
Abstract
Response to Gram-negative bacteria (GNB) is partially mediated by the recognition of GNB-derived endotoxin by host cells. Potent host response to endotoxin depends on the sequential interaction of endotoxin with lipopolysaccharide binding protein (LBP), CD14, MD-2 and TLR4. While CD14 facilitates the efficient transfer of endotoxin monomers to MD-2 and MD-2·TLR4, activation of MD-2·TLR4 can occur in the absence of CD14 through an unknown mechanism. Here, we show that incubation of purified endotoxin (E) aggregates (Eagg, M r ≥ 20 million) in PBS with ≥ 0.1% albumin in the absence of divalent cations Ca2+ and Mg2+, yields E·albumin complexes (M r ∼70,000). E·albumin transfers E monomers to sMD-2 or sMD-2·TLR4 ectodomain (TLR4ecd) with a ‘Kd’ of ∼4 nM and induces MD-2·TLR4-dependent, CD14-independent cell activation with a potency only 10-fold less than that of monomeric E·CD14 complexes. Our findings demonstrate, for the first time, a mechanistic basis for delivery of endotoxin monomers to MD-2 and for activation of TLR4 that is independent of CD14.
Introduction
Infection by even small numbers of Gram-negative bacteria (GNB) typically elicits a rapid and robust innate immune response, characterized by the production of pro-inflammatory cytokines (e.g. TNF-α, IL-6) and the recruitment and activation of phagocytes at the site of infection. 1 Endotoxin [E, LPS, lipooligosaccharide (LOS)] is a unique, abundant glycolipid located in the outer leaflet of the outer membrane of GNB (GNBom). 2 Potent host inflammatory responses to endotoxin are mediated by TLR4 and its obligatory co-receptor MD-2, which are expressed on the surface of multiple host cell types.3,4 While exposure to endotoxin facilitates the mobilization of the immune system against invading GNB, continued exposure to high concentrations of endotoxin, or the inability to resolve endotoxin-induced inflammation, can result in severe immunopathologies, such as sepsis.5,6
Potent host response to endotoxin involves the ordered interactions of endotoxin with lipopolysaccharide binding protein (LBP), membrane-bound CD14 (mCD14) or soluble CD14 (sCD14) and extracellular soluble MD-2 (sMD-2) or MD-2 bound to TLR4 (MD-2·TLR4).3,7– 14 LBP binds with high affinity to endotoxin-rich surfaces [e.g. GNBom, aggregates of purified endotoxin (Eagg)], and likely alters the arrangement of endotoxin within this interface.10,15 It increases the exposure of the normally concealed hydrophobic lipid A moiety of endotoxin, catalyzing the extraction of individual endotoxin molecules by soluble or membrane associated CD14. 16 – 19 This results in the formation of monomeric E·CD147,10 which can then rapidly deliver the endotoxin monomer to sMD-2 or to MD-2·TLR4, which results in cellular activation.8,11,12,14,20
While CD14 is required for maximal potency of response to endotoxin, activation of TLR4 by endotoxin can occur in the absence of CD14 through an unknown mechanism. 21 – 24 Given what is known about the requirements for activation of MD-2·TLR4 by endotoxin, it is likely that a CD14-indepenent mechanism also requires the extraction and transfer of an endotoxin monomer from endotoxin-rich interfaces (e.g. GNBom, Eagg) to MD-2·TLR4.8,20,25 This could involve an LBP-independent mechanism to destabilize packing of endotoxin monomers in endotoxin-rich surfaces and a host protein that, like CD14, acts as an endotoxin monomer acceptor/donor (i.e. ‘CD14-surrogate’). This CD14-surrogate would be capable of shuttling endotoxin monomers from disrupted interfaces to MD-2(·TLR4), thereby activating TLR4.
We now show that albumin can serve as a CD14 surrogate for delivery of endotoxin monomers to MD-2(·TLR4) and induces CD14-independent TLR4 activation. The rate-limiting step is extraction and transfer of endotoxin monomers from endotoxin-rich interfaces, as in the CD14-dependent pathway. Unlike CD14, transfer of endotoxin monomers to albumin is not promoted by LBP, but rather by depleting the divalent cations (Ca2+ and Mg2+) necessary to stabilize the dense packing of endotoxin monomers within endotoxin-enriched supra-molecular assemblies. 26 – 28 These results provide new insights concerning the mechanism and molecular requirements of endotoxin recognition by MD-2·TLR4.
Materials and methods
Materials
LBP and sCD14 were gifts from Xoma (Berkeley, CA, USA) and Amgen (Thousand Oaks, CA, USA) respectively. Acyloxyacyl hydrolase (AOAH) was a gift from Dr R. Munford (National Institute of Allergy and Infectious Diseases, Bethesda, MD, USA). Human serum albumin (HSA) was obtained as an endotoxin-free 25% stock solution from Baxter Healthcare (Glendale, CA, USA). HEK293 cells stably expressing TLR4 (HEK TLR4) and their parental cell line (HEK293) were a gift from Dr J. Chow (Eisai Research Institute, Andover, MA, USA). L-929 cell conditioned medium was generated as previously described. 29 [3H]Lipooligosaccharide (25,000 cpm/pmol) was isolated from an aceE mutant of Neisseria meningitidis serogroup B after metabolic labeling as described previously. 30 [14C]Lipopolysaccharide, Rc and S chemotypes, were isolated in the same way after metabolic labeling of aceE Escherichia coli CL99 (a galE mutant of E. coli O111:B4) during growth in medium supplemented with 1,2-[14C]acetate ± 2 mM galactose (manuscript in preparation). 31 Chromatography matrices (Sephacryl HR S200 and Ni2+ FF-sepharose) were purchased from GE Healthcare (Piscataway, NJ, USA). Anti-FLAG M2-agarose was purchased from Sigma (St Louis, MO, USA). Monoclonal Ab (mAb) specific for HSA (anti-HSA) [15C7] (ab10241) and control mouse IgG2b [MPC-11] (ab18469) were purchased from Abcam (Cambridge, UK).
Production of recombinant proteins
Recombinant human FLAG-TLR4 ectodomain wasgenerated by transient transfection of HEK293T cells as previously described. 14 In brief, HEK293T cells were grown in DMEM supplemented with 10% fetal calf serum. Cells in T75 flasks (∼80% confluent) were transfected with 12 µg of an expression vector containing cDNA encoding amino acids 24–631 of TLR4, corresponding to the predicted ectodomain of TLR4 (TLR4ecd) linked to a FLAG tag (pFLAG-CMV-TLR4ecd), using PolyFect reagent (Qiagen, Hilden, Germany). After 12 h, flasks were rinsed in PBS and 8 ml of serum-free medium (293 SFM, Invitrogen, Carlsbad, CA, USA) was added. Medium containing expressed protein was collected 24-48 h later. Medium was concentrated 10–20 fold using Millipore Centricon Plus-70 prior to use.
Recombinant human His6-MD-2 was generated as previously described. 15 Briefly, cDNA encoding MD-2 was inserted into pBAC3 (Novagen, Gibbstown, NJ, USA) to promote the secretion of MD-2 linked to a His6 tag. The generated baculovirus was then amplified in Sf9 cells and used to infect High Five™ (Invitrogen) insect cells in serum-free medium for protein production. Conditioned medium containing secreted His6-MD-2 was used to generate LOS·MD-2 in co-capture experiments and bioassays.
Preparation of [3H]LOSagg and [3H]LOS·protein complexes
[3H]Lipooligosaccharide (or LOS)agg,[3H]LOS·sCD14 and [3H]LOS·MD-2 were generated as previously described. 8 –10,30 Briefly, [3H]LOSagg (M r ≥ 20 million) was obtained after hot phenol extraction of [3H]LOS followed by ethanol precipitation of [3H]LOSagg and ultracentrifugation. Monomeric [3H]LOS·sCD14 (M r ∼60,000) was prepared by treatment of [3H]LOSagg for 30 min at 37°C with substoichiometric amounts of LBP (molar ratio 200:1 of LOS:LBP) and equimolar (to LOS) amounts of sCD14 followed by size-exclusion chromatography (Sephacryl S200, 1.6 × 70 cm column) in PBS, pH 7.4, 0.03% HSA to isolate monomeric [3H]LOS·sCD14 complexes. [3H]Lipooligosaccharide· MD-2 (Mr ∼25,000) was generated by treatment of [3H]LOS·sCD14 with ESF921 (Expression Systems, Woodland, CA, USA) insect cell conditioned media containing His6-MD-2 for 30 min at 37°C followed by isolation of [3H]LOS·MD-2 by S200 chromatography. [3H]Lipooligosaccharide·albumin was generated from sonicated (15 min) [3H]LOSagg that were then incubated for 18 h at 37°C in PBS (no Ca2+/Mg2+) supplemented with 1.0% HSA. Alternatively, sonicated [3H]LOSagg were incubated for 18 h at 37°C in 100 mM Tris-HCl/5 mM EDTA (pH 8.0) supplemented with 1.0% HSA. [3H]Lipooligosaccharide·albumin was isolated by size-exclusion chromatography (Sephacryl S200).
Co-capture of [14C]LOS·albumin with anti-HSA IgG
Mouse monoclonal anti-HSA (non-reactive with BSA) or isotype-matched control mouse IgG2b was coated on 96-well plates at a concentration of 30 µg/ml in 100 µl of 0.1 M sodium carbonate buffer (pH 9.5) by incubation for 18 h at 4°C with shaking. The buffered solution was then removed and the coated wells washed three times with PBS/0.05% Tween-20 (200 µl) followed by incubation for 2 h at 25°C with 100 µl of 3 nM [14C]LOS·albumin or 3 nM [14C]LOS·MD-2 in PBS/1.0% BSA. After this incubation, the supernatant containing [3H]LOS·protein complexes that did not bind to the Ab-coated well was removed from the welland transferred to additional Ab-coated wells to increase the anti-HSA mAb-dependent absorption of [3H]LOS·albumin. This was repeated for a total of five incubations. After the final incubation, the supernatant was removed and 100 µl of aqueous 5% SDS solution was added to each of the washed wells that had been exposed to [3H]LOS·protein complex. The plates were then warmed on a heating block for 10 min to elute the bound [3H]LOS·protein. The radioactivity in the recovered eluate was measured via liquid scintillation spectroscopy.
Aycloxyacyl hydrolase (AOAH) treatment of [3H]LOSagg and [3H]LOS·protein complexes
[3H]Lipooligosaccharideagg or the indicated [3H]LOS· protein (0.5 ng LOS) complexes were incubated in Hank’s buffered salt solution with Ca2+ or Mg2+ (pH 7.4), 0.1% HSA with or without AOAH (7.5 nU/sample, 5.0 ng/µl) for 2 h at 37°C. The extent of deacylation of [3H]LOS by AOAH was monitored by separation of released [3H]-free fatty acids from partially deacylated and remaining intact [3H]LOS by ethanol precipitation of the latter. Ethanol-soluble radioactivity representingreleased [3H]fatty acids was measured by liquid scintillation spectroscopy. The AOAH can release only two of the six fatty acids in LOS. 15 Hence, the extent of partial deacylation of [3H]LOS by AOAH was calculated as % of total cpm recovered in the ethanol-soluble fraction multiplied by three.
HEK293 activation assay
The HEK293 TLR4 cells or their parental cell line were seeded in a 96-well plate (1×105 cells/well) in DMEM/10% FBS/ciprofloxacin (10 µg/ml) and incubated for 18 h at 37°C in 5% CO2 and 95% humidity. The following day, cells were washed twice with warm PBS (pH 7.4) and the indicated treatments were added in 200 µl of DMEM/0.1% HSA. The cells were incubated with the indicated treatments for 18 h at 37°C. Activation of cells was assessed by measuring accumulation of extracellular IL-8 by ELISA (BD Biosciences, San Diego, CA, USA).
Alternatively, HEK293 TLR4 cells were seeded in a 96-well plate (5×104 cells/well) in DMEM/10% FBS/ciprofloxacin (10 µg/ml) and incubated for 18 h at 37°C in 5% CO2 and 95% humidity. The following day, the incubation medium was aspirated and cells were transfected with 0.5 µg/well of either empty vector (pCis-CK) or an MD-2 expression vector (MD-2-FLAG-His6; pEF-BOS) with PolyFect reagent (Qiagen) per manufacturer’s protocol. 32 The following day, the transfection medium was aspirated, 200 µl/well of fresh DMEM/10% FBS/ciprofoxacin was added and the cells were incubated for an additional 24 h to allow for maximal expression of plasmid products. The cells were then washed and treated as described above.
Preparation and activation of CD14KO bone marrow derived macrophages by LOSagg + LBP and LOS·protein complexes
Preparation of bone marrow derived macrophages (BMDM) was carried out as previously described.29,33 Briefly, tibias and femurs from C57BL/6 CD14KO mice were removed following isoflurane asphyxiation and cervical dislocation and the bones were flushed with DMEM using a 23-gauge needle. The flushed cells were grown on 100 × 15 mm dishes in DMEM/10% heat-inactivated FBS/20% L-929 cell conditioned medium/ciprofloxacin (10 µg/ml) for 7 d at 37°C in 5% CO2. On day seven, growth medium was aspirated, cells were dislodged by incubation with versene, seeded in a 48-well plate (2.5 × 105 cells/well) in DMEM/10% heat-inactivated FBS/ciprofloxacin (10 µg/ml), and incubated for 18 h at 37°C. The following day, cells were washed twice with warm PBS (pH 7.4) and incubated with the indicated treatments for 18 h at 37°C in DMEM/0.1% HSA. Activation of cells was assessed by measuring the accumulation of extracellular TNF by ELISA (BD Biosciences).
Reaction of His6-sMD-2 or His6-sMD-2/FLAG-TLR4ecd with either [3H]LOS·albumin or [3H]LOS·sCD14
[3H]Lipooligosaccharide·albumin or [3H]LOS·sCD14 were incubated for 3 h at 37°C at the indicated concentrations with insect cell conditioned medium containingor lacking His6-sMD-2 (4 µl, 0.2 nM active protein), or a combination of 8 µl of insect cell medium containing MD-2 with 25 µl of HEK293 cell medium containing FLAG-TLR4ecd (His6-sMD-2/FLAG-TLR4ecd, 0.15 nM active complex) brought to 0.2 ml (His6-sMD-2 samples) or 0.4 ml (His6-sMD-2/FLAG-TLR4ecd samples) with PBS (no Ca2+/Mg2+)/1.0% HSA, pH 7.4. Following this incubation, Ni2+ FF-sepharose beads (30 µl) to capture [3H]LOS·His6-sMD-2 or anti-FLAG M2-agarose beads (30 µl) to capture [3H]LOS-·His6-sMD-2/FLAG-TLR4ecd were added and incubated at 25°C with rotation for 45 min. Following this incubation, beads were spun-down via centrifugation at 1000 g for 2 min. The supernatant was collected and beads were washed twice in PBS (no Ca2+/Mg2+)/1.0% HSA, pH 7.4. The [3H]LOS was quantified in the recovered supernatant, wash and beads via liquid scintillation spectroscopy. Radioactivity recovered in the beads was converted to molar amounts of product (i.e. [3H]LOS·MD-2 or ([3H]LOS·MD-2·TLR4ecd)2 formed based on the known specific radioactivity of the [3H]LOS. 14 There was no specific capture of [3H]LOS from [3H]LOS·albumin or [3H]LOS·CD14 by anti-FLAG M2 agarose when incubations were carried out with media containing FLAG-TLR4ecd alone. Binding and Scatchard analyses were performed using Prism 5 (GraphPad, La Jolla, CA, USA) Software as described previously. 14
Results
LBP/CD14-independent disaggregation of LOSagg and formation of LOS·sMD-2 are promoted by limiting divalent cations (Ca2+/Mg2+)
The dense packing of endotoxin monomers within the GNBom or Eagg depends on the presence of Ca2+ and/or Mg2+ to reduce electrostatic repulsion between neighboring polyanionic endotoxin molecules.
26
–28,34 Binding of these divalent cations to endotoxin can be reduced by either treatment of endotoxin-rich surfaces with Tris/EDTA (pH 8.0) or by prolonged exposure to media depleted of these divalent cations.
35
–
37
We initially chose the latter conditions to determine whether limiting divalent cations could lead to LBP/CD14-independent disaggregation of purified high molecular mass LOSagg and the transfer of an endotoxin monomer to sMD-2. To facilitate quantitative analysis, we utilized uniformly radiolabeled [3H]LOS (Mr ≥ 20 million) sonicated for 15 min and then further incubated at 37°C in PBS ± Ca2+/Mg2+ (see Materials and methods) and supplemented with 1.0% HSA. Size-exclusion (Sephacryl S200) chromatography was utilized to determine the aggregation state of the [3H]LOS (Figure 1A). In the presence of divalent cations, nearly all of the [3H]LOS eluted in the void volume (V0), reflecting [3H]LOSagg of Mr > 250,000. However, in conditions of limiting Ca2+ and Mg2+, more than half of the [3H]LOS eluted later,at approximately the same elution volume as albumin alone (Figure 1A). Thus, conversion of the large [3H]LOSagg (Mr ≥ 20 million) to much smaller (Mr ∼70,000) complexes was greatly enhanced by incubation in medium lacking Ca2+ and Mg2+. Maximum conversion of the large LOS aggregates to Mr ∼70,000 required incubations for up to 18 h (Figure S1). All subsequent incubations intended to generate the Mr ∼70,000 complex were therefore carried out for 18 h.
LBP/CD14-independent disaggregation of [3H]LOSagg and formation of [3H]LOS·MD-2 requires limiting divalent cations and the presence of albumin. Purified [3H]LOSagg (2 nM) were incubated in PBS with or without Ca2+/Mg2+ for 18 h at 37°C in the presence of 1.0% HSA (A, B) followed by 3 h incubation at 37°C with added sMD-2-containing insect cell conditioned medium (100 ng active protein/ml incubation mixture) (B). Purified [3H]LOSagg (2 nM) were incubated in PBS without Ca2+/Mg2+ for 18 h at 37°C in the presence or absence of 1.0% albumin (C, D) followed by 3 h incubation at 37°C with added sMD-2-containing insect-cell conditioned medium (see above; D). Products were resolved via size-exclusion (Sephacryl S200) chromatography utilizing a 1.6/30 cm column. Radioactivity ([3H]LOS) in the recovered fractions was analyzed via liquid scintillation counting. Data shown are representative of > three independent experiments with overall [3H]LOS recovery ≥ 70%. Dotted lines indicate peak of elution volume of albumin. Void-volume (V0, ∼30 ml), representing [3H]LOSagg is indicated in (A). Arrow indicates peak elution of the [3H]LOS·MD-2 complex at 47 ml in (B).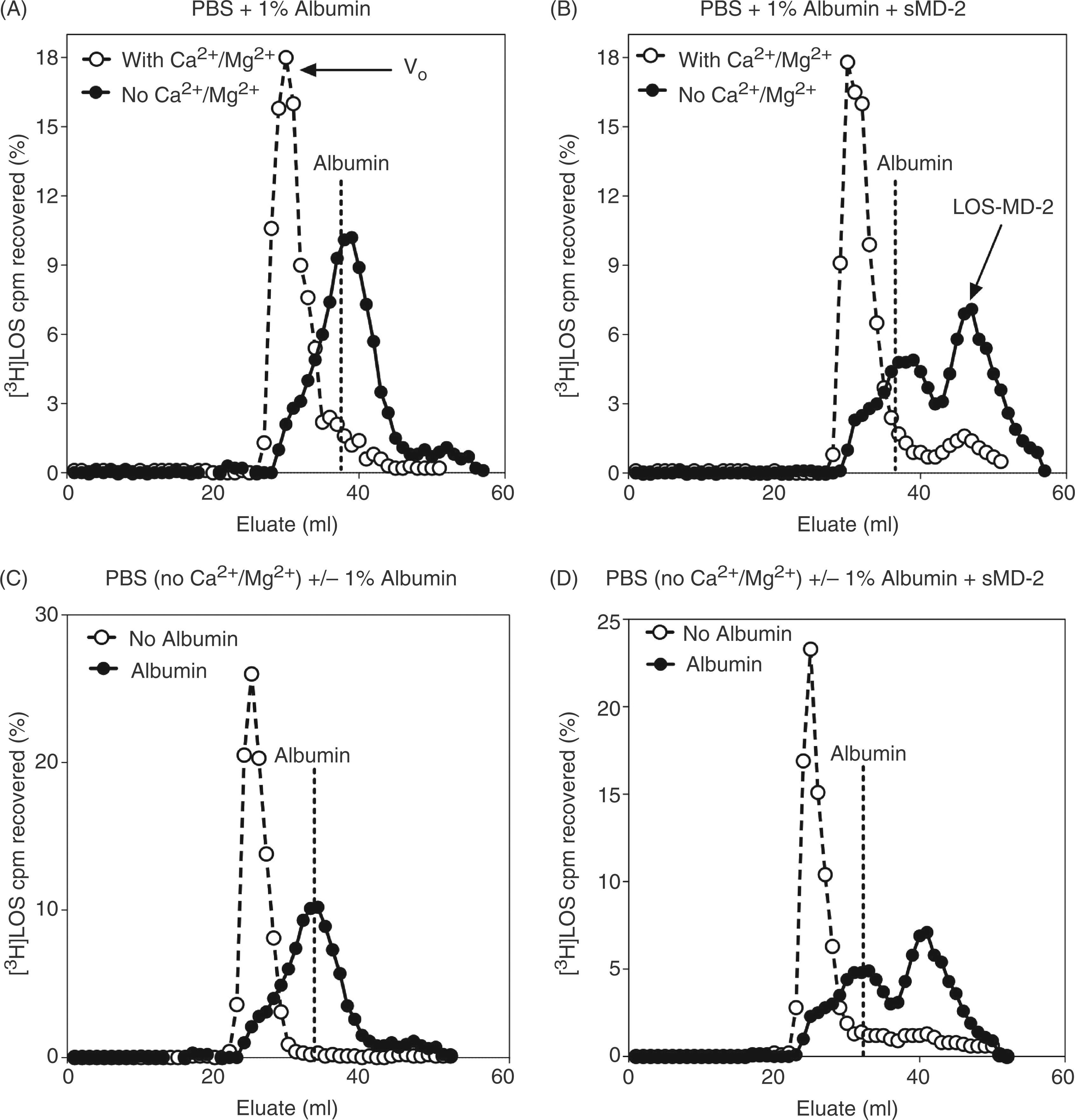
To test whether limiting divalent cations also promoted LBP/CD14-independent transfer of E monomers to sMD-2, [3H]LOS was incubated with insect cell conditioned medium containing sMD-2 after pre-incubation of [3H]LOS in PBS/1.0% HSA/ ± Ca2+ and Mg2+. A new, later eluting (i.e. lower Mr) complex containing [3H]LOS was formed to a much greater extent after the [3H]LOSagg were pre-incubated in PBS without divalent cations (Figure 1B). This later eluting complex corresponded to monomeric [3H]LOS·MD-2 (Mr ∼25,000) as determined by: 1) co-elution with purified LOS·MD-2; 2) the absence of this complex when [3H]LOSagg pre-incubated with PBS/1.0% HSA/± Ca2+and Mg2+ were incubated in control insect cell conditioned medium lacking sMD-2 (Figure S2A); and, 3) dose-dependent activation of HEK TLR4 cells with potency equal to that of purified LOS·MD-2 (Figure S2B). These data show that LOSagg are destabilized when divalent cations are limiting and can form smaller complexes similar in size to albumin. These smaller endotoxin-containing complexes are capable of delivering an endotoxin monomer to sMD-2, thus forming LOS·MD-2.
LBP/CD14-independent disaggregation of LOSagg and formation of LOS·MD-2 requires the presence of albumin
Albumin plays an essential role in LBP/CD14-dependent activation of MD-2·TLR4 by facilitating LBP-catalyzed extraction and transfer of endotoxin monomers to CD14 from endotoxin-rich surfaces and the subsequent transfer of the endotoxin monomer from CD14 to sMD-2 or MD-2·TLR4. 10 To test whether albumin was necessary for the observed LBP/CD14-independent disaggregation of LOSagg and formation of LOS·MD-2, these experiments were repeated in the presence or absence of albumin (Figure 1C, D). As shown, albumin is required for both the conversion of[3H]LOSagg to smaller (Mr ∼70,000) complexes (Figure 1C) and the formation of monomeric [3H]LOS·MD-2 (Figure 1D), from [3H]LOSagg incubated in PBS without Ca2+ and Mg2+ in the presence of sMD-2. This demonstrates that generation of the Mr ∼70,000, endotoxin-containing complex depends on both the destabilization of LOSagg by limiting divalent cations and the presence of albumin (Figure 1). A similar conversion of the large LOSagg complexes to the smaller Mr ∼70,000 complex was observed following incubation of the [3H]LOSagg in a divalent cation-chelating buffer, 100 mM Tris-HCl/5mM EDTA (pH 8.0), supplemented with albumin (Figure S1). Near maximal generation of the Mr ∼70,000 complex was achieved with molar ratios of albumin to LOS as low as ∼1:1 after 18 h incubation at 37°C (data not shown). Aggregates of rough (Rc chemotype) and smooth (S chemotype) LPS similarly generated a Mr ∼70,000 complex under the above conditions (Figure S3A, B), which could react with sMD-2, independent of LBP/CD14, to form LPS·MD-2 complexes (Figure S3C). Whereas LBP catalytically promotes the extraction and transfer of endotoxin monomers from endotoxin aggregates to CD14, the extraction and transfer of endotoxin monomers to albumin was not promoted by the presence of LBP (Figure S4).10,15,17
The Mr ∼70,000 complex is the donor of [3H]LOS monomers to sMD-2
To determine if the Mr ∼70,000 complex could directly transfer endotoxin monomers to sMD-2, this complex was isolated by Sephacryl S200 chromatography after pre-incubation of [3H]LOSagg in PBS/1.0% HSA (no Ca2+/Mg2+). A longer (1.6 x 7.0 cm) S200 column was used to improve resolution of the Mr ∼70,000 complex (second [3H]LOS-containing peak) from the larger [3H]LOSagg eluting in the V0 (Figure 2A). Pooled fractions from the up- and down-slopes of the first (Fx 53-59, corresponding to the V0) and second (Fx 67-73) [3H]LOS-containing peaks were re-analyzed by Sephacryl S200 chromatography to confirm the stability and enrichment of the larger [3H]LOSagg in the V0 and the Mr ∼70,000 complex in the second peak (Figure 2B). These combined fractions were then incubated with sMD-2 at 37°C and the formation of monomeric [3H]LOS·MD-2 was evaluated by Sephacryl S200 chromatography (Figure 2C). The pooled [3H]LOS-containing fractions (Fx 67–73) highly enriched in the Mr ∼70,000 complex reacted with sMD-2 to form monomeric [3H]LOS·MD-2 (Mr ∼25,000). In contrast, very little [3H]LOS·MD-2 was formed under the same incubation conditions when the pooled fractions (Fx 53-59) highly enriched in larger [3H]LOSagg were used (Figure 2C). Under these conditions, the Mr ∼70,000 complex was the preferred donor of monomeric [3H]LOS to sMD-2. The small amount of [3H]LOS·MD-2 formed when incubations were carried out with combined fractions 53–59 may reflect contaminating, incompletely resolved Mr ∼70,000 complex (Figure 2C).
The Mr ∼70,000 complex is a CD14-independent donor of [3H]LOS monomer(s) to sMD-2, yielding [3H]LOS·MD-2. Purified [3H]LOSagg (2 nM) was incubated in PBS (no Ca2+/Mg2+)/1.0% HSA for 18 h at 37°C (A) and reaction products were resolved via size-exclusion (Sephacryl S200) chromatography utilizing a 1.6/70 cm column. The indicated combined fractions from peaks corresponding to LOSagg or Mr ∼70,000 were collected. To test for the purity and stability of these two different [3H]LOS-containing species, an aliquot of each pool was analyzed by S200 size-exclusion chromatography utilizing a 1.6/30 cm column (B). An aliquot of the pooled fractions were then incubated with (see Figure 1 legend) sMD-2-containing insect-cell conditioned medium for 3 h at 37°C. The reaction products were resolved utilizing Sephacryl S200 (1.6/30 cm column) (C). Data shown are representative of > three independent experiments with [3H]LOS recovery at ≥ 75%. Dotted lines indicate peak of albumin elution (A, B, C). Arrow indicates elution of the [3H]LOS·MD-2 complex in (C).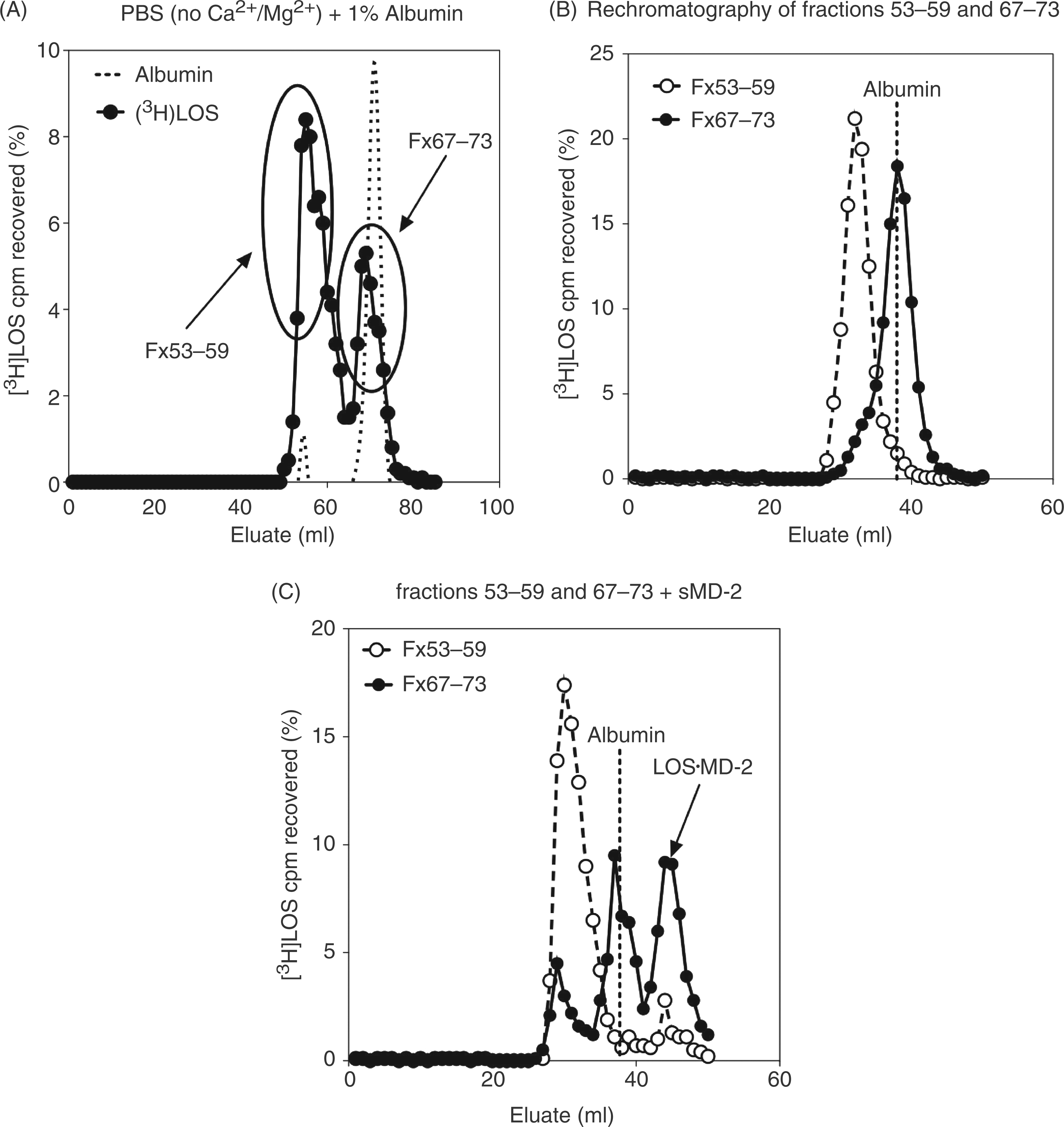
The Mr ∼70,000 complex contains E (LOS) monomer(s) complexed to albumin (LOS·albumin)
The dependence for generation of the Mr ∼70,000 complex on the presence of albumin and the similar elution profile under size exclusion chromatography to that of albumin (Figure 2A, B) strongly suggested that the Mr ∼70,000 peak represents a complex of [3H]LOS monomer(s) (Mr ∼5000) and albumin (Mr ∼65,000). To test this possibility more directly, the Mr ∼70,000 complex was purified followed by assay of the ability of radiolabeled LOS in this complex to be captured by an immobilized anti-HSA mAb compared to an unrelated, isotype-matched mouse IgG2b. As shown in Figure 3, the anti-HSA mAb captured nearly 40% of the radiolabeled LOS contained in the added Mr∼70,000 complex (i.e. pooled fractions of 67-73; Figure 2A), while virtually no capture was seen in wells coated with control mouse IgG2b (Figure 3). There was no capture of [3H]LOS·MD-2 by immobilized anti-HSA (or by the control mouse IgG2b; Figure 3), further demonstrating the specificity of the co-capture by anti-HSA mAb of the radiolabeled LOS in the Mr ∼70,000 complex.
Specific co-capture of radiolabeled LOS in the Mr ∼70,000 complex with anti-HSA Ab. Purified fractions of radiolabeled Mr ∼70,000 or LOS·MD-2 (3 nM) were incubated in 96-well plates pre-coated with either anti-HSA Ab or a control mouse IgG2b as described in 'Materials and methods'. Absorbed radiolabeled LOS was eluted with 2% SDS and measured via liquid scintillation spectroscopy. Data is expressed as % capture of total radiolabeled LOS added. ***P < 0.0001, as calculated by student’s t-test. Data shown are representative of > three independent experiments.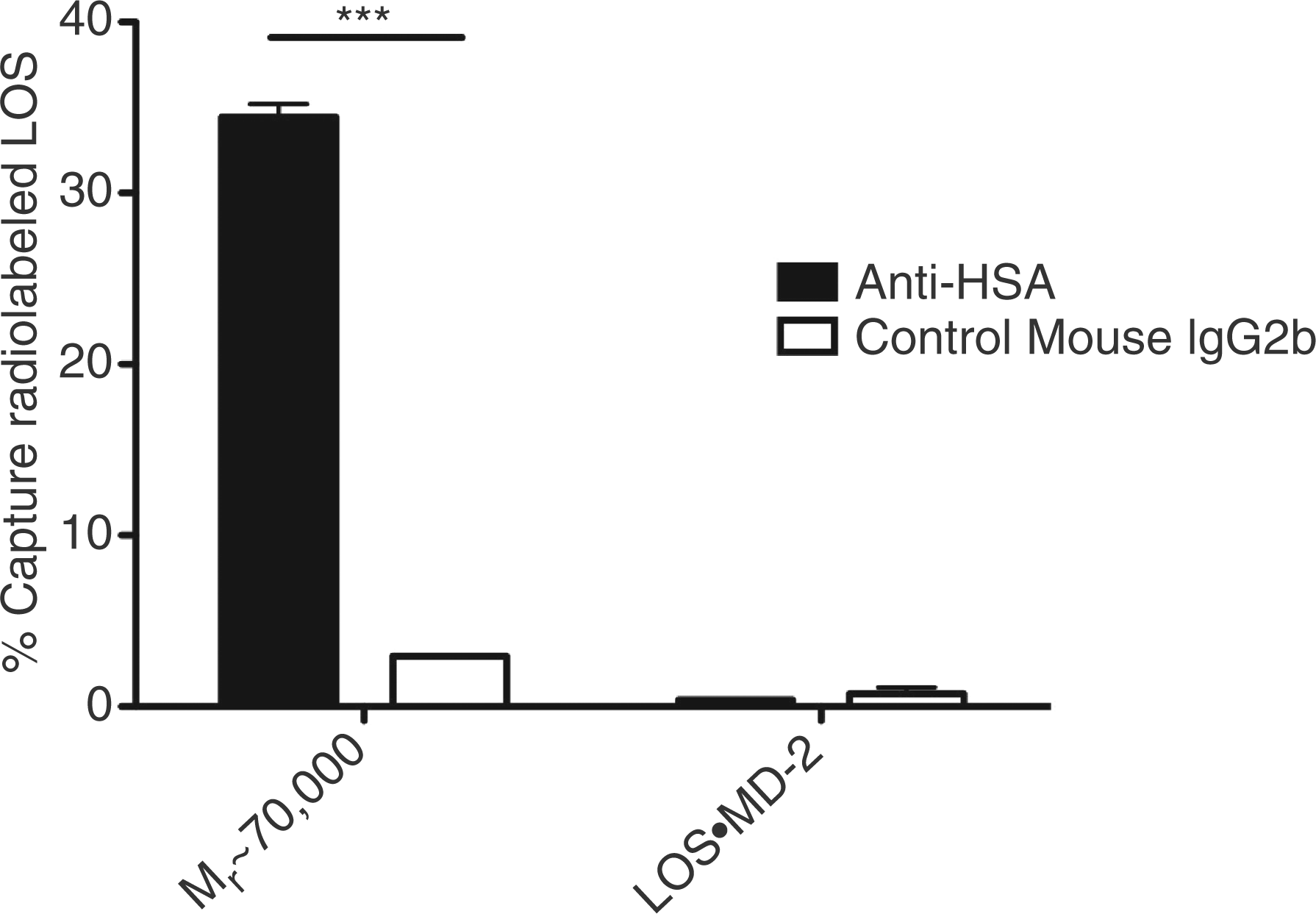
The LOS in the Mr ∼70,000 complex (LOS·albumin) is susceptible to deacylation by AOAH
In the LBP/CD14-dependent pathway of TLR4 activation by endotoxin, endotoxin monomer transfer occurs from Eagg or GNBom treated with very low amounts of LBP (e.g. E:LBP, 200:1 mol/mol) to CD14 and from monomeric E·CD14 to sMD-2 (or to MD-2·TLR4). At each of these steps, endotoxin in the endotoxin donor [i.e. Eagg·LBP (200:1), E·CD14] shows markedly increased susceptibility to the deacylase AOAH.
15
This may reflect increased accessibility of the lipid A fatty acyl chains to AOAH, a property which may favor endotoxin monomer transfer from one endotoxin binding protein to another. The ability of albumin to deliver an endotoxin monomer to MD-2 shown above (Figure 2C) indicates that endotoxin (LOS) is bound to albumin in a way that makes endotoxin monomer transfer possible and may, therefore, manifest increased susceptibility to AOAH. As previously shown, LOS within unmodified LOS aggregates are relatively refractory to AOAH (Figure 4) under physiologicbuffer conditions (see Materials and methods).
15
In contrast, LOS in the Mr∼70,000 complex (LOS·albumin), as in other complexes (e.g. LOSagg·LBP (200:1) and LOS·CD14) that favor LOS monomer transfer, is much more susceptible to AOAH The LOS in the Mr ∼70,000 complex is susceptible to deacylation by AOAH. [3H]Lipooligosaccharide or the indicated [3H]LOS·protein (0.5 ng LOS) complexes were incubated in HBSS/0.1% HSA with AOAH for 2 h at 37°C. The extent of LOS deacylation was determined as described in 'Materials and methods'. Radioactivity recovered in the supernatant after ethanol precipitation from [3H]LOS containing samples incubated without AOAH was < 5% of the total added radioactivity and was subtracted to calculate the % [3H]LOS deacylation by AOAH. The results shown represent the mean ± SEM of three or more determinations.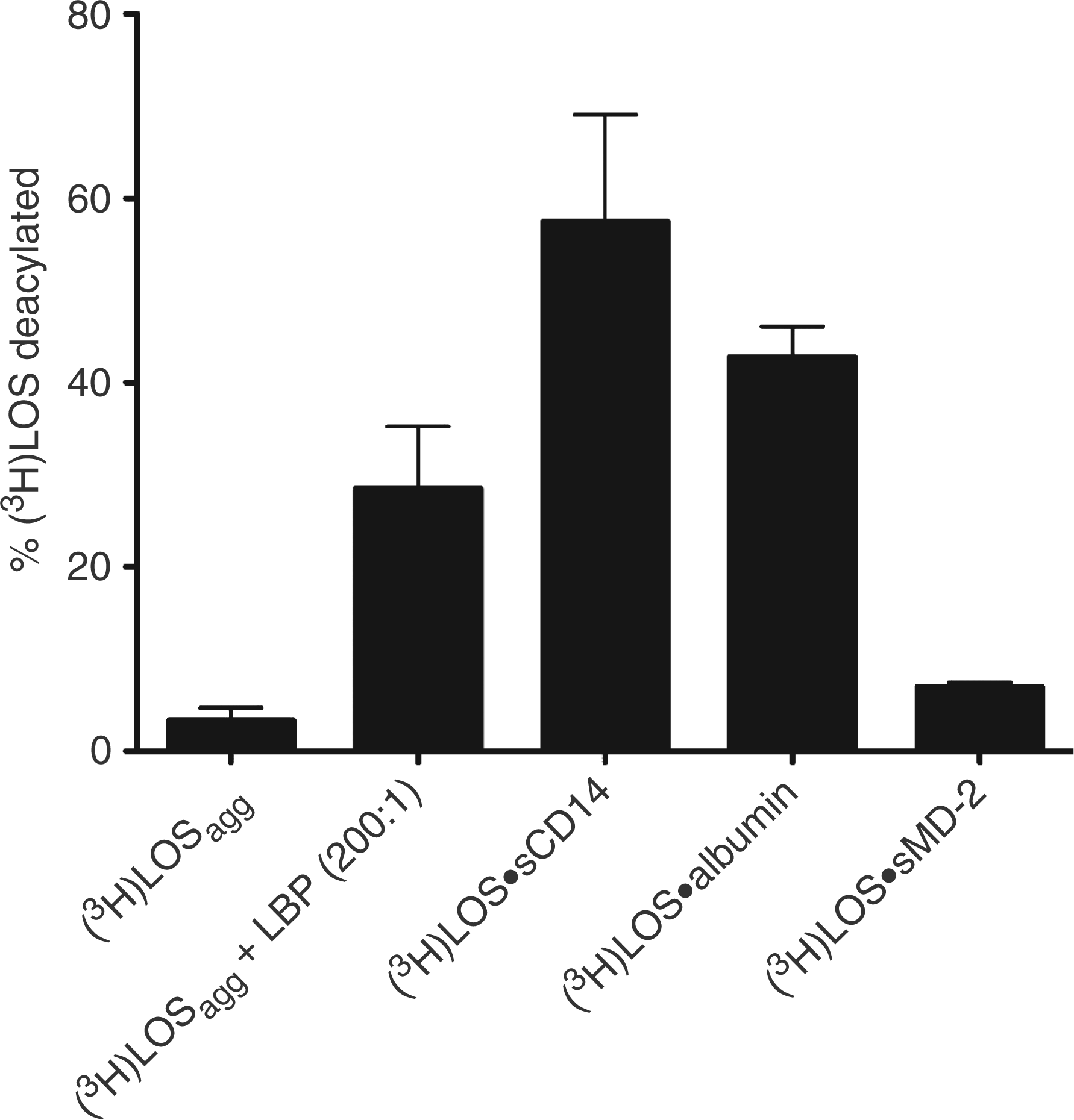
MD-2/TLR4 dependent, CD14-independent cell activation by LOS·albumin
The ability of LOS·albumin to deliver endotoxin (LOS) monomers to sMD-2 (Figures 1 and 2), generating bioactive LOS·MD-2 (Figure S2) strongly suggested that LOS·albumin complexes could provide a CD14-independent mechanism for cell activation. To test this hypothesis, we made use of HEK293 cells (which lack MD-2 and CD14) and tested the ability of LOS·albumin to activate HEK cells that stably express TLR4 (HEK TLR4) in the presence, or absence, of MD-2, monitoring secretion of IL-8 as an indicator of cell activation. Figure 5 shows that LOS·albumin induced robust activation of HEK TLR4 cells provided that MD-2 was also present, either as added sMD-2 in insect cell conditioned medium (Figure 5B) or by transient transfection of HEK TLR4 cells with an expression vector encoding MD-2 (Figure 5A). Little, or no, cell activation by LOS·albumin was observed toward either parental HEK293 cells lacking TLR4 regardless of whether sMD-2 was provided (Figure 5B) or HEK TLR4 cells without sMD-2 (‘empty vector’) or co-expressed MD-2 (Figure 5A). These findings demonstrate that LOS·albumin can activate cells that lack CD14 in an MD-2·TLR4-dependent manner.
CD14-independent, MD-2 dependent activation of HEK/TLR4 by LOS·albumin. HEK293 cells stably expressing TLR4 (HEK TLR4) or the parental HEK293 cell line (HEK) were incubated with LOS·albumin (2 nM) and increasing amounts of either sMD-2-containing insect cell conditioned medium (sMD-2 CM) or insect cell conditioned medium lacking sMD-2 (Ctrl CM) (B) for 18 h at 37°C. Alternatively, HEK TLR4 cells were transiently transfected with either an expression vector encoding human MD-2 (HEK TLR4 + pMD-2) or an empty control vector (HEK TLR4 + empty vector). Following transfection, cells were incubated with increasing amounts of LOS·albumin for 18 h at 37°C (A). Accumulation of IL-8 in culture supernatants was quantified by ELISA as a marker of activation. ***P < 0.0001, **P < 0.005, as calculated by t-test. Data shown are representative of > three independent experiments.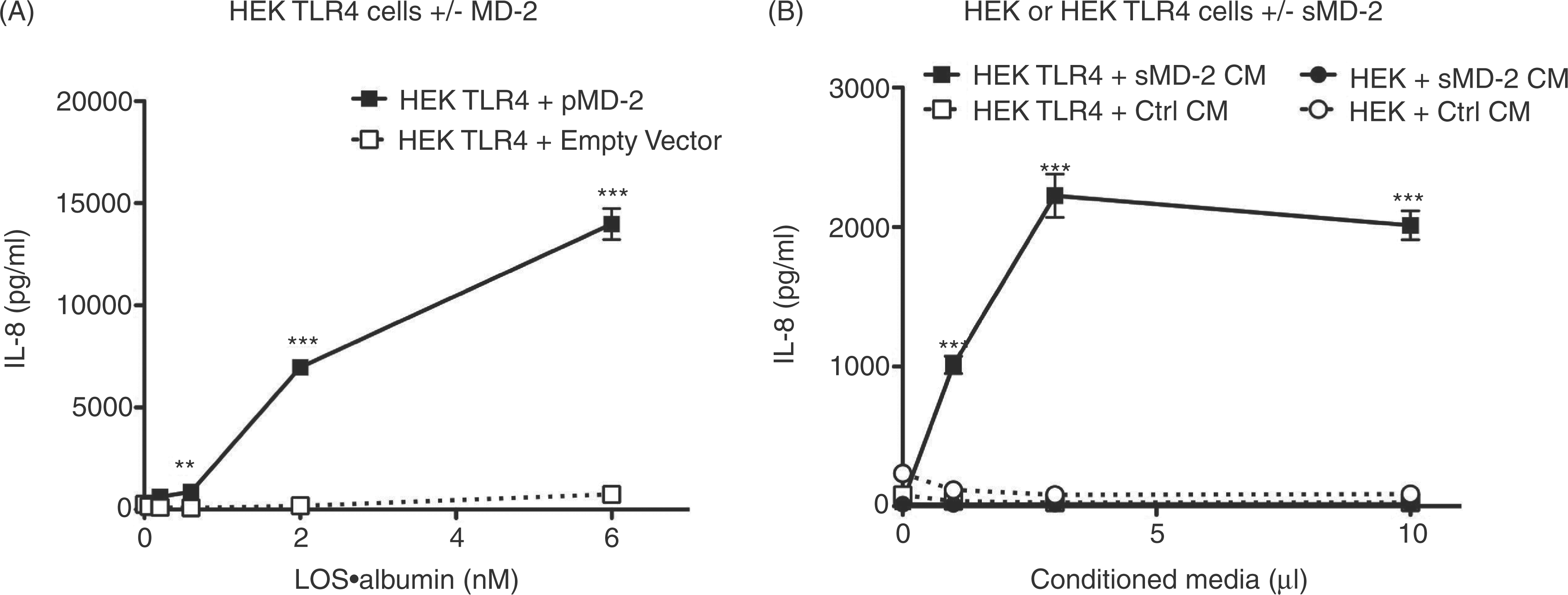
Comparison of LOS·albumin vs. LOS·sCD14 potency in MD-2·TLR4-dependent activation of HEK293 cells
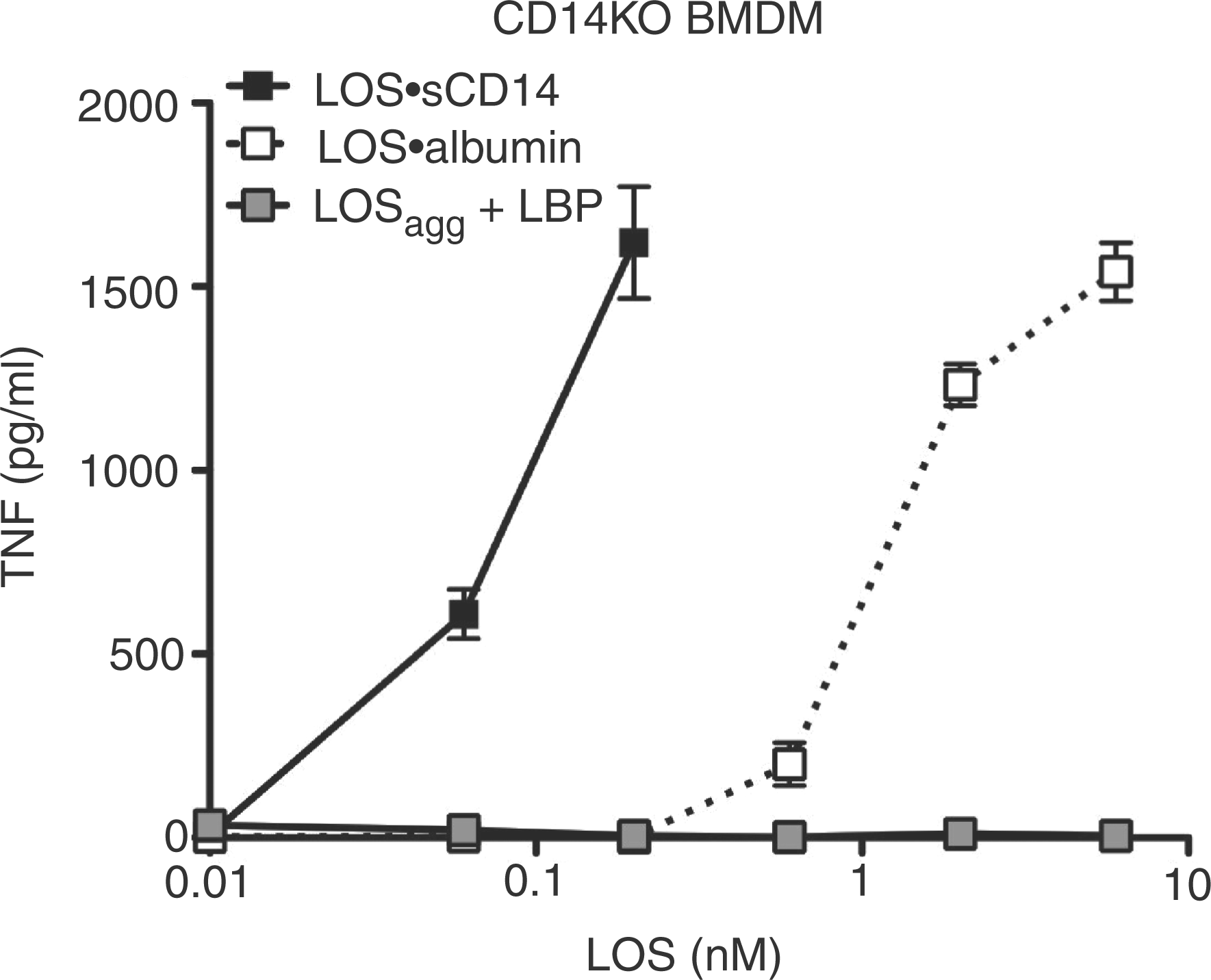
The above findings indicate that LOS·albumin, much like monomeric LOS·CD14 complexes, can induce MD-2·TLR4-dependent cell activation.7,8 To better gauge the relative potency of the LOS·albumin complexes, we directly compared the effects of increasing amounts of LOS·albumin vs. LOS·sCD14 on the activation of HEK/TLR4 cells co-incubated with sMD-2 Comparison of LOS·albumin vs. LOS·sCD14 potency in MD-2·TLR4-dependent activation of HEK cells. HEK TLR4 cells were incubated with increasing amounts of LOS·sCD14 or LOS·albumin in the presence of sMD-2-containing insect cell conditioned medium (A). Alternatively, HEK TLR4 cells were transfected with an expression vector encoding human MD-2 (HEK TLR4/MD-2) and then incubated with increasing amounts of either LOS·sCD14 or LOS·albumin (B). Accumulation of IL-8 in culture supernatants was quantified by ELISA. ***P < 0.0001, **P < 0.005, as calculated by t-test. Data shown are representative of > three independent experiments. CD14-independent activation of CD14KO BMDMs by LOS·albumin. CD14KO BMDMs were incubated with increasing amounts of LOS·albumin, LOS·sCD14 or LOSagg + LBP (10 ng/ml of LBP) at 37°C for 18 h. Accumulation of TNF in the culture supernatants was quantified by ELISA. Results shown represent mean ± SEM of triplicate samples.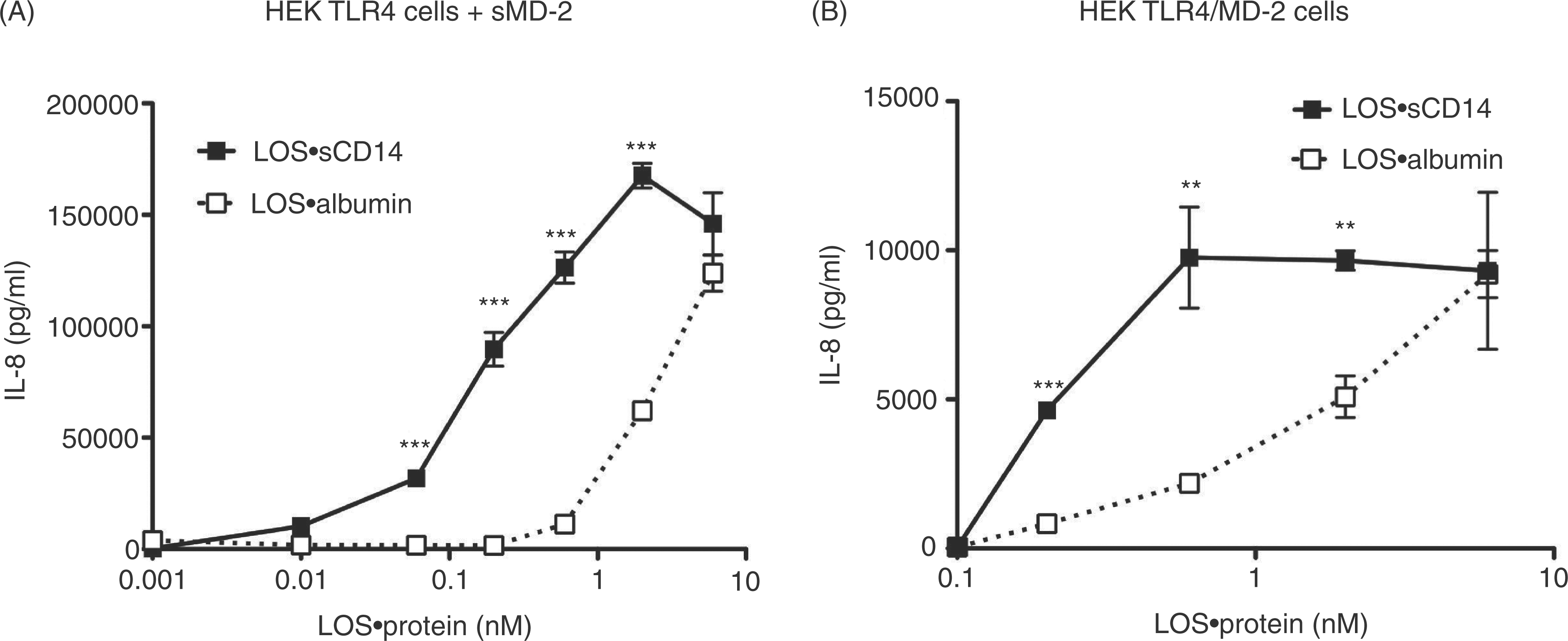
Comparison of [3H]LOS·monomer transfer from [3H]LOS·albumin or [3H]LOS·sCD14 to sMD-2(·TLR4ecd)
The lower potency of LOS·albumin vs. LOS·sCD14 ininducing MD-2·TLR4-dependent cell activation (Figure 6) could reflect lower efficiency (e.g. higher Kd) of transfer of LOS monomer from LOS·albumin vs. LOS·sCD14 to sMD-2 and/or MD-2·TLR4. To test this hypothesis, we took advantage of our ability to sensitively, specifically and quantitatively measure LOS transfer from [3H]LOS·albumin and from [3H]LOS·sCD14 to His6-sMD-2 and to His6-MD-2·FLAG-TLR4ecd by co-capture assays utilizing Ni2+ sepharose beads to capture complexes containing His6-MD-2 or anti-FLAG Ab-coated agarose beads tocapture complexes containing FLAG-TLR4ecd (Figure8).14,32 Preliminary experiments demonstrated that the rate of [3H]LOS transfer was slower from [3H]LOS·albumin than from [3H]LOS·sCD14 (Figure S5). Therefore, the apparent ‘Kd’ of transfer was measured after 3 h incubation, a time sufficient for maximum transfer of LOS from both LOS·albumin and LOS·sCD14 to sMD-2 and sMD-2·TLR4ecd. Scatchard analysis indicated an apparent Kd of 4.0 ± 0.3 nM (n = 4) for [3H]LOS transfer from [3H]LOS·albumin to His6-MD-2 (Figure 8A) vs. an apparent Kd of 0.9 ± 0.2 nM (n = 4) for [3H]LOS transfer from [3H]LOS·sCD14 to His6-MD-2. Similarly, the apparent Kd of [3H]LOS transfer to His6-MD-2·FLAG-TLR4ecd was higher (nearly ninefold) from [3H]LOS·albumin (2.7 ± 0.1 nM; Figure 8B) than from [3H]LOS·sCD14 (0.3 ± 0.1 nM) (Figure 8D). The lower potency of LOS·albumin vs. LOS·sCD14 in stimulating MD-2·TLR4-dependent cell activation thus roughly parallels the lower efficiency of LOS transfer from LOS·albumin vs. LOS·sCD14 to sMD-2 and to sMD-2·TLR4ecd.
Analysis of [3H]LOS transfer from [3H]LOS·albumin or [3H]LOS·sCD14 to His6-sMD-2 or His6-sMD-2·FLAG-TLR4ecd. [3H]Lipooligosaccharide capture to Ni2+ FF-sepharose was measured after incubation of increasing concentrations of [3H]LOS·albumin (A) or [3H]LOS·sCD14 (C) with either His6-sMD-2 (0.2 nM) containing insect cell conditioned medium (total capture, --Δ--) or control insect cell conditioned medium lacking sMD-2 (non-specific capture, --▽--) in PBS (no Ca2+/Mg2+)/1% HSA for 3 h at 37°C. [3H]Lipooligosaccharide co-capture to anti-FLAG agarose was measured after incubation of increasing concentration of [3H]LOS·albumin (B) or [3H]LOS·sCD14 (D) with medium containing His-6-sMD-2·FLAG-TLR4ecd (0.15 nM, total capture, --Δ--) or medium containing FLAG-TLR4ecd (0.15 nM, non-specific capture, --▽--) alone. Specific capture (solid line) was calculated as the difference between total and non-specific capture and used for Scatchard analysis (figure inserts) as described in 'Materials and methods'. Data shown are representative of > three independent experiments.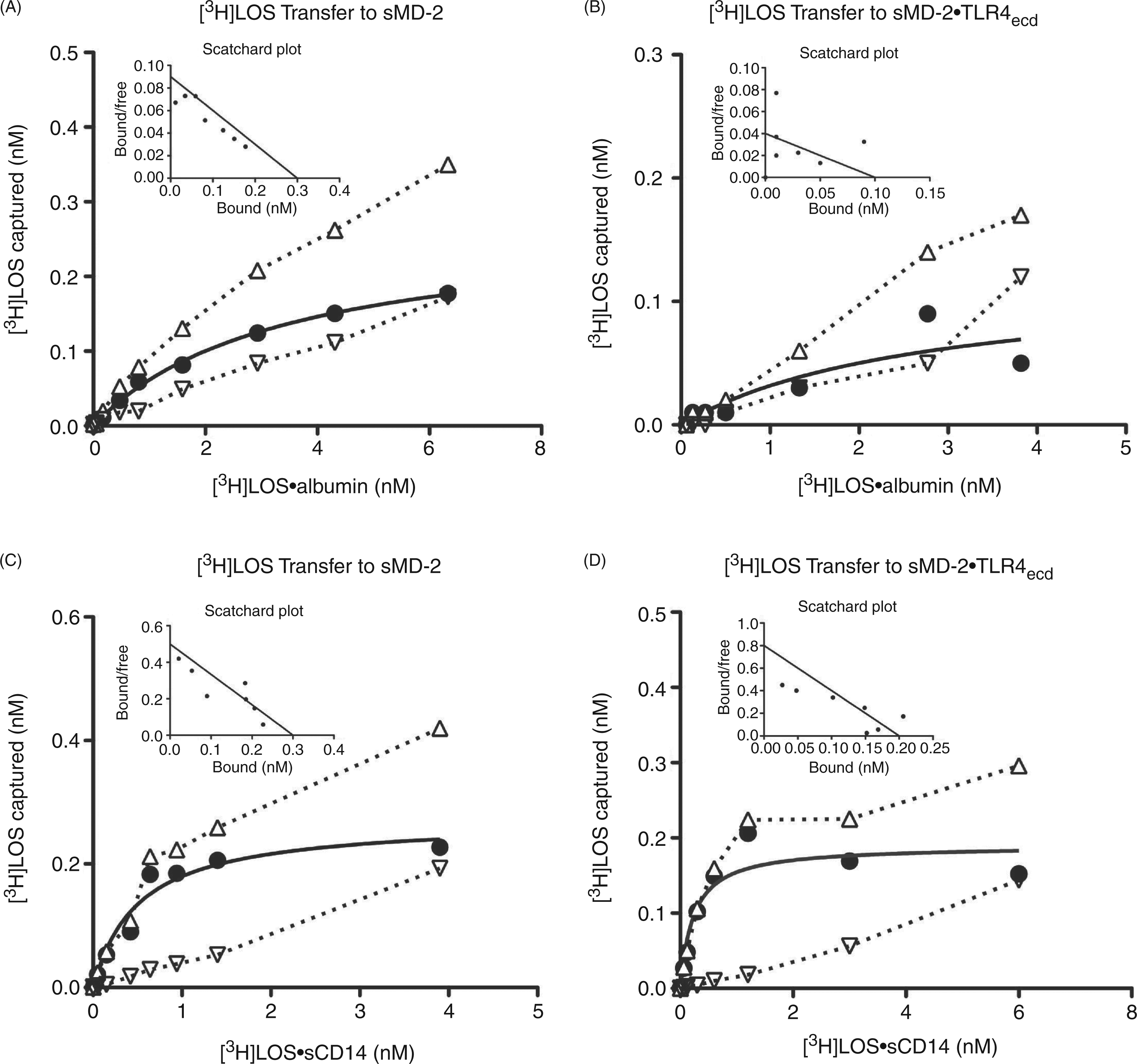
Discussion
Previous work by our laboratory, and others, has shown that mammalian cell activation by endotoxin depends on the sequential interaction of endotoxin with specific host proteins and the delivery of endotoxin monomers to MD-2·TLR4.3,9,14,19,38,39 The best studied and most efficient of these mechanisms involves the formation of monomeric E·CD14 complexes in a process dependent on the presence of LBP and albumin.8,10,17,19 However, activation of TLR4 by endotoxin can occur in the absence of CD14. 21 – 24 As activation of MD-2·TLR4 by endotoxin depends on the binding of a monomer of endotoxin to MD-2, this CD14-independent mechanism would still likely involve the alteration of endotoxin-rich surfaces to favor the extraction and binding of an endotoxin monomer by a CD14-surrogate host protein, which would then transfer the monomeric endotoxin to MD-2(·TLR4).3,8,11,40,41 Here we show, for the first time, a mechanistic basis for the activation of MD-2·TLR4 by endotoxin in the absence of CD14 and LBP utilizing apparently monomeric E·albumin complexes.
It has been known for some time that albumin interacts with endotoxin, although the nature and physiologic consequences of these interactions have remained relatively uncharacterized. 42 – 45 Studies investigating the physical and chemical determinants of endotoxin association with albumin have suggested that binding of endotoxin to albumin is primarily mediated by hydrophobic interactions of the fatty acyl chains of lipid A with one or more hydrophobic fatty acid binding sites within albumin.42,46 As such, the uptake of endotoxin by albumin may reflect albumin’s role as a relatively non-specific transporter of various hydrophobic molecules, rather than a specific and dedicated endotoxin carrier function. Fluorescent displacement assays utilizing dansylsarcosine and warfarin have suggested that the binding of lipid A by albumin occurs in domain III of the protein. 42 Domain III contains two high-affinity fatty acyl chain-binding pockets capable of accommodating long-chain (> 12 C) fatty acids. 47 – 50 However, the mechanism by which albumin gained access to the fatty acids of lipid A, which are normally sequestered within the GNBom or within aggregates/micelles of purified lipid A (endotoxin), and the biologic properties of the resulting endotoxin·albumin complexes could not be elucidated from these earlier studies.15,17,19,28
We have now shown that the depletion or sequestration of the divalent cations necessary for the dense packing of the highly negatively charged endotoxin monomers within endotoxin-rich interfaces such as Eagg (Mr ≥ 20 million) favors the extraction and delivery of endotoxin to albumin and formation of stable E·albumin complexes (Mr ∼70,000) (Figures 1 to 3). Near maximal generation of the Mr ∼70,000 E·albumin complex can occur with molar ratios of endotoxin to albumin approaching 1:1, suggesting that the Mr ∼70,000 complex contains 1 mol of endotoxin (LOS, Mr ∼5000) with 1 mol of albumin (Mr ∼65,000). Importantly, E·albumin is capable of donating endotoxin to sMD-2 or MD-2 bound to TLR4, forming bioactive E·MD-2(·TLR4) complexes (Figures 2 and S2). Endotoxin bound to albumin is susceptible to partial deacylation by AOAH, as is endotoxin bound to CD14, suggesting the acyloxyacyl linkage of the branched fatty acids of the lipid A moiety of endotoxin is at least partially exposed to the aqueous environment when endotoxin is bound to either protein (Figure 4). 15 Partial exposure of lipid A acyl chains may be a necessary characteristic of endotoxin bound to proteins that promote the transfer of endotoxin monomers to downstream endotoxin binding proteins which substantially interact with the acyl chains of lipid A, such as MD-2.15,51 Indeed, like monomeric E·sCD14, apparently monomeric E·albumin complexes are capable of activating cells that express TLR4 in the presence of sMD-2, or cells that express MD-2·TLR4, in an LBP/mCD14-independent manner (Figure 5). 9 Taken together, these data show that albumin functions as a CD14-surrogate, binding endotoxin monomers released from endotoxin-rich interfaces and delivering those endotoxin monomers to MD-2(·TLR4), resulting in TLR4 activation.
Our cell activation data show that E·albumin is ∼10-fold less potent in its ability to activate MD-2·TLR4 than E·sCD14 (Figure 6). This correlates with a ∼4–10-fold higher apparent Kd of endotoxin transfer from E·albumin vs. E·sCD14 to MD-2(·TLR4) and slower transfer kinetics of endotoxin monomers to MD-2(·TLR4) (Figure 8, S5). These differences may reflect more favorable endotoxin presentation by CD14 and/or a role for specific protein-protein (e.g. CD14-MD-2) contacts needed for initial docking of the endotoxin monomer donor to MD-2 and efficient endotoxin transfer.8,52– 54
The differences (∼10-fold) in the potency of MD-2·TLR4 activation by E·albumin vs. E·sCD14 are not nearly as pronounced as differences (∼1000-fold) previously observed in vitro and in vivo when comparing endotoxin potency in CD14-containing vs. CD14-free settings.22,23,55 One possible explanation is that, in those earlier studies, the rate-limiting step in CD14-independent pro-inflammatory action of endotoxin was the extraction and delivery of endotoxin monomers to albumin, rather than the activity of E·albumin per se. Our findings clearly show that neither albumin alone nor albumin in the presence of LBP promotes appreciable formation of bioactive monomeric E·albumin complexes (Figure S4). Therefore, different biologic conditions may be required for the extraction and delivery of endotoxin monomers to albumin and subsequent E monomer delivery to MD-2 (Figure 9). In our study, the transfer of endotoxin monomers to albumin was accomplished by manipulating the incubation media to deplete endotoxin-rich interfaces of stabilizing divalent cations. We speculate that similar disruption of endotoxin packing within the GNBom may be effected at extravascular neutrophil-rich infection sites where cationic antimicrobial proteins, such as bactericidal/permeability-increasing (BPI) protein, cathelicidins and defensins accumulate and target anionic groups in endotoxin that normally bind Ca2+ and Mg2+.
56
–
61
The interactions of these proteins with endotoxin may also reduce LBP-endotoxin interactions, inhibiting LBP-dependent delivery of endotoxin to CD14 and thereby favoring transfer of endotoxin to albumin (which would be present in abundance).
62
The lower levels of plasma lipoproteins at extravascular (vs. intravascular) sites could favor delivery of endotoxin monomers to MD-2(TLR4) by reducing competing endotoxin binding reactions with lipoproteins.
63
This may be particularly important for E·albumin complexes, given the slower rate of transfer of E monomers from albumin (vs. CD14) to MD-2 (Figure S5).
LBP/CD14-independent activation of host cells by E·albumin. Maximum potency of TLR4 activation occurs through the LBP/CD14-dependent pathway (left) when LBP alters the dense packing of endotoxin within Eagg and catalyzes the extraction of an endotoxin monomer by mCD14 or sCD14, which then transfers those monomers to MD-2(·TLR4). Alternatively, LBP/CD14-independent activation of host cells by endotoxin (right) can occur following destabilization of endotoxin aggregates by limiting or chelating divalent cations or perhaps by the action of host defense proteins, such as the complement membrane attack complex or host cationic antimicrobial peptides. This leads to the extraction and binding of endotoxin by albumin. Albumin can serve as a CD14-surrogate by transferring endotoxin monomers to MD·2(·TLR4), resulting in activation of TLR4. As activation of TLR4 by E·albumin occurs in the absence of CD14, it is possible that the resulting intracellular signaling would proceed predominantly through the MyD88-dependent pathway.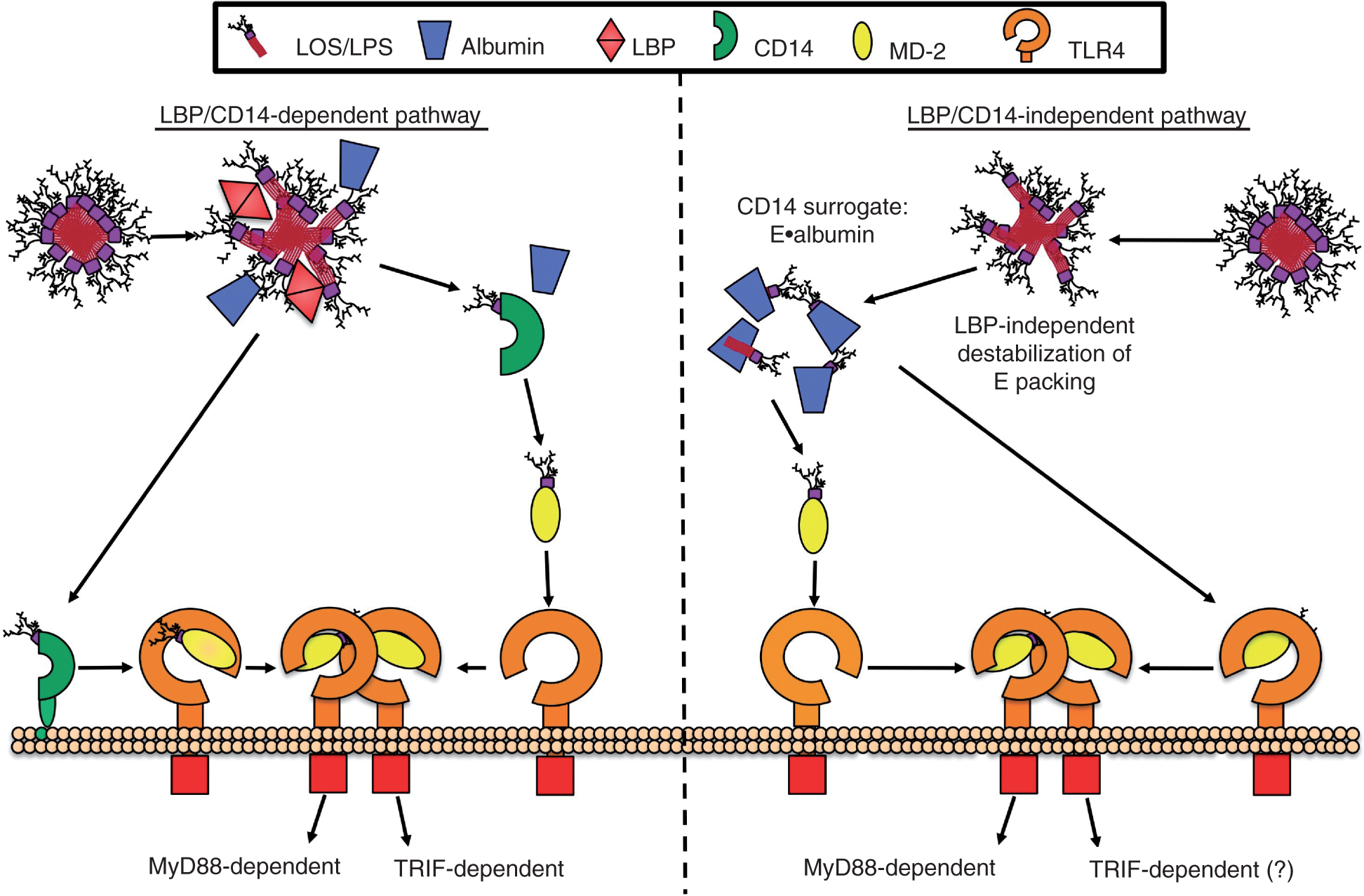
TLR4 is the only TLR capable of triggering two distinct intracellular signaling pathways: the MyD88-dependent pathway, which utilizes the adaptor molecules MyD88 and Mal and results in production of pro-inflammatory cytokines, such as TNF; and the TRIF-dependent pathway, which signals through the adaptor molecules TRIF and TRAM and results in theproduction of type 1 interferons. 64 – 67 The concerted action of these pathways helps produce an effective and appropriate immune response during most GNB infections. The mechanisms that govern the initiation and magnitude of activation of each TLR4 signaling pathway remain unclear. A role for CD14 in facilitating TRIF-dependent TLR4 signaling in response to endotoxin has been shown, but these observations have thus far been limited to macrophages from wt- and CD14-null mice activated with a narrow range of endotoxin species (i.e. lipid A and Re LPS). 68 The ability of E·albumin to activate MD-2·TLR4 independently of CD14, combined with the capacity of E·albumin to bind both R and S species of endotoxin (Figure S3) should provide a means for a more comprehensive analysis of mammalian host cell TLR4 activation by endotoxin in the absence of CD14 and the effects of this stimulation on subsequent activation of TRIF and/or MyD88-dependent signaling.
Footnotes
Acknowledgements
This work was supported by U.S. Public Health Service Grant A105732 (to J.P.W.), a Department of Veterans Affairs, Veterans Health Administration, Office of Research and Development grant (to T.L.G.), and the University of Iowa Dean's Graduate Fellowship and a Training in Mechanisms of Parasitism grant T32 A10075 (to G.E.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.