
Abstract
LPS-binding protein (LBP) functions as a crucial molecule in innate immune responses to bacterial challenge. Our study has shown the expression of LBP in human gingiva and its significant association with periodontal health and disease. Porphyromonas gingivalis is a key pathogen of periodontal disease. P. gingivalis LPS as a main virulence factor is strongly involved in periodontal pathogenesis and it displays a significant lipid A structural heterogeneity. Currently, it remains unknown whether, and to what extent, the lipid A structural heterogeneity of P. gingivalis LPS affects LBP expression. The present study investigated the expression profile of LBP in human oral keratinocytes (HOKs) stimulated by two isoforms of P. gingivalis LPS [tetra- (LPS1435/1449) and penta-acylated (LPS1690)] and Escherichia coli LPS, and the involvement of TLRs in LBP expression. The results showed that the expression of LBP mRNA and peptide was significantly up-regulated by P. gingivalis LPS1690 and E. coli LPS, while P. gingivalis LPS1435/1449 did not affect LBP expression. Blocking assay and siRNA gene silencing revealed that P. gingivalis LPS1690-induced LBP expression was through both TLR2 and TLR4. This in vitro study demonstrates that P. gingivalis LPS with a lipid A structural heterogeneity differentially modulates LBP expression in HOKs.
Introduction
The innate immune system is the first line of host defence and provides an immediate response against infection. LPS-binding protein (LBP) as an acute-phase protein is a major participant in innate immune responses to LPS. 1 It is a 50-ku polypeptide mainly synthesized in hepatocytes. It is released as a 58–60-ku glycoprotein into the bloodstream after glycosylation.1,2 LBP is present in the blood at a concentration of approximately 10 µg/ml in healthy individuals, 3 and its plasma level rises dramatically during acute phase responses, such as Gram-negative bacteria-induced sepsis. 4 LBP functions catalytically as a lipid transfer protein by presenting LPS to the pattern recognition receptors on cell surface, such as mCD14 and TLR4, to initiate a host immune response. Many observations suggest that LBP may play a crucial role in the clearance and killing of Gram-negative bacteria within injured and periodontally inflamed tissues.5,6 Over the years, serum levels of LBP have been widely studied as a predictive biomarker in certain endotoxin-related diseases, such as sepsis, infectious endocarditis and Crohn’s disease.7–10 Emerging evidence shows that oral infections, such as periodontal disease, are significantly linked to systemic diseases or disorders through contributing to the systemic level of inflammation with the measurement of certain serum biomarkers, such as C-reactive protein (CRP) and IL-6.11–13 Our recent in vivo study shows that LBP can be expressed by gingival epithelial cells and its expression level in periodontal healthy subjects is significantly higher than that in patients with chronic periodontitis,14,15 suggesting that LBP may be not only significantly involved in innate response to LPS in human gingiva—thereby contributing to periodontal pathogenesis—but may also be potentially linked to systemic inflammation.
Porphyromonas gingivalis is a key periodontopathogen owing to its strong association with periodontitis.16,17 P. gingivalis LPS is one of the most crucial virulence factors and it is significantly involved in periodontal pathogenesis.6,18 It differs from Escherichia coli LPS in structure and various functional activities.18–20 The exact nature and mechanism of P. gingivalis and its LPS interactions with host defence remain unclear.16,21 It is currently evident that P. gingivalis LPS is highly heterogeneous and contains both a tetra- (LPS1435/1449) and penta-acylated (LPS1690) lipid A structure. 22 It may utilize its lipid A structural content to modulate the innate host response in different microenvironmental conditions, such as increased hemin concentration in periodontally diseased sites that can change the lipid A moiety. This may account for some of the apparent discrepant results gained on P. gingivalis LPS-mediated host immuno-inflammatory responses.17,21–25
In the human oral cavity, the oral mucosa forms a crucial interface between bacteria/bacterial products, such as endotoxin, and the interior of host. Human oral keratinocytes (HOKs)—as the main structural cells in oral and gingival epithelia—function as the first line of physical and biological barriers in preventing bacterial challenge, thereby maintaining tissue homeostasis. The active and appropriate interaction of HOKs with P. gingivalis and its LPS plays an important role in the maintenance of oral and gingival health. Currently, it remains unknown how the different isoforms of lipid A structure of P. gingivalis LPS (LPS1435/1449 and LPS1690) may affect host defence through modulation of the expression of innate defence proteins, such as LBP. Furthermore, despite a number of studies conducted on the role of LBP in certain inflammatory diseases,1,26 there are currently no studies on how the expression of LBP per se in human gingiva is regulated upon the stimulation of bacteria and/or bacterial endotoxin.
The present study examined, for the first time, the effects of the two isoforms of P. gingivalis LPS1690 and LPS1435/1449 on the expression of LBP in HOKs, and investigated the possible involvement of pattern recognition receptors like TLR2 and TLR4 in P. gingivalis LPS-induced LBP expression.
Materials and methods
Cell culture
The primary HOKs (ScienCell Research Laboratories, Carlsbad, CA, USA) were cultured in oral keratinocyte medium (OKM) at 37°C in a humidified atmosphere of 5% CO2 in air. The OKM is serum free and contains basal medium, 1% oral keratinocyte growth factor supplement, and 1% penicillin and streptomycin solution. Cells from passage three were used for the subsequent experiments.
Preparation of LPS
P. gingivalis (ATCC 33277) LPS was a gift from one of the co-authors (RPD). It was prepared by an established cold MgCl2-ethanol procedure.22,23,27 The LPS was then purified to keep final protein contamination <0.1%. Tetra- (LPS1435/1449)) and penta-acylated (LPS1690) lipid A structures were prepared and analysed using the matrix-assisted laser desorption ionization time-of-flight mass spectrometry. 22 E. coli LPS (0111:B4) (Sigma, St Louis, MO, USA) was used as a reference control. Pam3Cys was obtained from InvivoGen (San Diego, CA, USA).
Stimulation of HOKs by LPS
The cells were seeded in 6-well plates at a density of 5 × 104 cells/well. Cells were treated with various concentrations (10 ng/ml–1 µg/ml) of P. gingivalis LPS1690, P. gingivalis LPS1435/1449 or E. coli LPS for 24 h in dose-dependent assays. Upon the results, 100 ng/ml of P. gingivalis LPS1690 or E. coli LPS was selected as the optimal concentration for the subsequent time-dependent assay, in which HOKs were treated for 6 h, 12 h, 24 h, 48 h and 72 h. HOKs treated with culture medium alone served as the negative controls.
Immunofluorescence staining and confocal microscopy
The cells were seeded on coverslips in six-well plates, fixed, permeabilized and then incubated with goat anti-human LBP polyclonal IgG (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at a 1:200 dilution at 4°C for 18 h. After further incubation with Alexa Fluor 488 donkey anti-rabbit IgG at 1:400 (Invitrogen, Carlsbad, CA, USA) and Alexa Fluor 555 phalloidin at 1:40 (Invitrogen), the samples were examined by a confocal microscopy (Olympus Fluoview FV1000; Olympus, Tokyo, Japan). Cells treated with goat IgG isotype control (Santa Cruz Biotechnology) instead of primary Ab served as the negative control.
Reverse transcription and real-time PCR
Total cellular RNA was extracted using RNeasy Mini Kit (Qiagen, Hilden, Germany). One microgram of total RNA from each sample was reverse transcribed into cDNA in a final volume of 20 μl using the QuantiTect® Reverse Transcription kit (Qiagen). Reverse transcription (RT-PCR) was performed using the StepOne RT-PCR System (Applied Biosystems, Foster City, CA, USA). β-Actin was amplified as an internal control. The specific sequences of primers were synthesized as follows: LBP (NM_004139.2), forward: 5′-CTG AAG CCA GGA AAG GTA AAA G-3′; reverse: 5′-GCA GCC CAA GGT CGT AGA-3′; TLR4 (NM_138554.4), forward: 5′-AATCCCCTGAGGCATTTAGG-3′; reverse: 5′-CACAGCCACCAGCTTCTGTA-3′; TLR2 (NM_003264.3), forward: 5′-CTT TCA CTG CTT TCA ACT GGT AG-3′; reverse: 5′-TTG CGG TCA CAA GAC AGA G-3′; and β-actin (NM_001101.3), forward: 5′-TTG GCA ATG AGC GGT T-3′; reverse: 5′-AGT TGA AGG TAG TTT CGT GGA T-3′. The reaction conditions were set at 95°C for 20 s followed by 40 cycles at 95°C for 3 s and 60°C for 30 s. The results were analysed using the comparative cycle threshold (Ct) method. 28
ELISA
The culture media were collected and the level of IL-6 was determined by ELISA (R&D Systems Inc., Minneapolis, MN, USA) with a minimal detectable concentration of 10 pg/ml. The absorbance values were determined by a microplate reader (Victor, Vienna, VA, USA) at an optical absorbance of 450 nm. The final concentration was determined with reference to a standard curve for IL-6.
Blocking assay against TLR2 and TLR4
For blocking assay, firstly the efficiency of anti-human TLR2 (eBioscience Inc., San Diego, CA, USA) and anti-human TLR4 (eBioscience) mAbs were tested. Pam3Cys (a TLR2 agonist) and E. coli LPS (a TLR4 agonist) were employed as the positive controls for the efficiency of anti-TLR2 and TLR4 mAb respectively. HOKs were pre-incubated for 1 h with anti-TLR2 mAb or anti-TLR4 mAb at 10 µg/ml prior to incubation with 100 ng/ml of E. coli LPS and Pam3Cys for 24 h. The IL-6 level in the culture media was detected by the ELISA. After confirmation of the blocking efficiency, the above two Abs were used prior to the 100 ng/ml of P. gingivalis LPS1690 and E. coli LPS stimulation for 24 h or 48 h. Subsequently, LBP level was detected by RT-PCR and Western blot. The HOKs incubated with medium alone were used as the negative control, and HOKs incubated with LPS and IgG served as the positive control.
Western blot
Total cell lysates were prepared. Cellular extracts were subjected to SDS/PAGE and transferred onto a PVDF membrane. After incubation with anti-human LBP (Santa Cruz Biotechnology) or alpha-tubulin (Cell Signaling Technology, Danvers, MA, USA) Abs (1:2,000) at 4°C for 18 h, secondary Abs (Invitrogen) at 1:20,000 were added and incubated for 1 h. The signal was detected by SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific, Rockford, IL, USA). To quantify the bands obtained via Western blot analysis, ImageJ software-based analysis (http://rsb.info.nih.gov/ij/) was performed to quantify the integrated density.
TLR2 siRNA silencing knock-down assay
Three Silencer® Select siRNAs (Applied Biosystems) targeting human TLR2 (#168, #169 and #170), and one Silencer® Select GAPDH siRNA or Silencer® Select Negative Control#1 siRNA were selected. All transient transfections were carried out using siPORT NeoFX Transfection Agent (Applied Biosystems) according to the manufacturer’s instructions. After 48 h, the efficiency of gene knock-down was evaluated by real-time RT-PCR. The cells were then exposed to P. gingivalis LPS1690 or E. coli LPS for 24 h for detection of LBP transcripts.
Statistical analysis
All the experiments were repeated in three independent assays. The results were presented as mean ± SD. If both the normality test and the test of homogeneity of variances were justified, the statistical significance was determined using one-way ANOVA (Dunnett test or Bonferroni and least significant difference methods). Otherwise, a non-parametric test was used. A P value of <0.05 was considered statistically significant. The analysis was undertaken by IBM SPSS Statistic 19 (SPSS Inc., IBM Corp., Armonk, NY, USA).
Results
Basal expression of LBP in HOKs
The expression of LBP protein was detected basally in the cytoplasm and nucleus of HOKs, and intensive expression was observed around the nucleus (Figure 1). Cells treated with isotype control instead of primary Ab served as the negative control.
(A) LBP peptide is detected basally in the cytoplasm and nucleus of HOKs and intensive expression is observed around the nucleus (white arrow). The cells were fixed and incubated with anti-LBP Ab. After further incubation with Alexa Fluor 488 secondary Ab and Alexa Fluor 555 phalloidin, the samples were examined by confocal microscopy. (B) The negative control. Scale bar = 50 µm. One representative experiment of three is shown.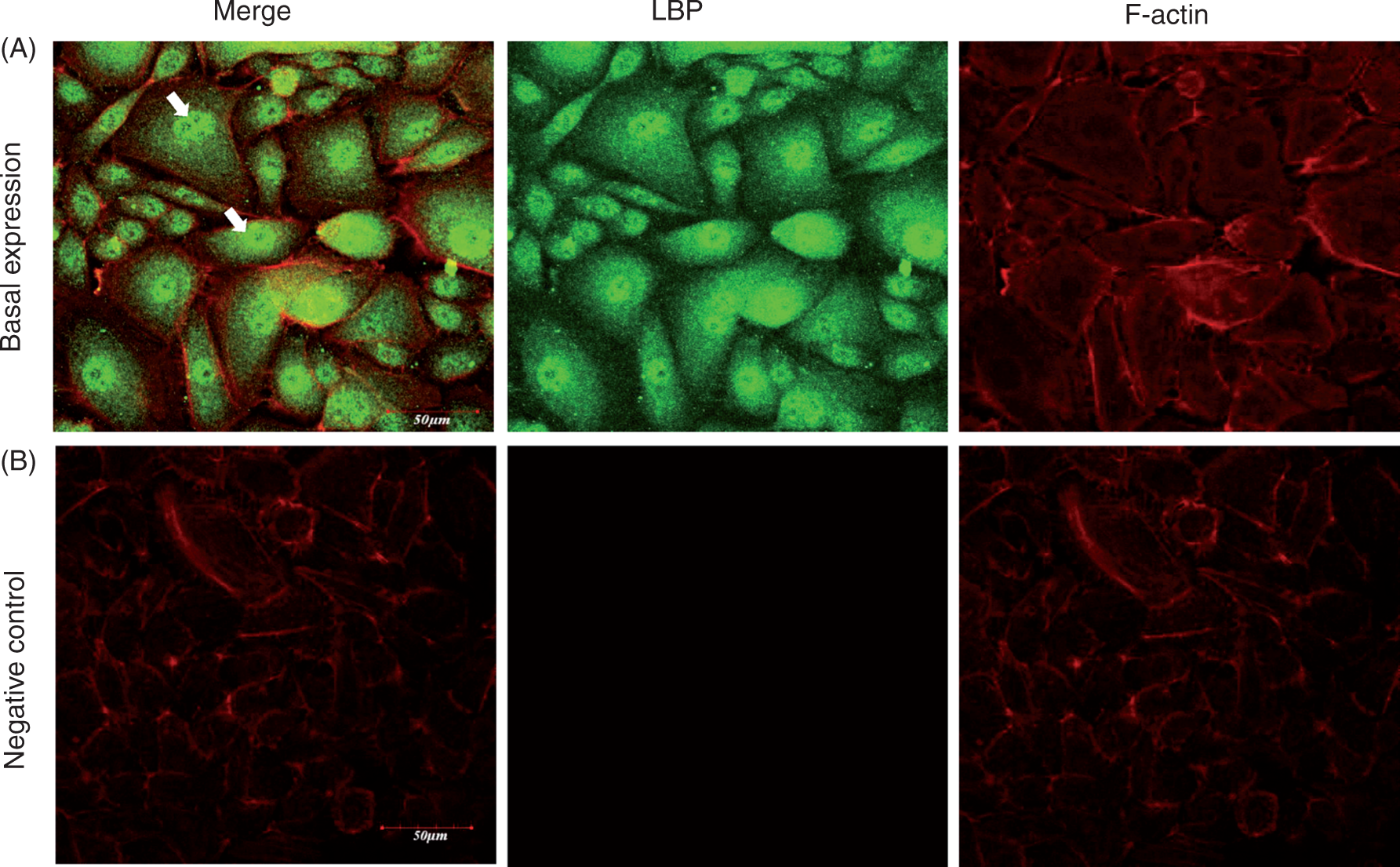
LBP expression differentially modulated by P. gingivalis LPS1690 and LPS1435/1449
Both human LBP mRNA and peptide were expressed basally in HOKs. Escherichia coli and P. gingivalis LPS1690 at 100 ng/ml significantly up-regulated LBP mRNA expression; in contrast, P. gingivalis LPS1435/1449 did not significantly affect the transcript expression at any concentrations examined (Figure 2A). In the time-dependent assay, the LBP transcript level gradually increased and reached the peak at 48 h following the stimulation with either E. coli LPS or P. gingivalis LPS1690 at 100 ng/ml (Figure 2B). Based upon the above results, an optimal concentration of 100 ng/ml for both P. gingivalis LPS and E. coli LPS was selected for the subsequent experiments. As showed in Figures 2C and 2D, Western blot results indicated that LBP peptide was also significantly up-regulated by E. coli LPS and P. gingivalis LPS1690, whereas it was not significantly affected by P. gingivalis LPS1435/1449.
P. gingivalis LPS differentially regulates the expression of LBP mRNA and protein. (A) Expression of LBP mRNA stimulated with various concentrations of LPS for 24 h. P. gingivalis LPS1690 and E. coli LPS up-regulated LBP mRNA at 100 ng/ml, whereas P. gingivalis LPS1435/1449 did not significantly affect LBP transcript expression. (B) Expression of LBP mRNA stimulated with LPS of 100 ng/ml at different time points. P. gingivalis LPS1690 gradually up-regulated LBP transcript level and reached a peak at 48 h. However, the up-regulation of LBP by E. coli LPS was more prompt and the expression reached a peak at 24 h. (C, D) Western blot results showed that LBP peptide was significantly up-regulated by P. gingivalis LPS1690, whereas it was not significantly affected by P. gingivalis LPS1435/1449. Up-regulation of LBP peptide was also observed in E. coli LPS-treated cells. One representative experiment of three is shown. *P < 0.05, **P < 0.001 (reference to control group).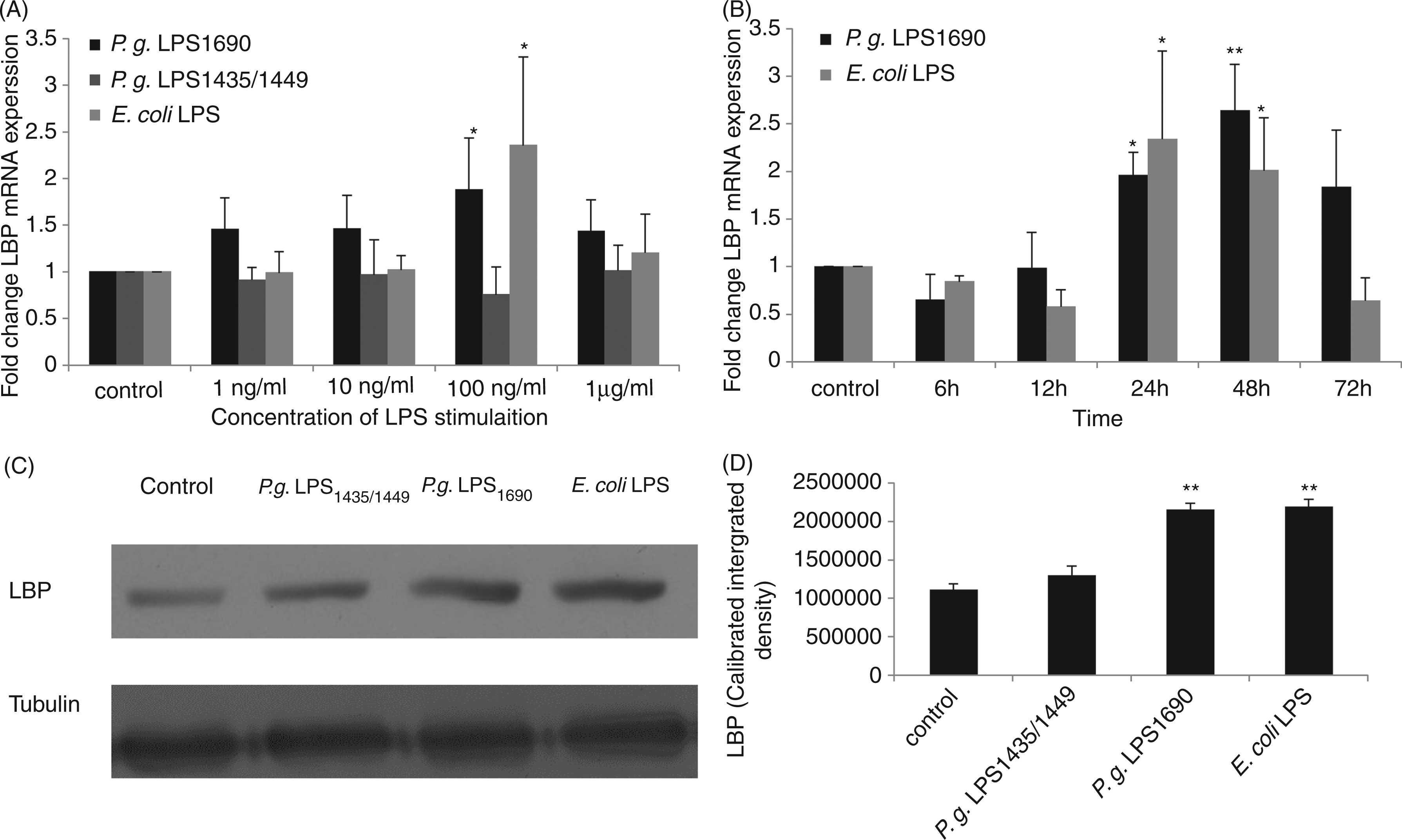
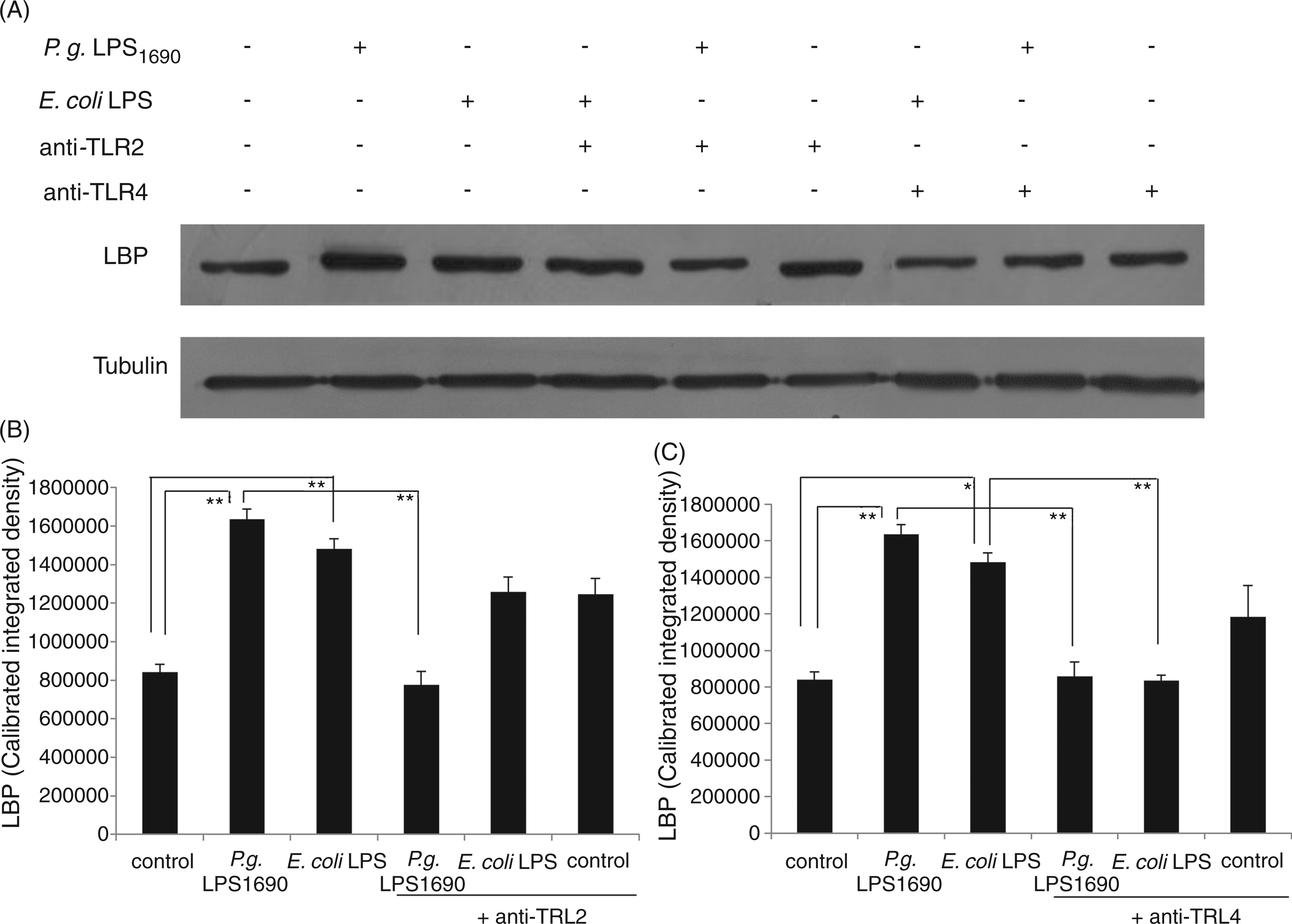
P. gingivalis LPS1690-induced LBP expression neutralized by TLR2 and TLR4 Abs
Pam3Cys as a synthetic lipopeptide and an excellent TLR2 agonist was used to determine the blocking efficiency of anti-TLR2 mAb. Meanwhile, E. coli LPS as a representative TLR4 agonist was employed as the positive control for the efficiency of anti-TLR4 mAb used in the study.
29
IL-6, a well-known marker involved in TLR signalling pathways, served as the outcome measure.
29
Both anti-TLR2 and anti-TLR4 mAbs at 10 µg/ml significantly blocked Pam3Cys- and E. coli LPS-induced IL-6 expression respectively (Figure 3). Therefore, in the subsequent blocking assays, 10 µg/ml of the above Abs was selected to pretreat for 1 h to block TLR2 and TLR4 in HOKs prior to the stimulation of P. gingivalis LPS1690 and E. coli LPS.
Anti-TLR2 and anti-TLR4 mAbs significantly inhibit Pam3Cys and E. coli LPS-induced IL-6 expression. Pam3Cys (B) is a synthetic lipopeptide and is an excellent TLR2 agonist, while E. coli LPS (A) is an optimal TLR4 agonist. They were therefore used as the positive controls for testing the efficiency of anti-TLR2 and anti-TLR4 mAbs respectively. Both mAbs at 10 µg/ml significantly blocked Pam3Cys- and E. coli LPS-induced IL-6 expression. One representative experiment of three is shown. *P < 0.05, **P < 0.001.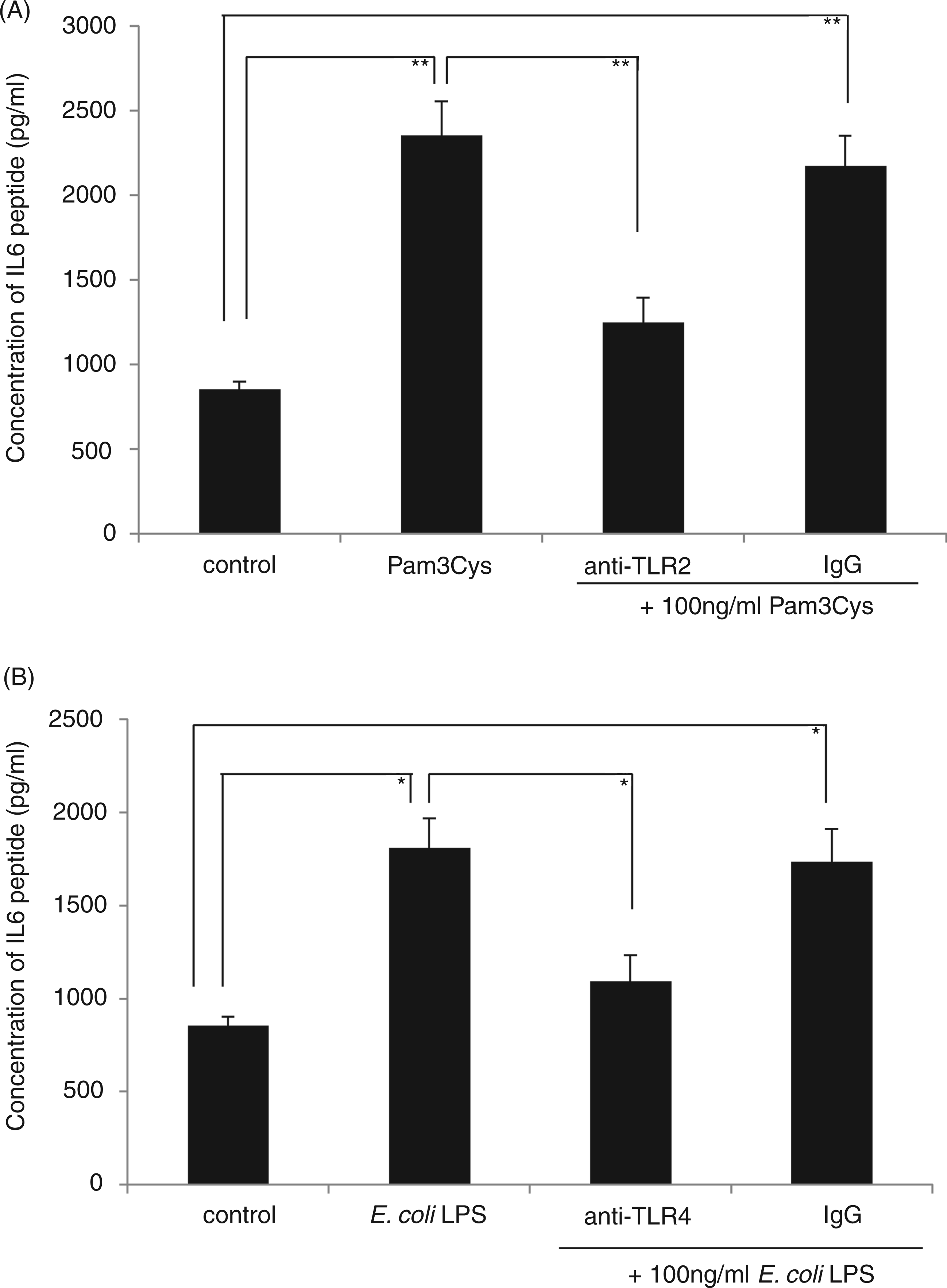
The expression of both TLR2 and TLR4 peptides was significantly up-regulated in HOKs stimulated with P. gingivalis LPS1690, whereas only TLR4 expression increased upon stimulation with E. coli LPS (data not shown). To further confirm whether both TLR2 and TLR4 were involved in P. gingivalis LPS1690-induced LBP expression, anti-TLR2 and anti-TLR4 mAbs were employed. Pre-incubation with TLR2 or TLR4 mAb significantly blocked P. gingivalis LPS1690-induced expression of LBP mRNA at 24 h (Figure 4A) and its peptide at 48 h (Figure 5), while E. coli LPS-induced LBP expression was significantly blocked by TLR4 mAb (Figures 4B and 5).
TLR2 and TLR4 Abs neutralize the modulatory effects of P. gingivalis LPS1690 on LBP mRNA expression. Pre-incubation with TLR2 or TLR4 mAb significantly blocked P. gingivalis LPS1690-induced expression of LBP mRNA at 24 h (A). However, E. coli LPS-induced LBP expression was significantly blocked by TLR4 mAb (B). One representative experiment of three is shown. *P < 0.05, **P < 0.001. TLR2 and TLR4 Abs neutralize the modulatory effects of P. gingivalis LPS1690 on LBP peptide expression. Pre-incubation with TLR2 (B) or TLR4 (C) mAb significantly blocked P. gingivalis LPS1690-induced expression of LBP peptide at 48 h (A), whereas E. coli LPS-induced LBP expression was significantly blocked by TLR4 mAb (A, C). One representative experiment of three is shown. *P < 0.05, **P < 0.001.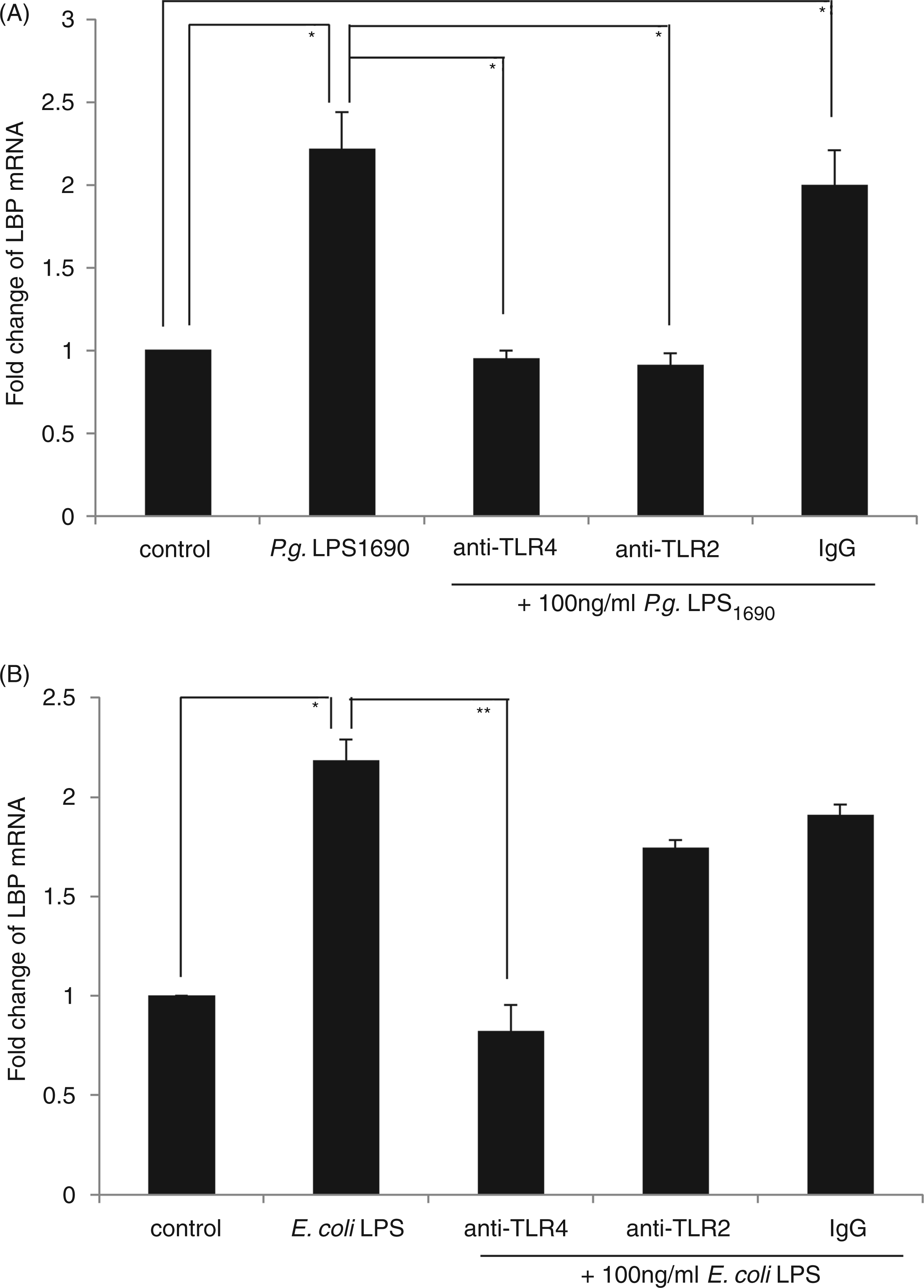
The involvement of TLR2 in P. gingivalis LPS1690-induced LBP expression confirmed by TLR2 siRNA
Much evidence indicates that TLR4, and not TLR2, is involved in host innate response to LPS. To further confirm the involvement of TLR2 in P. gingivalis LPS1690-induced LBP expression, three siRNAs were tested with excellent efficacy. TLR2 mRNA levels were suppressed by around 70% by 5 nM siRNA(#169) in P. gingivalis LPS1690 and E. coli LPS-treated HOKs (Figure 6A). TLR2 siRNA(#169) was then selected to determine if the knock-down of the TLR2 gene would affect the LBP expression observed. Pre-incubation of HOKs with TLR2 siRNA (#169) significantly blocked P. gingivalis LPS1690-induced LBP mRNA expression (P < 0.05), whereas no significant effect was observed on E. coli LPS-induced LBP expression (Figure 6B).
TLR2 siRNA neutralizes the modulatory effects of P. gingivalis LPS1690 on LBP mRNA expression. TLR2 mRNA levels were reduced 70% at 5 nM TLR2 siRNA(#169) in the P. gingivalis LPS1690 and E. coli LPS-treated groups (A). Pre-incubation of HOKs with TLR2 siRNA (#169) significantly blocked P. gingivalis LPS1690-induced LBP mRNA expression (P < 0.05), whereas no significant effect was observed on E. coli LPS-induced LBP expression (B). One representative experiment of three is shown. *P < 0.05.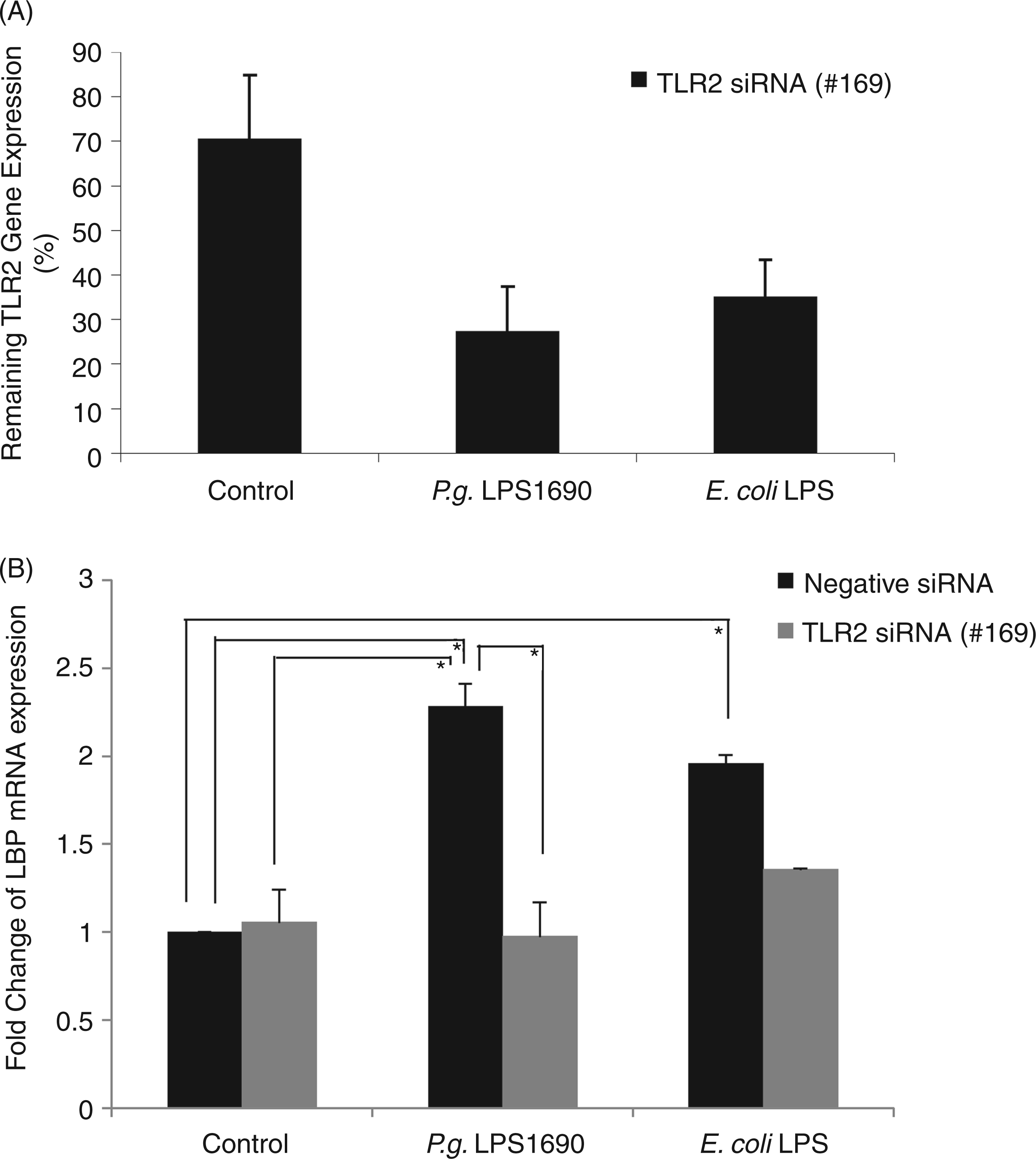
Discussion
LBP is a serum protein that strongly modulates the host response to LPS. 2 LBP has a two-sided effect in the interaction with LPS. 1 In the classic literature, the role of LBP is believed to be in aiding in LPS recognition by transferring LPS to the LPS receptor on the cell surface and subsequently activating adaptive molecules, such as mCD14, TLR4 and MD2. 30 LBP at a lower concentration augments the innate immune response to bacteria and bacterial products other than LPS. 31 In contrast, a higher concentration of LBP can inhibit cellular responses to LPS through neutralizing LPS.7,31 Hence, LBP interacts with LPS in a complex, while orchestrated, manner. Currently, there is little information on how LPS with a lipid A structural heterogeneity could modulate LBP expression in epithelial cells, such as HOKs, and the potential mechanism involved.
Our previous study found, for the first time, that LBP mRNA and protein can be expressed in human gingival tissues and that its mean expression level in periodontally healthy subjects was significantly higher than that of periodontitis patients, which supports the notion that healthy periodontal tissue is armed by the innate host defence system to protect against bacterial infection.14,15 The present in vitro study provides novel information on how the two isoforms of P. gingivalis—LPS1690 and LPS1435/1449—differentially modulate the expression of LBP in HOKs and the involvement of TLR signalling pathways. It is found that both LBP mRNA and peptide are expressed basally in HOKs, in line with its in vivo expression pattern. Furthermore, LBP expression is differently modulated by P. gingivalis LPS with a lipid A structural heterogeneity (i.e. P. gingivalis LPS1690 and LPS1435/1449). This observation suggests that innate immune response in human gingiva could be significantly modulated by different isoforms of P. gingivalis LPS through regulation of the expression of innate defence molecules, such as LBP and human β-defensins (hBD). 32 Our recent study found that the levels of hBD-1, -2 and -3 in the reconstituted human gingival epithelia are significantly up-regulated by P. gingivalis LPS1690, while P. gingivalis LPS1435/1449 down-regulates hBD expression. 32 Recently, it has been realized that the expression of P. gingivalis LPS isoforms may be regulated by hemin in the immediate microenvironment, where tetra-acylated LPS1435/1449 and penta-acylated LPS1690 are produced predominantly under relatively high and low hemin concentrations respectively. 33 It implies that P. gingivalis LPS1435/1449 may render the bacterium able to paralyze the innate host defence and invade the tissues in inflammatory conditions with a high level of hemin.27,34,35 Hence, our current finding provides additional evidence that heterogeneous lipid A structure of P. gingivalis could differentially modulate innate host defence in human gingiva. In addition, it was noted that P. gingivalis LPS-induced LBP expression level peaked at 48 h, while E. coli LPS-induced LBP expression peaked at 24 h. The time discrepancy observed may be due to the difference in structure and biological activities of P. gingivalis LPS and E. coli LPS. P. gingivalis LPS structure lacks heptose, 18 and a synthetic lipid A of P. gingivalis exhibits low endotoxic potency and immunobiological activity with reference to E. coli -type synthetic lipid A. 36 Our recent study showed that gingival epithelia exhibited a relatively prompt responsiveness to E. coli LPS in the expression of hBD-2, while a higher and persistent responsiveness to P. gingivalis LPS1690 was found. 32 Basically, the present results are consistent with this previous observation.
Recognition of LPS is a key event in the innate host response to microbial challenge. LPS-induced signalling in human cells is commonly initiated by CD14, TLRs and MD2.37,38 Currently, at least nine TLR family members have been identified and characterized in humans. 39 Of them, TLR2 and TLR4 have been confirmed to be involved in LPS-induced signalling and expression of immuno-inflammatory molecules. 40 More recent data demonstrate that TLR4, instead of TLR2, is involved in the E. coli LPS signalling.41–43 Yet, unlike E. coli LPS, P. gingivalis LPS activates the innate host response through both TLR2 and TLR4.44,45 This discrepancy is thought to have been attributed to differences in the chemical structure of lipid A. 46 However, Hashimoto et al. 47 found that the TLR2-active component in P. gingivalis LPS resulted from its lipopeptide, and this lipopeptide cannot be removed by the routine phenol re-extraction procedure in the preparation of P. gingivalis LPS. Furthermore, chemically synthesized different forms of lipid A which mimic the natural lipid A portion of LPS from P. gingivalis showed that their activation of host cells was only through TLR4 but not TLR2. 47 However, it has also been shown that trace molecules, such as lipopeptide, are part of the natural LPS of P. gingivalis. 48 In the present study, blocking TLR4 significantly inhibited the modulatory effects of P. gingivalis LPS1690 and E. coli LPS on LBP expression, implying a crucial role of TLR4 in the regulation of LBP by both P. gingivalis LPS1690 and E. coli LPS. Moreover, blocking TLR2 by both TLR2 siRNA and TLR2 Ab also significantly down-regulates P. gingivalis LPS1690-induced LBP expression. This TLR2-mediated effect is known to be due to the lipoprotein component in P. gingivalis LPS. Interestingly, the present study found that only P. gingivalis LPS1690 can significantly up-regulate LBP expression, while P. gingivalis LPS1435/1449 could not affect its expression, although both isoforms of P. gingivalis LPS contain a lipopeptide component. One interpretation is that in contrast to P. gingivalis LPS1690, P. gingivalis LPS1435/1449 weakly activates TLR4 signalling 27 and that this activation is not strong enough to induce LBP expression. Furthermore, P. gingivalis LPS1435/1449 may act as an antagonist to TLR2 for LBP expression in response to lipopeptide; a similar effect has been observed in its inhibition of E-selectin expression as an antagonist in response to E. coli LPS. 27 These results imply that a different lipid A structure of P. gingivalis LPS may influence TLR-dependent LBP expression in HOKs. Further study is required to confirm this hypothesis.
In conclusion, within the limitations of the study, the current findings suggest that P. gingivalis LPS, with a lipid A structural heterogeneity, differentially modulates LBP expression in HOKs in vitro.
Footnotes
Funding
This study was supported by the Hong Kong Research Grants Council (HKU766909M and HKU 768411 M to LJJ).
Acknowledgements
The authors are grateful to Dr Wei Luo from the Faculty of Dentistry, The University of Hong Kong, for his technical assistance.