
Abstract
Zinc signals, i.e. changes in the free intracellular Zn2+concentration, are an integral component of signal transduction in several immune cells. The aim of the present study was to investigate if this is also the case in neutrophil granulocytes. One neutrophil function is NETosis, the release of a matrix composed of DNA, chromatin and granule proteins to capture extracellular bacteria within so-called neutrophil extracellular traps (NET). NETosis can be induced by the protein kinase C (PKC) activator 12-myristate 13-acetate (PMA). PMA treatment led to a zinc signal in neutrophil granulocytes. NETosis was inhibited when the zinc signal was sequestered by the membrane permeable high affinity chelator N,N,N′,N′,-tetrakis (2-pyridylmethyl)ethylenediamine (TPEN). PKC-mediated NET formation depends on the production of reactive oxygen species (ROS) by NADPH oxidase. Inhibition of NADPH oxidase with diphenyleneiodonium chloride blocked ROS formation and NETosis, as well as the zinc signal. TPEN, however, had no effect on PKC activity and ROS production, indicating that Zn2+ is not required for activation of PKC, but for signals downstream of ROS production. In conclusion, zinc signals are an essential component of the ROS-dependent signal transduction leading to NETosis.
Introduction
Neutrophil granulocytes, also known as polymorphonuclear leukocytes (PMN), are the largest population among the cells of the innate immune system and an essential component of the first line of defense against invading pathogens. Once PMN arrive at the site of infection, they employ different strategies to kill pathogens. 1 Primarily, PMN engulf microorganisms and ingest them by phagocytosis. Upon internalization of pathogens into phagocytic vesicles, the latter are fused with lysosomes containing proteins with antimicrobial activity. These proteins include NADPH oxidase, an enzyme capable of producing high amounts of reactive oxygen species (ROS).
PMN extend their antimicrobial activity even beyond their life by a newly described strategy in which they cast out their DNA, chromatin and granule proteins into the extracellular space, forming a matrix known as neutrophil extracellular traps (NET). 2 These NETs capture and kill bacteria, contributing to clearance of the infection. This specialized form of cell death, so-called NETosis, is NAPDH oxidase-dependent, 3 and differs both biochemically and morphologically from necrosis and apoptosis. 4 Several different triggers for NETosis have been identified, including bacteria, LPS and phorbol 12-myristate 13-acetate (PMA), an activator of protein kinase C (PKC). 3 The latter is a potent activator of NADPH oxidase. This enzyme produces superoxide anions, which can be converted into H2O2. Notably, exogenous H2O2 can promote NET production by PMN isolated from chronic granulomatous disease patients, whose cells lack functional NAPDH oxidase. 4
PMN have a short lifespan, are terminally differentiated and there is considerable variation in their ability to produce NETs between different donors.1,4 Moreover, the isolation of primary PMN is time consuming and might cause pre-activation of the cells, complicating the analysis of NETosis. 4 Accordingly, the identification of a human cell line with neutrophil characteristics as a model for NET formation would provide a valuable alternative to primary PMN in the investigation of NETosis.
The cell line NB4, established from a patient with acute promyelocytic leukemia, 5 shows bilineage potential. 6 It undergoes differentiation into morphologically mature PMN in response to all-trans retinoic acid (ATRA) and DMSO, or toward monocytes after exposure to 1α,25-dihydroxyvitamine D3 (1,25D3) and PMA.6,7 N’Diaye and colleagues have reported that ATRA-differentiated NB4 cells express functional NAPDH oxidase and exhibit respiratory burst activity in response to PMA, 8 suggesting that they might have the potential for NET formation.
Zinc was identified as an essential trace element in humans in the early 1960s. 9 In addition to several other biological functions it is indispensible for the immune system; zinc deficiency is associated with an increased risk of infections. 10 Among the cells affected by zinc deficiency are PMN. Here, chemotaxis and the production of ROS were reported to be affected. 11 Notably, the impact of zinc on NET formation has not been investigated so far.
The essentiality of zinc is based partially on its role as a structural component of a great number of proteins, including more than 300 enzymes and a multitude of transcription factors. 12 Consequently, zinc homeostasis is tightly regulated by zinc-binding molecules, such as metallothionein, and two dozen zinc transporters, which mediate Zn2+ transfer through the plasma membrane and between intracellular compartments. 13 Additionally, alternations in the intracellular concentration of free Zn2+ occur, so-called zinc signals. These are induced by extracellular stimuli and participate in several signaling pathways in immune cells. 14 One of these stimuli is PMA, which causes zinc signals in monocytes, suggesting that some signaling pathways downstream of PKC could involve Zn2+. 15
The aim of this study was to investigate a potential role of free intracellular Zn2+ in PKC-mediated NET formation. To this end, NB4 cells differentiated along the neutrophilic lineage were established as a cell culture model for NET formation in response to stimulation with PMA. In these cells, PMA-induced Zn2+ signals were observed and were found to be essential, but not sufficient, for NETosis.
Materials and methods
Materials
RPMI-1640 cell culture medium, pencillin, streptomycin,
Isolation of primary human cells
Primary human leukocytes were isolated from heparinized whole blood of healthy, consenting donors (ethical approval for the use of blood from human volunteers had been obtained from the institutional ethics review board of RWTH Aachen University Hospital under protocol number EK 023/05). One part of a 6% hydroxyethyl starch solution was added to two parts of blood and sedimentation for 45–60 min at room temperature (20–22℃) was followed by two washes with PBS and hypotonic lysis of remaining erythrocytes.
Cell culture and differentiation
The human promyelocytic cell line NB4 was cultured at 37℃ in a humidified 5% CO2 atmosphere. Cells were grown in RPMI-1640 containing 10% FCS, 2 mM
Measurement of cell surface markers
NB4 cells were taken up in PBS and incubated with anti-CD66b FITC-conjugated Ab (clone G10F5) and anti-CD14 PE-conjugated Ab (clone MøP9). Mouse IgMκ-FITC and mouse IgG2bκ-PE were used as isotype-matched controls (all Abs were from BD PharMingen, Heidelberg, Germany). Abs and cells were incubated for 30 min at room temperature in the dark, and washed once with PBS. Fluorescence was measured with a FACScan (Becton Dickinson, Heidelberg, Germany).
Fluorescence microscopy
Cells were activated with PMA (50 ng/ml) at 37℃ in culture medium. After 3 h Hoechst 33258 was added to a final concentration of 10 µg/ml, followed by further incubation for 1 h. Subsequently, cells were transferred onto glass slides by cytospin at 300 g for 5 min. Fluorescence was monitored with a Zeiss Axiskop and images were taken at 100-fold magnification using a Nikon Coolpix 4500 digital camera.
Quantification of NET formation
The release of NET was measured by the enhanced fluorescence emission of PI after its interaction with extracellular DNA. 16 To this end, cells were taken up either in PBS or measurement buffer (25 mM HEPES, pH 7.35, 120 mM NaCl, 5.4 mM KCl, 5 mM Glc, 1.3 mM CaCl2, 1 mM MgCl2, 1 mM NaH2PO4) and seeded into 96-well plates. Subsequently, cells were activated with PMA (50 ng/ml) or H2O2 (1 mM) at 37℃ for 4 h, followed by incubation with propidium iodide (PI, 10 µg/ml) for 5 min in the dark. The resulting fluorescence was recorded on a Tecan Ultra 384 fluorescence well plate reader using an excitation wavelength of 360 nm and emission wavelength of 612 nm
Free Zn2+ measurement
Free intracellular Zn2+ was measured as described previously. 15 Briefly, cells were loaded with 1 µM FluoZin-3 acetoxymethyl ester for 30 min at 37℃ and their fluorescence was measured by flow cytometry. Free zinc concentrations were calculated using a dissociation constant for the Zn2+/FluoZin-3 complex of 8.9 nM, 17 determining the minimal and maximal fluorescence by addition of 50 µM TPEN or a combination of Zn2+ (100 µM) and the ionophore pyrithione (50 µM).
Determination of ROS production
Cells were taken up in PBS and incubated with 1 µg/ml DHR 123 (Invitrogen, Karlsruhe, Germany) for 30 min at 37℃. Afterwards, cells were washed once with PBS and the oxidation of DHR 123 was analyzed by flow cytometry.
Immunoblotting
After incubation as indicated in the figure legend, cells were collected by centrifugation and lysed in sample buffer [6.25 mM Tris-HCl, pH 6.8, 2% (w/v) SDS, 10% glycerol, 0.1% (v/v) β-mercaptoethanol, 0.01% (w/v) bromophenol blue, and 1 mM sodium orthovanadate], sonicated for 10 s and boiled for 5 min at 95℃. An equivalent of 1.5 × 105 cells per lane was separated on 10% polyacrylamide gels at 160 V and blotted onto nitrocellulose membrane (BioRad, Munich, Germany). Uniform loading of gels was confirmed by staining with Ponceau S (Sigma-Aldrich). After destaining, membranes were blocked with 5% fat-free dry milk in TBS-T [20 mM Tris-HCl, pH 7.6, 136 mM NaCl, 0.1% (v/v) Tween 20] for at least 1 h, followed by incubation at 4℃ with a primary Ab against phospho-(Ser) PKC substrate or β-actin, gently shaking overnight (18–20 h; both Abs and biotinylated protein ladder were obtained from Cell Signaling Technology, Frankfurt, Germany). Subsequently, membranes were washed three times with TBS-T and incubated at room temperature for at least 1 h with HRP-linked anti-rabbit IgG secondary Ab and HRP-coupled anti-biotin Ab, followed by detection with LumiGlo reagent (Cell Signaling Technology) on a LAS-3000 (Fujifilm Lifescience, Düsseldorf, Germany).
Statistical analysis
Statistical significance of experimental results was calculated by one-way ANOVA followed by Tukey’s post hoc test using GraphPad prism software. All experiments were performed independently at least three times.
Results
NB4 cells as a model system for NET release
The promyelocytic cell line NB4 is devoid of the characteristic surface markers for monocytes (CD14) and granulocytes (CD66b) respectively (Figure 1A). However, it has the potential to be differentiated along the monocytic or neutrophilic lineage. Incubation of NB4 cells with a combination of 1,25D3 and PMA resulted in differentiation into a monocyte-like phenotype (NB4MONO), which is characterized by augmented expression of CD14 (Figure 1B). In contrast, treatment of NB4 cells with ATRA and DMSO induced the expression of CD66b, confirming a differentiation toward PMN (NB4PMN) (Figure 1C). As shown in Figure 1D, both treatments resulted in a specific differentiation of a significant fraction of cells into the respective phenotypes.
Differentiation of NB4 cells. The expression of characteristic surface markers for monocytes (CD14) or granulocytes (CD66b) was analyzed by flow cytometry. Data are shown as representative histograms comparing Ab-labeled cells (black lines) with isotype-matched controls (filled gray) for NB4 (A), NB4MONO (B), NB4PMN (C) cells and (D) as means of the percentage of CD14 or C66b positive cells + SEM from n = 3 independent experiments.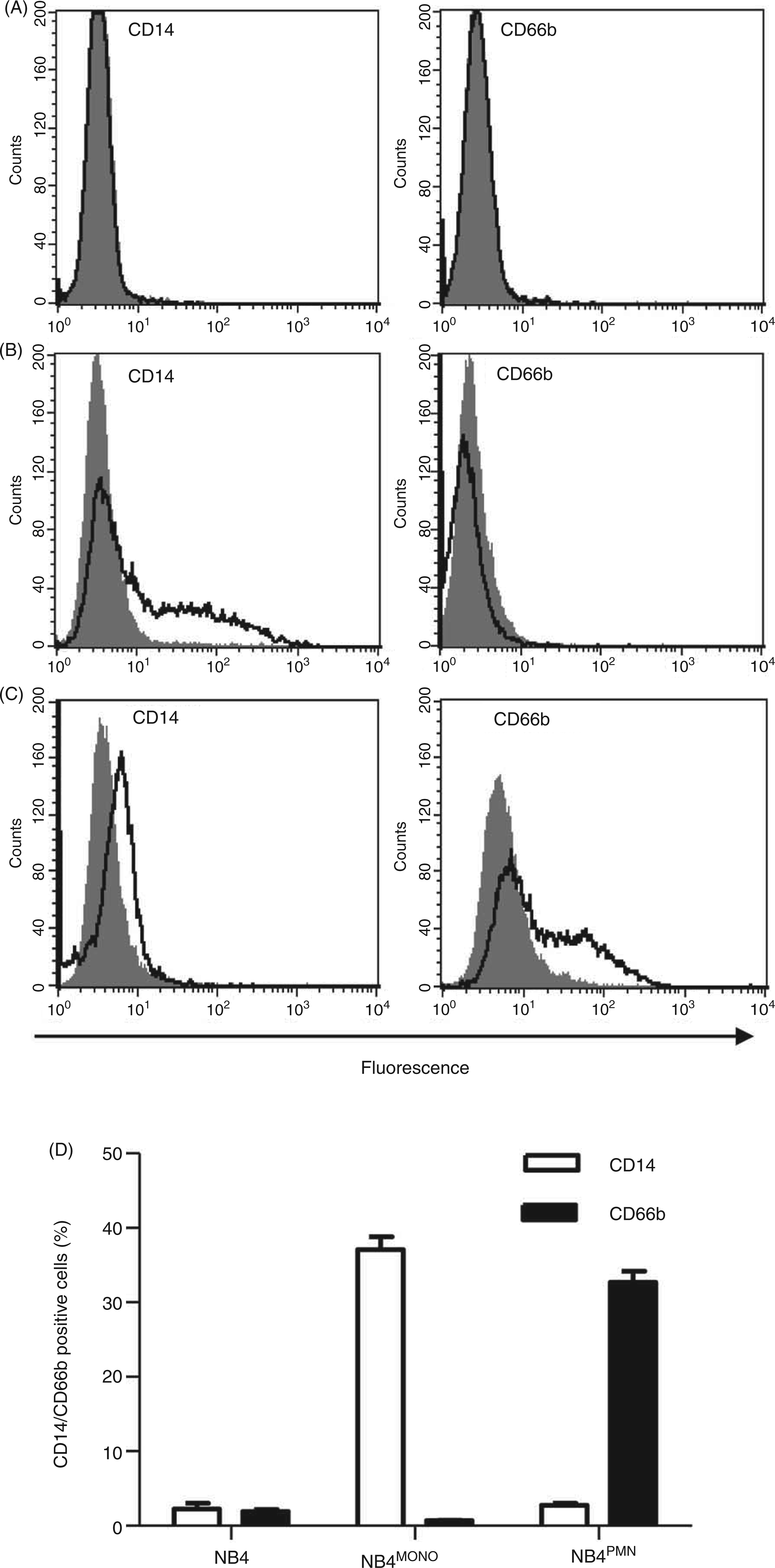
Stimulation with PMA induces NET formation by primary PMN.
3
When PMA was applied to NB4, DNA staining with the membrane-permeable dye Hoechst 33258 in undifferentiated NB4 and in NB4MONO cells showed only intact nuclei, with no difference between untreated controls and cells after PMA stimulation (Figure 2A,B). In contrast, in NB4PMN cells, treatment with PMA resulted in the formation of NET structures (Figure 2C). Based upon its enhanced fluorescence emission after its interaction with extracellular DNA, propidium iodide (PI) can be used to quantify NET formation.
16
As shown in the right panel of Figure 2, NET formation in response to PMA was only observed in NB4PMN cells (Figure 2C), but not NB4 or NB4MONO cells (Figure 2A,B). The formation of NET in NB4PMN cells was completely reversible by the NAPDH oxidase inhibitor diphenyleneiodonium chloride (DPI), confirming an active, ROS-dependent process.
NET formation in NB4 cells. The ability to form NET was compared in undifferentiated NB4 (A), NB4MONO (B) and NB4PMN (C) cells. Fluorescence microscopic images of cells stained with the membrane-permeable DNA dye Hoechst 33258 were taken from control cells (left panel) and cells treated with PMA (50 ng/ml for 4 h, middle panel). Images are representative for n = 4 independent experiments (scale bars = 25µm). In the right panel, extracellular NET DNA was quantified with the membrane-impermeable fluorescent DNA dye propidium iodide (PI). Cells were stimulated as indicated with PMA (50 ng/ml for 4 h) in the presence or absence of DPI (10 µM, added 20 min prior to PMA). Data are shown as means + SEM of n = 3 independent experiments. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other.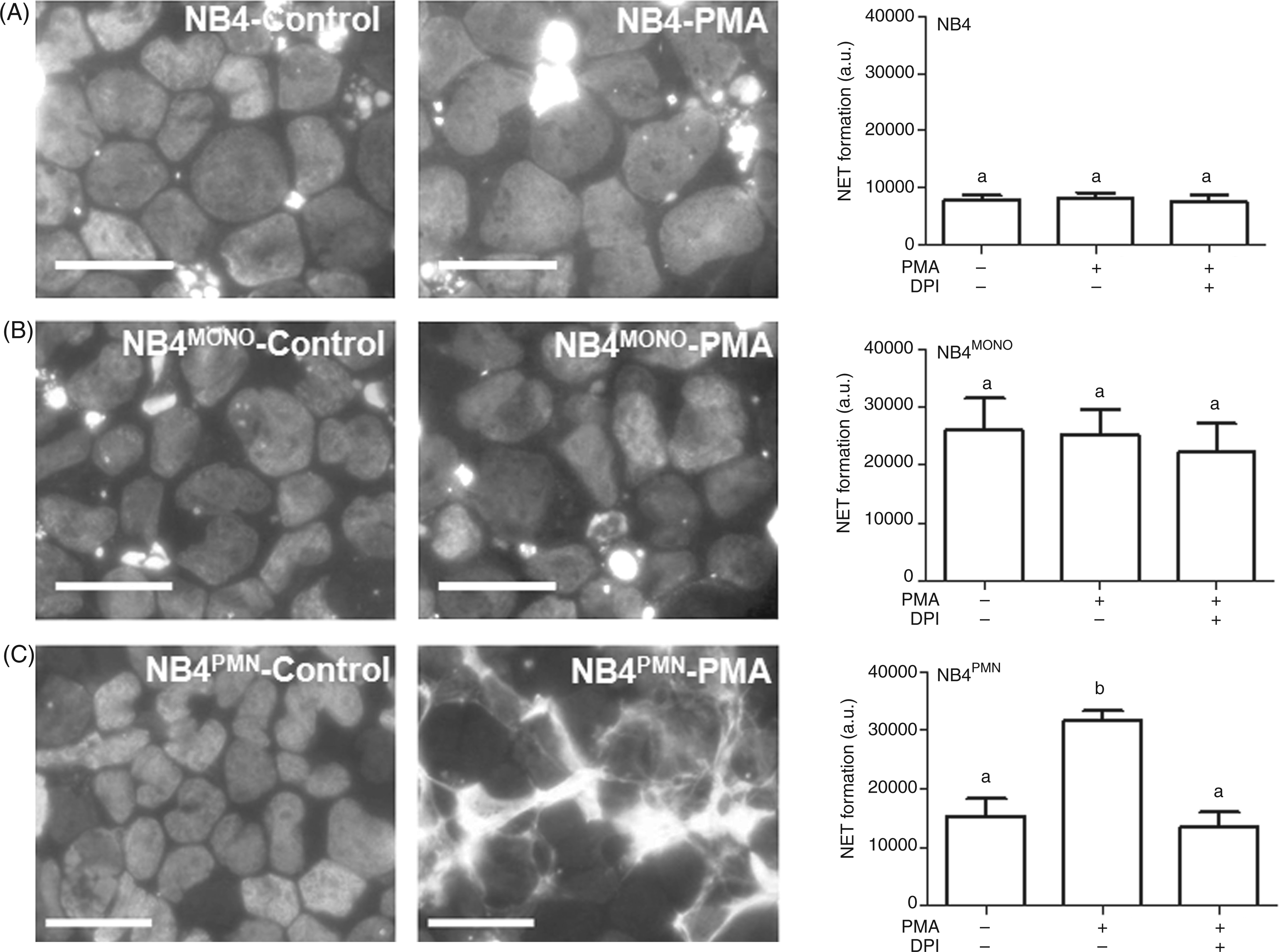
PMA-induced Zn2+ release
PMA increases the intracellular concentration of free Zn2+ in primary human monocytes.
15
To investigate whether the level of intracellular free Zn2+ in granulocytes also changes in response to stimulation with PMA, the zinc-sensitive fluorescent probe FluoZin-3 was used to quantify free Zn2+ in these cells. After loading with FluoZin-3, NB4PMN cells were stimulated with PMA and the corresponding fluorescence was detected by flow cytometry. NB4PMN cells showed a concentration-dependent increase of free Zn2+ upon stimulation with PMA (Figure 3A, Supplementary Figure 1A). Similar experiments were done with isolated leukocytes that were gated into different subpopulations (lymphocytes, monocytes and granulocytes) based on the forward/sideward scatter density plot (Supplementary Figure 1B). A rise in intracellular free Zn2+ in primary granulocytes and monocytes was observed in response to treatment with PMA, whereas no increase was detected in lymphocytes (Figure 3B, Supplementary Figure 1C).
Measurement of intracellular free Zn2+ after activation with PMA. The intracellular free Zn2+ concentration was determined by flow cytometry with FluoZin-3 as described under ‘Materials and methods’. Cells were treated with the indicated concentrations of PMA for 30 min. Results are shown for NB4PMN (A) and primary human leukocytes that were distinguished into granulocyte, monocyte and lymphocyte subpopulations by forward and side scatter (B). Data are shown as means + SEM of n = 6 independent experiments (A) or n = 6 different donors (B) respectively. Statistically significant differences compared with corresponding untreated controls are indicated (*; P < 0.05; ANOVA).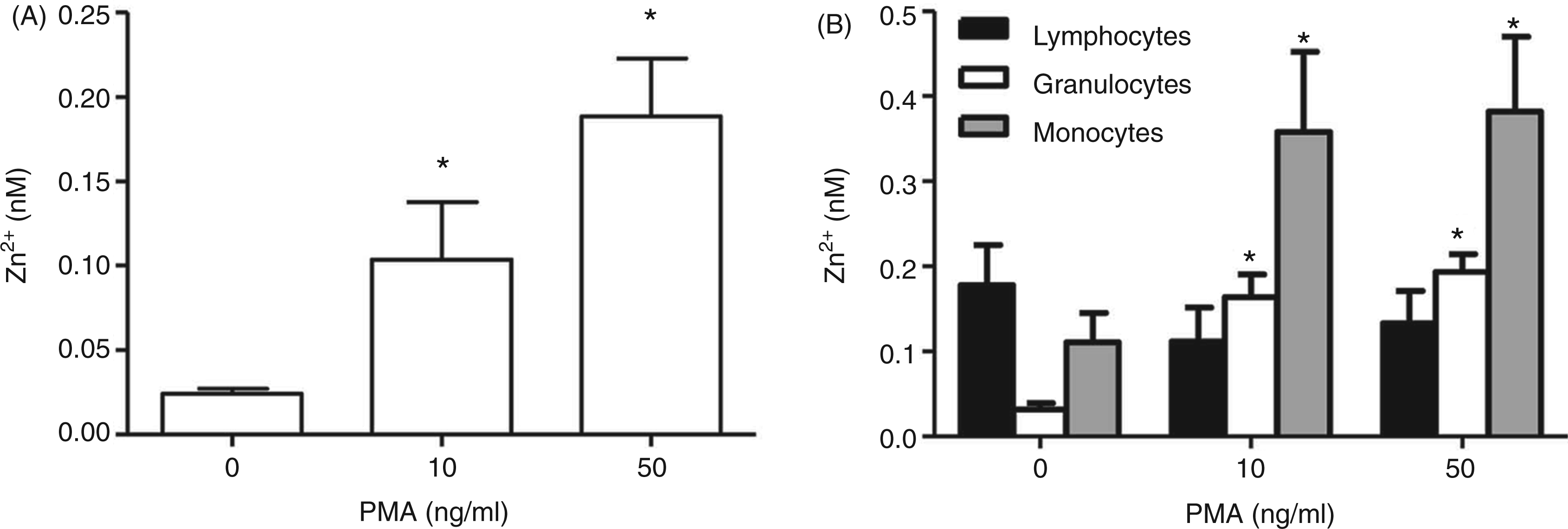
The production of ROS in response to PMA was clearly detectable with the oxidation-sensitive pro-fluorophore DHR123 in NB4PMN cells (Figure 4A, Supplementary Figure 2A) and primary human PMN (Figure 4B, Supplementary Figure 2B). This increase of ROS was completely blocked in the presence of DPI. The Zn2+ signal in both cell types was also sensitive to DPI (Figure 4C,D, Supplementary Figure 2C,D). Taken together, these data indicate a role for NADPH-mediated ROS in generating the Zn2+ signal in response to stimulation with PMA.
Role of ROS in PMA-induced zinc signals. Cells were incubated as indicated with PMA (50 ng/ml, 30 min) and DPI (10 µM, added 20 min prior to PMA). ROS production was measured by flow cytometry with the oxidation-sensitive pro-fluorophore DHR123 in NB4PMN cells (A) and primary human granulocytes (B). Free Zn2+ was measured by flow cytometry with FluoZin-3-loaded NB4PMN cells (C) and primary human granulocytes (D). All data are shown as means + SEM of at least n = 3 independent experiments. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other.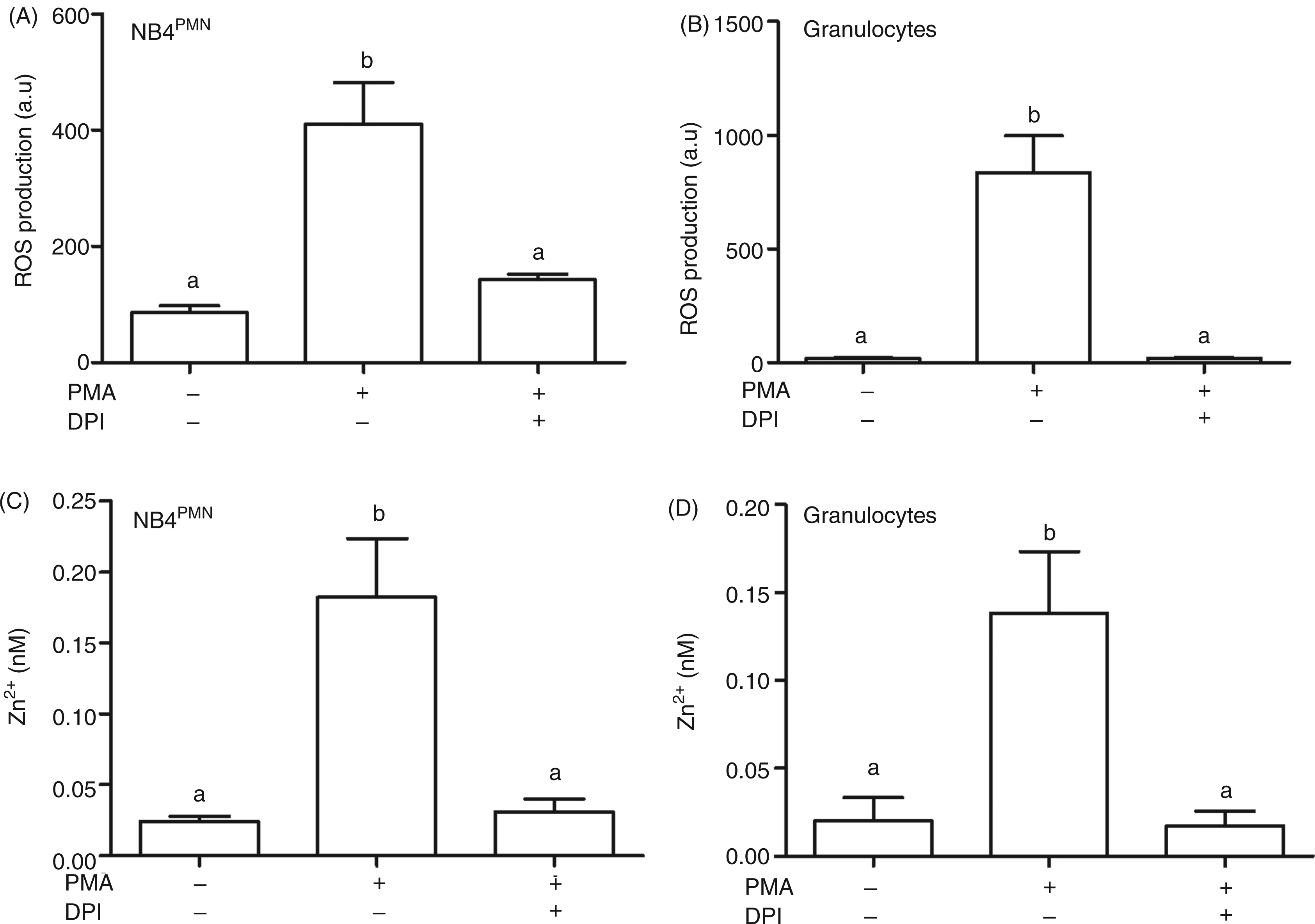
Role of Zn2+ in NET formation
To study the role of the PMA-induced Zn2+ signal in NET formation, Zn2+ ions were sequestered by pre-treatment of the cells with the membrane-permeable Zn2+ selective chelator TPEN. The formation of NETs was reduced by the chelator, both in NB4PMN cells (Figure 5A) and leukocytes (Figure 5B). This concentration of TPEN abolished the PMA-induced Zn2+ increase, but did not completely deplete the cells of free Zn2+ (Figure 5C, Supplementary Figure 3A). At the same time, the production of ROS was unaffected by TPEN (Figure 5D, Supplementary Figure 3B). Moreover, as an indicator of PKC activity, the phosphorylation of typical target sequences for PKC was investigated by a phosphorylation-specific Western blot. It was also unchanged in the presence of the chelator (Figure 5E). These observations indicate that the PMA-induced zinc signal is essential for NET formation, but is not required for PKC activity or the production of ROS.
Role of Zn2+ in PKC-activation. The inhibition by DPI (10 µM) and the concentration-dependent effect of TPEN on NET formation in response to PMA (50 ng/ml, 4 h) was measured by propidium iodide (PI) in NB4PMN cells (A) or primary human leukocytes (B). NB4PMN were incubated with FluoZin-3 (C) or DHR123 (D) in order to measure free Zn2+ or ROS, respectively. This was followed by incubation with the indicated concentrations of TPEN (20 min prior to PMA) and PMA (50 ng/ml, 30 min). All data are presented as means + SEM of at least n = 3 independent experiments (A,C,D) or n = 8 donors (B), respectively. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other. E, NB4PMN cells were treated as indicated with PMA (50 ng/ml for 5 min) and different concentrations of TPEN (added 20 min prior to PMA). To measure PKC activity, Western blot analysis was performed using Abs against phospho-(Ser) PKC substrate (large gel) detecting phosphorylation of PKC target sequences in several proteins. β-Actin (small gel) was used as a housekeeping protein to confirm uniform loading of the gel. One representative Western blot of n = 3 independent experiments is shown.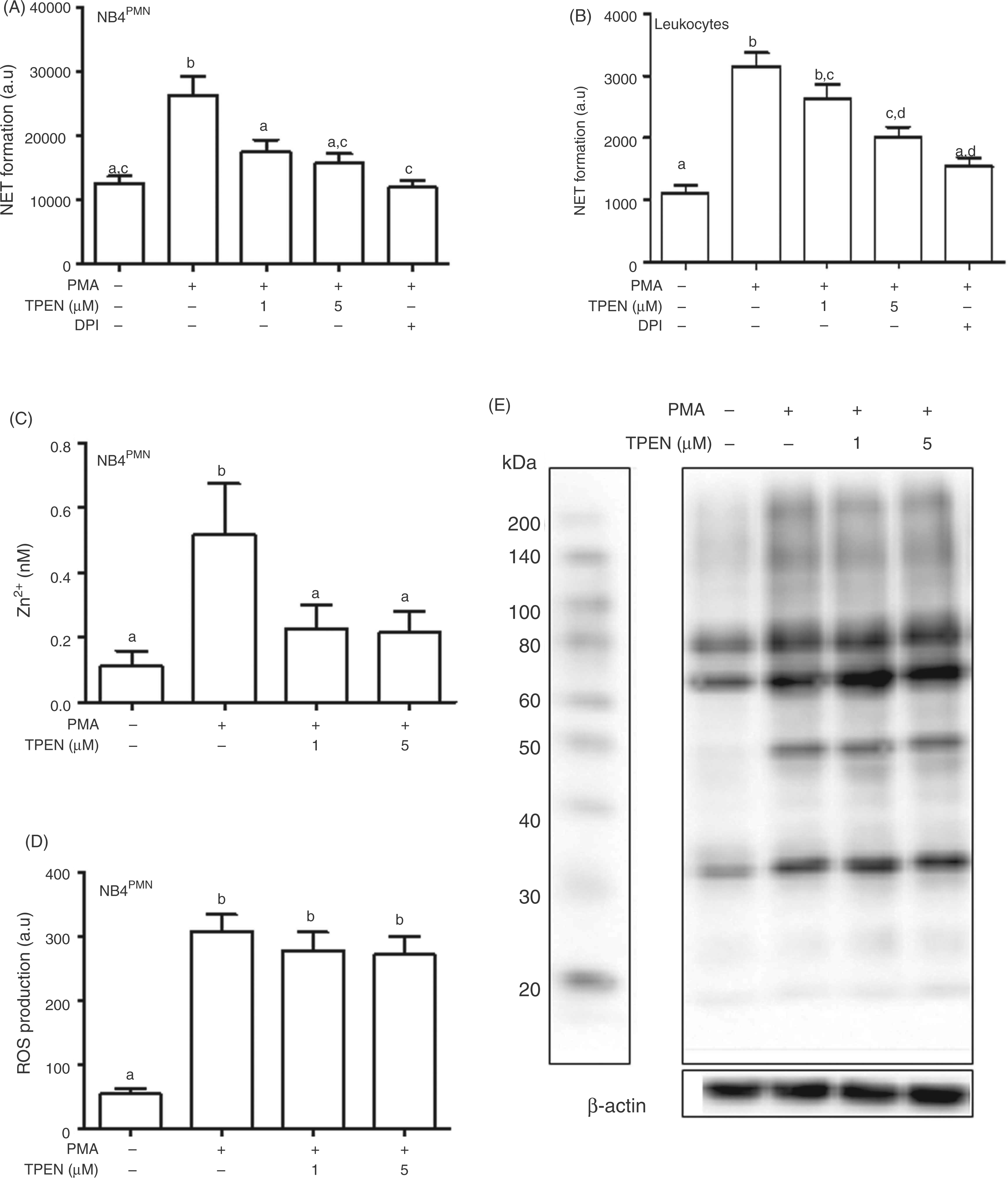
When cells were co-exposed to increasing concentrations of Zn2+ mixed with TPEN (5 µM), addition of equimolar Zn2+ to TPEN almost completely blocked the inhibiting effect of TPEN on NET formation, confirming that the ability to chelate metals is required for its impact on NET (Figure 6A). In further experiments, the effects of TPEN applied together with equimolar amounts of Zn2+, Cu2+, Ca2+ or Mg2+ were compared. NET formation was inhibited by treatment with TPEN alone and was rescued by co-application of equimolar concentrations of Zn2+ or Cu2+, but not Ca2+ or Mg2+ (Figure 6B). The corresponding concentration of free intracellular Zn2+ was measured using FluoZin-3, indicating that TPEN/Ca2+ and TPEN/Mg2+ reduced free Zn2+ in resting cells compared with the chelator alone, whereas TPEN/Zn2+ and TPEN/Cu2+ had almost the same concentration of free Zn2+ as untreated cells in the absence of the chelator (Figure 6C, Supplementary Figure 4).
Effect of co-administration of TPEN with different metal ions on NET formation. (A, B) NB4PMN cells were pre-incubated as indicated with TPEN and different metal ions 20 min before addition of PMA (50 ng/ml, 4 h), followed by quantification of NET formation with propidium iodide (PI). (C) Intracellular free Zn2+ was measured with FluoZin-3 in NB4PMNcells pre-incubated as indicated with TPEN and different metal ions for 30 min. All data are shown as means + SEM from at least n = 3 independent experiments. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other.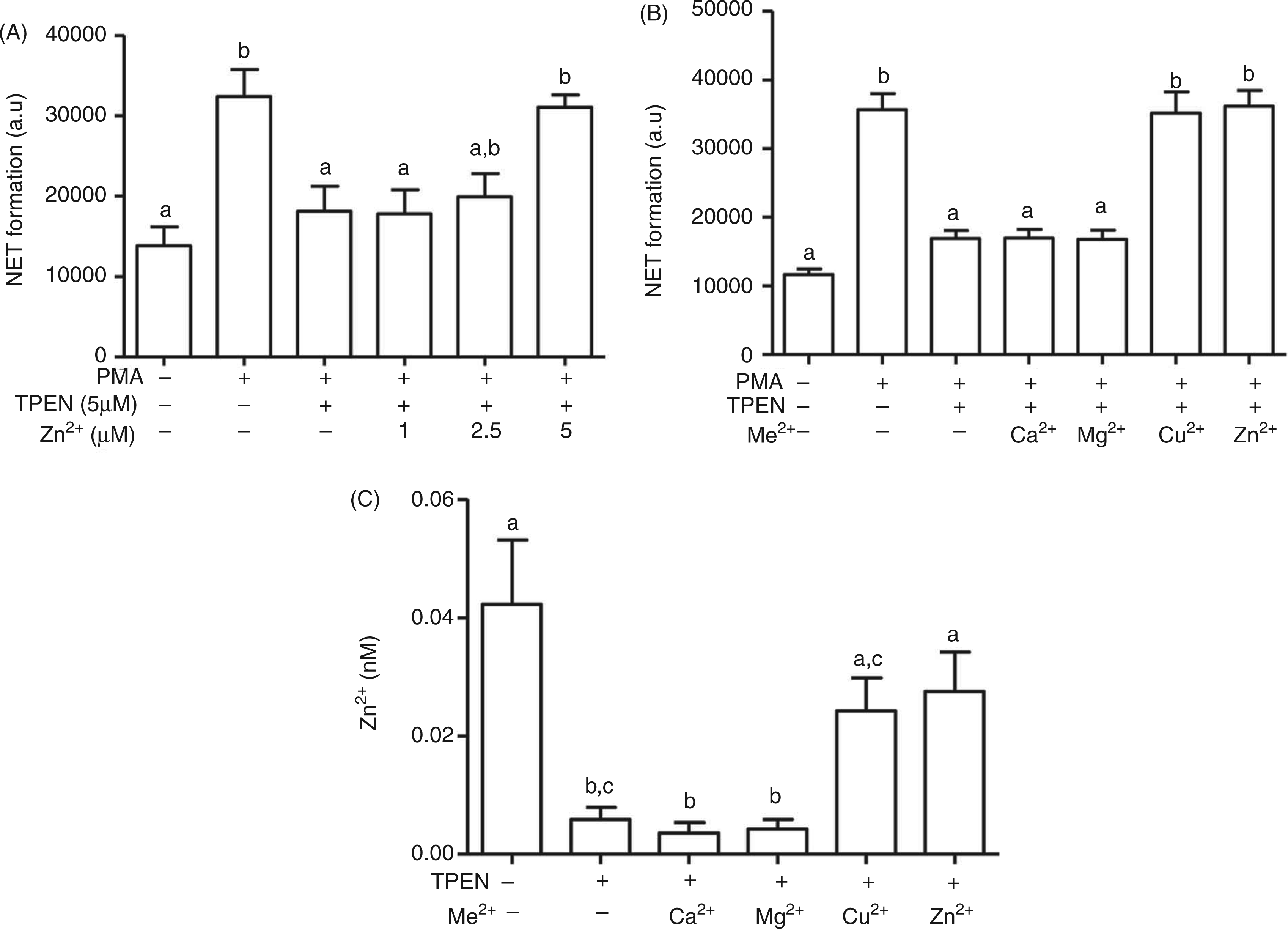
To identify the timepoint during NETosis at which the zinc signal is required, TPEN was added at different times relative to PKC activation, either 20 min prior to, or 30 to 180 min after, PMA. Addition of TPEN up to 30 min after PMA was still sufficient to block NET formation, whereas later administration was ineffective (Figure 7A). Notably, at 60 min no significant NET formation was observed (Figure 7B), demonstrating that the zinc signal precedes NETosis.
Time-dependent effects of PMA and TPEN on NET formation. (A) NB4PMN cells were treated with PMA (50 ng/ml, 4 h), followed by quantification of NET formation with propidium iodide (PI). TPEN (5 µM) was added at the indicated time points relative to PMA, between 20 min before and up to 180 min after addition of the PKC activator. (B) NB4PMN cells were treated with PMA (50 ng/ml) for the times indicated, followed by quantification of NET formation with PI. All data are shown as means + SEM from at least n = 3 independent experiments. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other.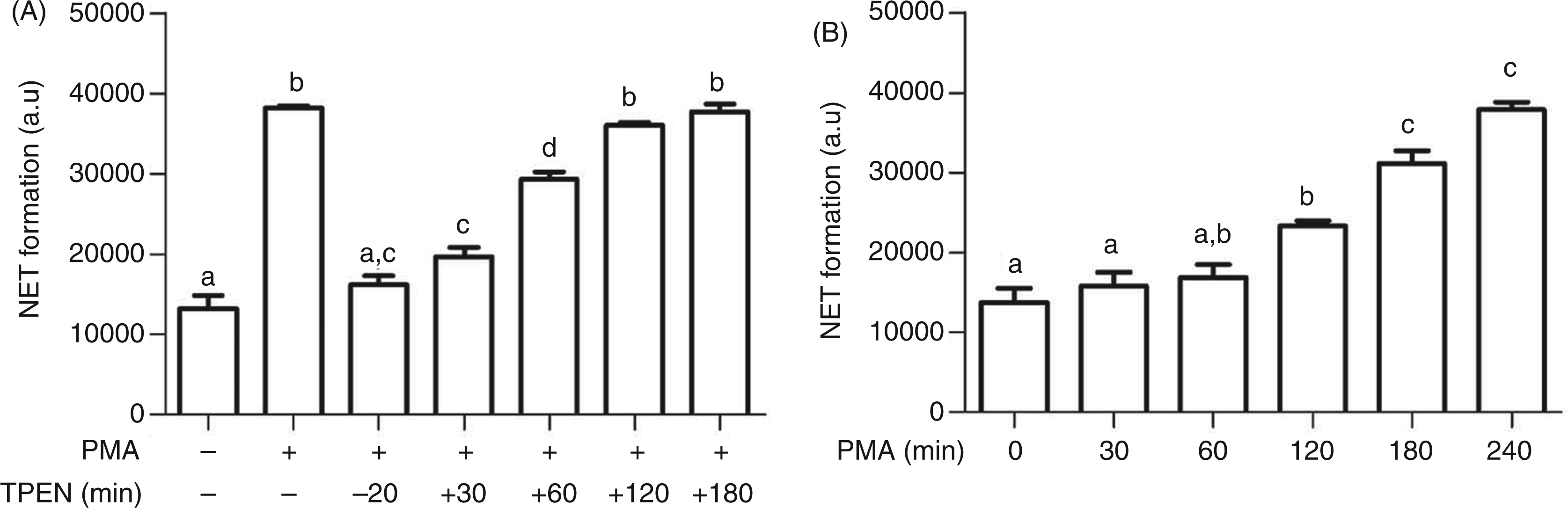
Role of ROS and Zn2+ in NET formation
Mimicking the synthesis of ROS by NADPH oxidase through addition of exogenous H2O2 leads to elevated free Zn2+ (Figure 8A, Supplementary Fig. 5A). This further supports an oxidative mechanism for generation of the zinc signal. In addition, treatment with H2O2 resulted in the formation of NETs, indicating that generation of ROS is sufficient for triggering all signaling pathways necessary for NET formation (Figure 8B). Chelation of the zinc signal by TPEN inhibited the H2O2-induced NET release, confirming that Zn2+ is essential for NET formation in response to H2O2, as well as PMA (Figure 8B).
Role of ROS in Zn2+ signals and NET formation. (A) NB4PMN cells were loaded with FluoZin-3 followed by treatment with H2O2 (1 mM, 30 min) and measurement of the Zn2+-dependent fluorescence by flow cytometry. (B) NB4PMN cells were pre-incubated with TPEN (5 µM, 20 min) prior to stimulation with PMA (50 ng/ml) or H2O2 (1 mM) for 4 h. NET formation was assessed by propidium iodide (PI). (C,D) NB4PMN cells were treated with either PMA (50 ng/ml) or Zn2+ (2.5 µM) and the ionophore pyrithione (1 µM). Intracellular free Zn2+ was determined with FluoZin-3 after 30 min (C) and NET formation by staining with PI after 4 h (D). All data are shown as means + SEM of at least n = 3 independent experiments. Bars that do not share the same letters are significantly different (P < 0.05; ANOVA) from each other.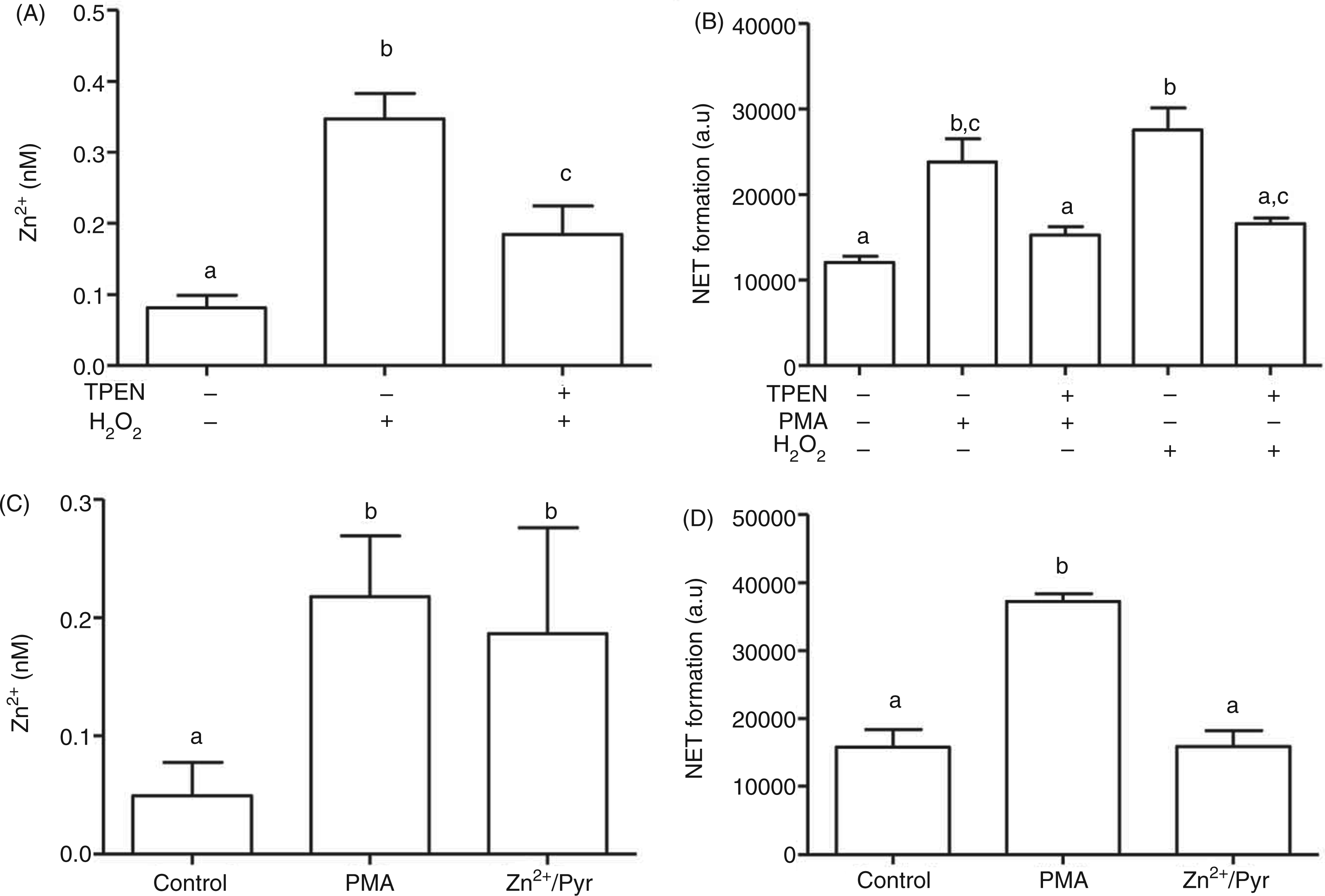
Addition of Zn2+ (2.5 µM) in the presence of the ionophore pyrithione (1 µM) led to a comparable increase of intracellular free Zn2+ than PMA treatment (Figure 8C, Supplementary Figure 5B). However, this zinc signal alone did not result in NET formation (Figure 8D), which may be based on a different time course of this artificial zinc signal compared with the one triggered by ROS. Alternatively, this observation could suggest that the zinc signal, even though required, is not sufficient for triggering NET in the absence of additional ROS-dependent signals, such as the ones initiated by PMA or H2O2.
Discussion
Zinc deficiency negatively affects human health, particularly through its impact on the immune system. Without a sufficient supply with zinc, defense against infectious agents is impossible; infections as a result of zinc deficiency are a major cause of the loss of healthy life years. 18 This is based on two biological roles of zinc: (i) its role as a prosthetic group in proteins, and (ii) a function of free Zn2+ in signal transduction. Several examples for the latter have been reported, including the regulation of the activity of monocytes, dendritic cells, mast cells, and T cells. 14 PMN are also functionally impaired in zinc deficient individuals, but a function of Zn2+ in their signal transduction still remains to be shown.
One particular function of PMN—NETosis—which was discovered a few years ago, is still poorly understood with regard to the intracellular mechanisms by which it is regulated. The most frequently used pathway of induction is triggered by PMA, a PKC activator. PKC, in turn, activates the NADPH oxidase complex. It produces superoxide anions, the starting product of the ROS that constitute the oxidative burst, a mechanism by which PMN kill phagocytosed pathogens. In addition, it was shown that these ROS are also required to trigger the release of NETs, consisting of DNA, chromatin and antibacterial granule proteins, forming a matrix in which extracellular pathogens are captured and killed. 3
NB4 cells are a pro-myeloid cell line that have retained the ability to develop into monocytes or PMN. 6 For neutrophil differentiation, this includes expression of functional NAPDH oxidase, 8 the starting point of the signals that trigger NETs. The data in Figure 1 demonstrate that by differentiating into PMN, NB4PMN cells have gained the ability to undergo PMA-dependent NETosis, which is blocked by inhibition of NADPH oxidase. In contrast, no NETosis is observed in undifferentiated NB4 and NB4MONO cells, demonstrating that NB4PMN cells are a suitable and specific cell culture model for investigation of NETosis.
An increase in the intracellular concentration of free Zn2+ has been observed in T cells stimulated by phorbol ester. 19 PMA has also been reported to cause zinc signals in monocytes, a cell type which develops from the same precursor as neutrophils. 15 A comparable zinc signal was also found in NB4PMN cells and primary human PMN, indicating that Zn2+ could be involved in the signal transduction leading to NETosis.
Inhibition of zinc signals by the NADPH oxidase inhibitor DPI suggests that Zn2+ is released as a consequence of ROS production. This is in contrast to reports in several other signaling contexts, in which zinc is acting further upstream, activating PKC. Activation of this kinase was inhibited by the zinc chelator TPEN,20,21 but the concentrations used for inhibition (up to 100 µM) were significantly higher than the 5 µM or less that were used in the present study. Moreover, addition of exogenous Zn2+ to cell cultures induced PKC translocation to the cytoskeleton, which is part of its activation. 22 As shown in Figure 5, production of ROS was unaffected by the same concentrations of TPEN that inhibited NETosis. Furthermore, PMA-mediated phosphorylation of several proteins containing a PKC target sequence, which was taken as a measure for PKC activity, was also unchanged by TPEN. This shows that the rise in free Zn2+ in response to stimulation with PMA is a consequence of PKC-mediated NADPH oxidase activation, but is not required for activation of PKC. The source of the Zn2+ remains unknown; it has been demonstrated that ROS can release Zn2+ from Zn2+-binding proteins such as metallothionein 23 or PKC itself. 24 This is clearly different from other signaling pathways, such as IL-2-signaling in T cells, during which lysosomal Zn2+ is released.25,26
Investigation of the time course in Figure 7 showed that TPEN blocks NET formation even if added up to 30 min after PMA, which is the timepoint at which the Zn2+ measurements in Figures 3, 4 and 5 showed elevated levels of free intracellular Zn2+. TPEN enters cells within seconds; 15 therefore, it should chelate the zinc signal immediately after its addition. This indicates that free Zn2+ is required during NETosis between 30 and 60 min after PKC activation. These data support a sequence of events in which PKC triggers production ROS, which, subsequently, raise intracellular Zn2+.
An effect of TPEN downstream of its conventional effect on PKC signaling calls for an investigation if it is really acting as a Zn2+ chelator, or, potentially, as a low molecular mass inhibitor independent of its metal chelating ability. The latter effect has already been observed in TPEN-induced apoptosis of PC12 cells.
27
Application of a Zn2+/TPEN complex has no impact on NETosis, showing that TPEN is ineffective when it can no longer bind cellular metal ions because it is already saturated. This was not observed for TPEN complexes of Ca2+ or Mg2+, demonstrating that chelation of these two ions is not the molecular basis for the effect of TPEN. However, addition of Cu2+ also prevented TPEN from inhibiting NETosis, raising the question of whether TPEN acts via chelation of Zn2+ or Cu2+. The affinity of TPEN for Cu2+ is about five orders of magnitude stronger than the one for Zn2+ (
The high affinity of TPEN for Zn2+ raises the possibility that, instead of chelating free Zn2+, the chelator could inhibit NETosis by removal of Zn2+ from a critical protein, which subsequently loses its function. However, after addition of 5 µM TPEN (Figure 5C), there still was some free intracellular Zn2+ left for detection by FluoZin-3, which has a nanomolar dissociation constant for this ion. 17 Thermodynamically, a chelator will prefer the pool of Zn2+ that is bound with the lowest affinity. Hence, TPEN will not remove tightly bound Zn2+ from proteins, unless all free (and FluoZin-3-bound) ions are already chelated. Notably, in the literature more than 100 µM of TPEN were required to remove protein-bound Zn2+ from a zinc finger structure. 29 Additionally, as shown in Figure 5, no effect of 5 µM TPEN on PKC activation was observed. As mentioned above, higher concentrations of TPEN do inhibit PKC.20,21
Addition of H2O2 to NB4PMN cells confirms that ROS are sufficient for triggering NETosis, as reported by Fuchs et al. 4 As for PMA-stimulated NETosis, a zinc signal is also involved if H2O2 is used for inducing NETs. Chelation of Zn2+ abrogates NETosis, demonstrating that zinc signals are essential for the signal transduction leading to NET formation in response to H2O2, as well as PMA. However, when an intracellular zinc signal was generated by application of extracellular Zn2+ in the presence of the ionophore pyrithione, no NETosis was observed. Hence, simply raising the intracellular Zn2+ concentration is not sufficient to trigger NETs in the absence of ROS. The time course after treatment with Zn2+ and pyrithione may be different from the physiological course of the zinc signal and therefore unsuitable to cause NETosis. Alternatively, it is very likely that Zn2+ is only one second messenger acting in concert with other ROS-derived signals.
In conclusion, zinc signals are utilized by PMN. In the present example, PKC activates NADPH oxidase, which synthesizes ROS that release Zn2+. This Zn2+ is then one of several ROS-derived signals leading to NETosis. It remains to be identified which molecular targets interact with Zn2+ in the downstream signal transduction. So far, only some parts of the mosaic of signaling pathways involved in NETosis have been identified. Many different signal transduction pathways in immune cells are known to involve Zn2+, 14 and it will have to be determined which of them are involved in the formation of NETs and which one(s) are actually responsible for the observed zinc-dependence.
Footnotes
Funding
This study was supported in part by German Research Council (DFG) grant HA4318/3 to H. H. Furthermore, R. H. received a scholarship from the German Academic Exchange Service (DAAD).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.