissue site of interest. 87 This leads to transient, but prolonged, supply of BPI protein into the recipient’s body. 87 Unlike previous studies involving the use of an amino terminal fragment, a full-length BPI gene was cloned, leading to the preservation of all biological activities of BPI. 87 Yet another strategy was designed by Li et al. where they make use of the chimeric protein, which consists of a functional amino terminal domain of human BPI and the Fcγ segment of human IgG1. 88 A gutless adeno-associated vector (AAV2) was used for the delivery of BPI-Fcγ gene. It was demonstrated that the strategy had potential in relieving symptoms of endotoxemia and endotoxin shock in mice. 88
Future perspectives
Considering its multiple functions, BPI has come into the limelight as a therapeutic, especially against various inflammatory diseases. However, much focus needs to be placed on the cost-effective production of BPI. Recent studies by Alexander et al. have already demonstrated a cost-effective method for the constant supply of BPI. 87 Even though the anti-angiogenic function of BPI is well characterized, further work needs to be carried out in order to use BPI as a therapeutic agent for targeting cancerous cells. In patients with airway infection where colonization of P. aeuroginosa, as well as persistence of ANCA against the carboxy terminal of BPI, is known72,73 rBPI may be used for the clearance of bacterial colonization, as well as for preventing inflammation. New roles for this molecule, the induction of differential apoptosis and proliferation of certain cells,44,47 and the presence of ANCA against BPI in TAP-deficient patients 70 open up new avenues for using BPI as a therapeutic agent.
Footnotes
Funding
Arjun Balakrishnan is a Senior research fellow of Council for Scientific and Industrial Research (CSIR) and currently receiving a senior research fellowship as a scholarship.