
Abstract
We investigated the ability of tetherin, a recently identified antiviral factor, in restricting hepatitis C virus (HCV) in the Japanese fulminant hepatitis-1 (JFH-1) infectious cell culture system. Human hepatocytes (Huh7, Huh7.5.1) expressedlow levels of endogenous tetherin, which could be induced by IFN-α. However, tetherin contributes little to IFN-α-mediated anti-HCV JFH-1 activity. Although tetherin could inhibit Vpu-deleted HIV-1 release, it had negligible activity in restricting HCV JFH-1 release from hepatocytes, which was evidenced by unaffected levels of intracellular/extracellular HCV RNA and infectious virus. The failure of tetherin’s anti-HCV activity could not be related to the counteraction of HCV, as HCV infection of hepatocytes affected neither tetherin expression nor anti-HIV function of tetherin. These observations imply that tetherin has negligible activity in the restriction of HCV JFH-1 in human hepatocytes.
Introduction
A number of cellular factors that restrict viral replication through interactions with viral components have been recently identified.1–5 Among these factors, tetherin (also known as BST2, CD317 and HM1.24) is known to have the ability of inhibiting HIV-1 release.2,6 Tetherin has antiviral activity against a broad range of viruses, including all retroviruses (alpha-, beta-, delta-, spuma- and lentiviruses),2,6,7 arenaviruses (Lassa), 8 filoviruses (Ebola, Marburg)7,8 and Kaposi’s sarcoma-associated herpesvirus (KSHV).9,10 Tetherin is constitutively expressed in terminally differentiated B cells, bone marrow stromal cells and plasmacytoid dendritic cells, and can be induced by type I and type II IFNs in various cells.11,12 Therefore, tetherin is currently considered as an important component of antiviral innate immune defence. 13
Tetherin is found in both the plasma membrane and within the endosomal system, including the trans-Golgi network.14–16 The inhibitory mechanism by which tetherin functions is still largely unclear and is believed to be non-specific as tetherin can inhibit the release of a broad spectrum of viruses that are known either to bud from cholesterol-enriched domains of the plasma membrane, such as HIV and Ebola 2,7 or to envelope the capsids at the endoplasmic reticulum and trans-Golgi network, followed by transport to the cell surface in secretory vesicles, such as herpesvirus KSHV. 17 These viruses are all lipid-enveloped and but share few common features. 13 Hepatitis C virus (HCV) is an enveloped RNA virus belonging to the family Flaviviridae, genus Hepacivirus. 18 HCV replication occurs on a lipid raft membrane structure that requires the existence of cholesterol.19–21 It has also been proposed that infectious HCV particles are assembled when its genomic RNA-containing core particles bud through the endoplasmic reticulum membrane, 22 obtaining viral envelope and surface glycoproteins, followed by transportation from the endoplasmic reticulum through the trans-Golgi network to the secretory pathway.23,24 These biological features of HCV suggest that the virus would be sensitive to tetherin. Thus, we used HCV genotype 2a Japanese fulminant hepatitis-1 (JFH-1) strain to investigate whether HCV release from hepatocytes can be restricted by tetherin and whether HCV JFH-1 has the ability to counteract the antiviral function of tetherin.
Materials and methods
Reagents
Recombinant IFN-α was purchased from R&D Systems Inc. (Minneapolis, MN, USA). Mouse anti-HCV core Ab was purchased from ABR Affinity BioReagents, Thermo Scientific (Rockford, IL, USA) and rabbit anti-HA Ab from Sigma-Aldrich (St Louis, MO, USA).
Cell cultures and HCV infection
Huh7, Huh7.5.1 and Hela cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal calf serum (FCS). The generation of infectious HCV JFH-1 and infection of Huh7 [multiplicity of infection (MOI) of 0.01)] were carried out as described.25,26
Quantitative real-time RT-PCR
Total cellular RNA was extracted from cells using Tri-Reagent (Molecular Research Center, Cincinnati, OH, USA). The real-time RT-PCR for the quantification of cellular HCV RNA 27 and tetherin 28 was performed with the iQ SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA). The levels of GAPDH mRNA were used as an endogenous reference to normalize the quantities of target mRNA. Total RNA in cell culture supernatant was extracted with Tri-Reagent-BD (Sigma-Aldrich) in accordance with the manufacturer’s instructions. The real time RT-PCR assay developed 29,30 was used for the quantification of HCV RNA in culture supernatant.
Titration of extracellular and intracellular infectious HCV
The HCV JFH-1 infectivity was determined in Huh7 cells by end point dilution and immunofluorescence as described previously 26 with minor modifications. 31 For titration of extracellular infectious HCV, 25 µl supernatant was serially diluted 10-fold in DMEM-10% FCS. For titration of intracellular infectious HCV, 25 µl cell lysate was used to measure the infectious virus as described.26,32 The virus titer is expressed as focus forming units per milliliter (FFU/ml).
Transfection
The plasmid pCR-tetherin-HA (1 µg; encoding tetherin with a HA tag) 2 was transfected to Huh7 cells in 24-well plates via GenJet™ in vitro DNA transfection reagent for Huh7 cells (SignaGen Laboratories, Gaithersburg, MD, USA). The cotransfection of either plasmids pNL4.3 (full-length HIV-1, 0.6 µg) with tetherin-HA (0.4 µg), or plasmids pNL4.3-delVpu (Vpu-deleted HIV-1, 0.6 µg) with tetherin-HA (0.4 µg) to Huh7 cells in 24-well plates was also carried out via GenJet™ transfection reagent. Tetherin-specific siRNAs (L-01187; Dharmacon Smartpool,Lafayette, CO, USA ) and control siRNAs were transfected to Huh7 cells using the transfection reagent (DharmaFect 3; Dharmacon) in accordance with the manufacturer’s instructions. Forty-eight hours after transfection, the cells or culture supernatant were either evaluated by immunofluorescence staining, for RNA quantification by real time RT-PCR or for HIV reverse transcriptase (RT) activity by HIV RT assay.
Immunofluorescence
Mouse anti-HCV core and rabbit anti-HA Abs were used for immunofluorescent evaluation of HCV core protein and tetherin expression respectively. Immunofluorescence staining was carried out as previously described. 33
HIV reverse transcriptase assay
Culture supernatant containing HIV-1 was collected as described above. Extracellular HIV RT activity in the supernatant was determined based on the technique of Willey et al., 34 with modification. 35 For analysis of intracellular HIV RT activity, cells were washed once with PBS and then incubated with 50 mmol/l Tris/HCl (pH 8.0), 150 mmol/l NaCl, 5 mmol/l CaCl2 and 1 mg/ml subtilisin for 30 min at 37°C. The reaction was then stopped with 0.5 ml 10% DMEM containing 5 mmol/l phenylmethylsulfonyl fluoride (PMSF). The supernatant was filtered (0.2 µm) and HIV RT activity was measured as described above.
Results
Tetherin plays little role in IFN-α-mediated anti-HCV activity
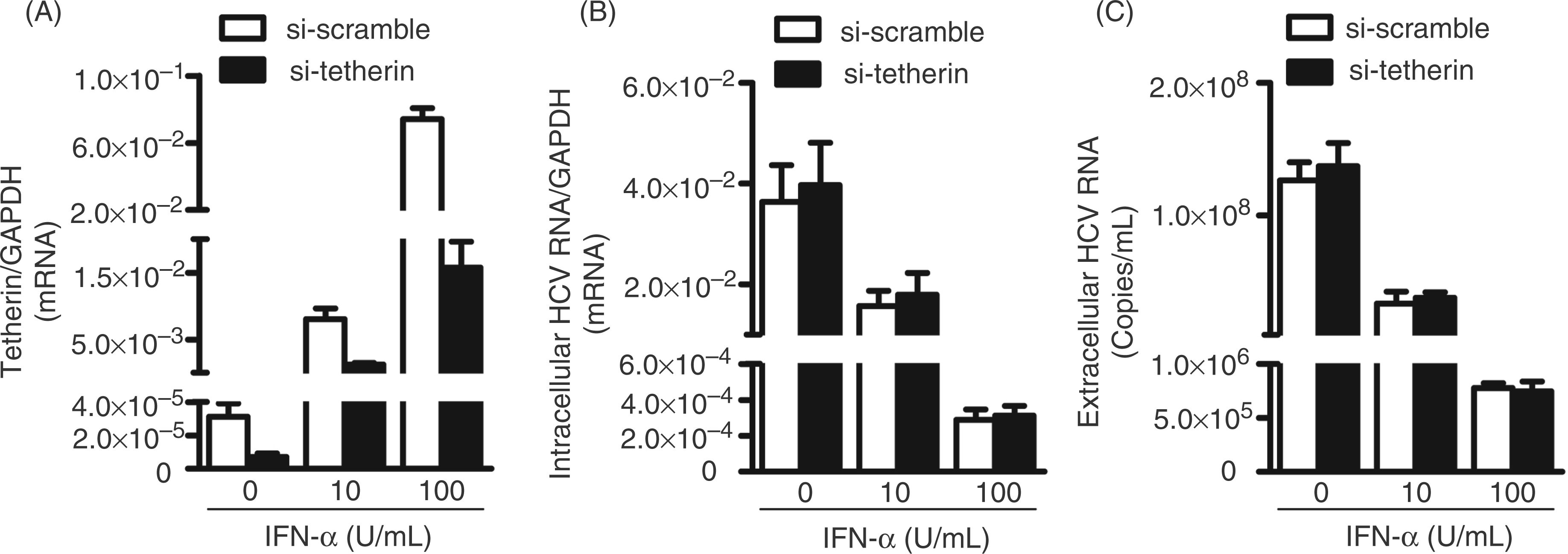
We first analyzed endogenous tetherin expression in human hepatocytes (Huh7 and Huh7.5.1). Both Huh7 and Huh7.5.1 cells expressed significantly lower levels of tetherin mRNA than Hela cells (positive control; Figure 1A). However, IFN-α treatment of Huh7 cells greatly induced tetherin mRNA expression (Figure 1B). As expected, IFN-α treatment of HCV-infected cells significantly inhibited viral RNA expression, either within cells (Figure 1C) or in the supernatant (Figure 1D). To determine whether tetherin plays a role in anti-HCV activity of IFN-α, we used the siRNAs specifically against tetherin to suppress the expression of tetherin induced by IFN-α. As shown in Figure 2A, transfection of IFN-α-treated Huh7 cells with the siRNAs against tetherin downregulated the expression of tetherin. However, this downregulation of tetherin had little effect on HCV RNA expression, either within cells (Figure 2B) or in the supernatant (Figure 2C).
Endogenous tetherin expression in human hepatocytes. (A) Endogenous tetherin expression in Huh7 and Huh7.5.1 cells. Total cellular RNA extracted from Hela (positive control), Huh7 and Huh7.5.1 cells was subjected to the real-time RT-PCR. The relative ratio of the tetherin mRNA to GAPDH mRNA is given to quantification of tetherin expression. (B–D) Effect of IFN-α on tetherin and HCV expression in Huh 7 cells. At 72 h post-infection, HCV JFH-1-infected Huh 7 cells were treated with recombinant IFN-α at the indicated concentrations for 48 h. The levels of tetherin (mRNA) (B), intracellular (C) or extracellular (D) HCV RNA were determined by real-time RT-PCR. The levels of tetherin and intracellular HCV RNA were expressed as relative ratios to GAPDH mRNA. The levels of extracellular HCV RNA are expressed as copies/ml. The results shown in A–D are the mean ± SD of triplicate cultures, representative of three experiments. Effect of IFN-α-induced tetherin on and HCV. At 72 h post-infection, HCV JFH-1-infected Huh7 cells were treated with recombinant IFN-α at indicated concentrations. Simultaneously, siRNAs specific to tetherin or the nontargeting scrambled siRNA were transfected into the cells. The levels of tetherin (mRNA) (A), intracellular (B) or extracellular (C) HCV RNA were determined by real-time RT-PCR. The levels of tetherin and intracellular HCV RNA are expressed as relative ratios to GAPDH mRNA. The levels of extracellular HCV RNA are expressed as copies/ml. The results shown in A–C are the mean ± SD of triplicate cultures, representative of three experiments.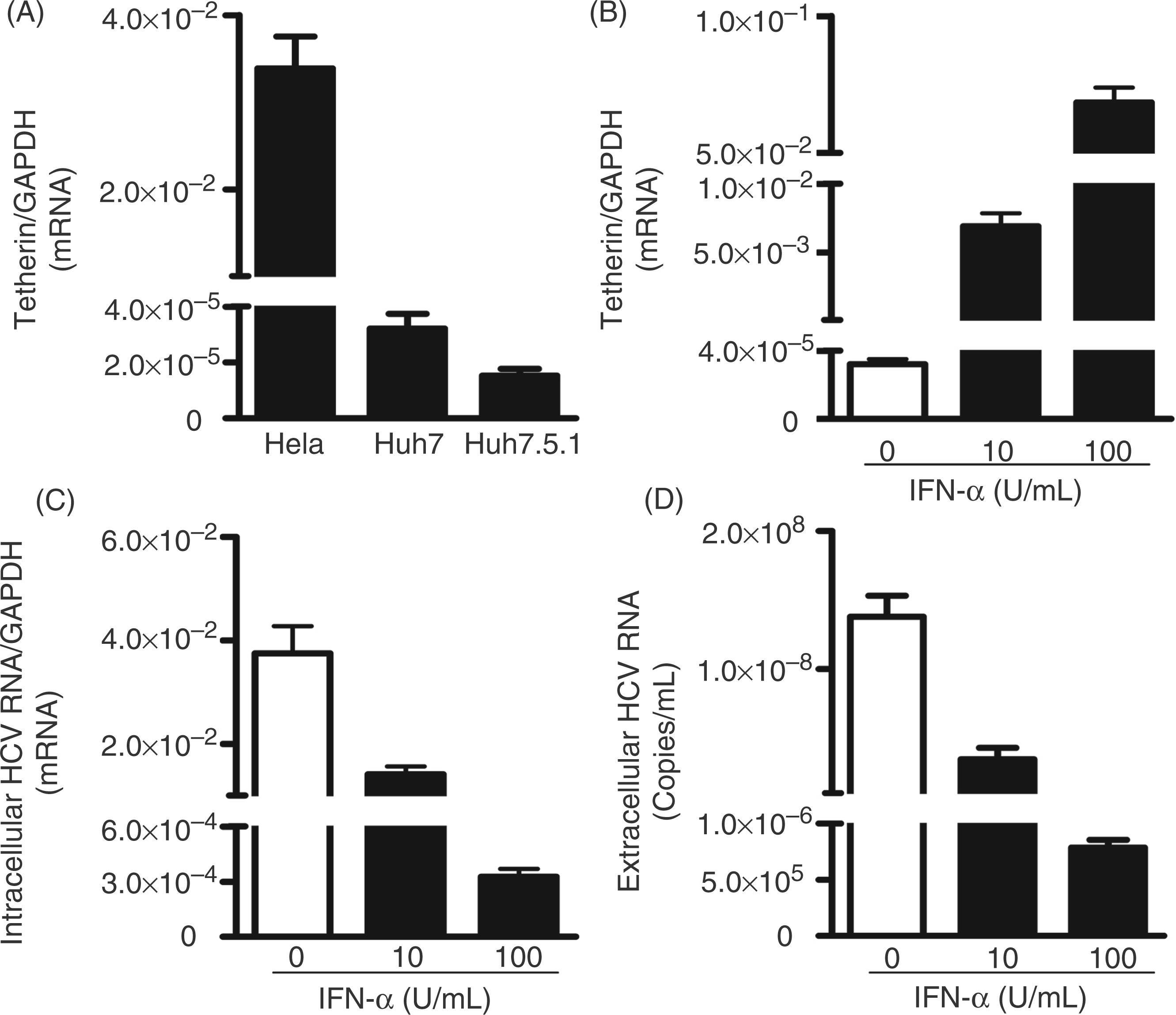
Tetherin inhibits Vpu-deleted HIV-1 release from hepatocytes
As a result of the low expression of tetherin in hepatocytes, we transfected tetherin plasmids (pCR-tetherin-HA) into Huh7 cells. The cells transfected with pCR-tetherin-HA showed positive immunofluorescence staining (Figure 3B). To determine whether exogenous tetherin expression in hepatocytes is functional, we examined the known anti-HIV ability of tetherin. We co-transfected Huh7 cells with plasmids pNL4.3 (full-length HIV-1) plus tetherin-HA, or plasmids pNL4.3-delVpu (Vpu-deleted HIV-1) plus tetherin-HA. As shown in Figure 3, exogenous tetherin expression had little effect on intracellular HIV replication, either for full- length HIV-1 (Figure 3C) or for Vpu-deleted HIV-1 (Figure 3E). In contrast, although exogenous tetherin expression had little effect on full-length HIV-1 release (Figure 3D), its expression significantly inhibited Vpu-deleted HIV-1 release as evidenced by reduced RT activity in culture supernatant (Figure 3F).
Effect of exogenous tetherin on HIV-1 release. (A, B) Exogenous tetherin expression in Huh7 cells. The plasmid encoding tetherin with a HA tag (pCR-tetherin-HA) was transfected to Huh7 cells. At 48 h post-transfection, the exogenous tetherin was shown by immunofluorescence staining with a rabbit anti-HA antibody (magnification:100 ×; scale bars: 100 µm). (B) Cell nuclei were stained with Hoechst 33258. Huh7 cells without plasmid transfection served as control. (C–F) Effect of tetherin on intracellular HIV-1 replication (C, E) and HIV-release (D, F). Huh7 cells were co-transfected with plasmids pNL4.3 (full-length HIV-1) plus tetherin-HA (C, D) or plasmids pNL4.3-delVpu (Vpu-deleted HIV-1) plus tetherin-HA (E, F). Co-transfections of proviral plasmid (pNL4.3 or pNL4.3-delVpu) with vector pCR-HA served as controls. Cells (C, E) and supernatant (D, F) containing released viruses were harvested at 48 h post-transfection. The intracellular (C, E) and extracellular (released; D, F) viral titers were determined by HIV RT activity assay. The results shown in C–F are the mean ± SD of triplicate cultures, representative of three experiments.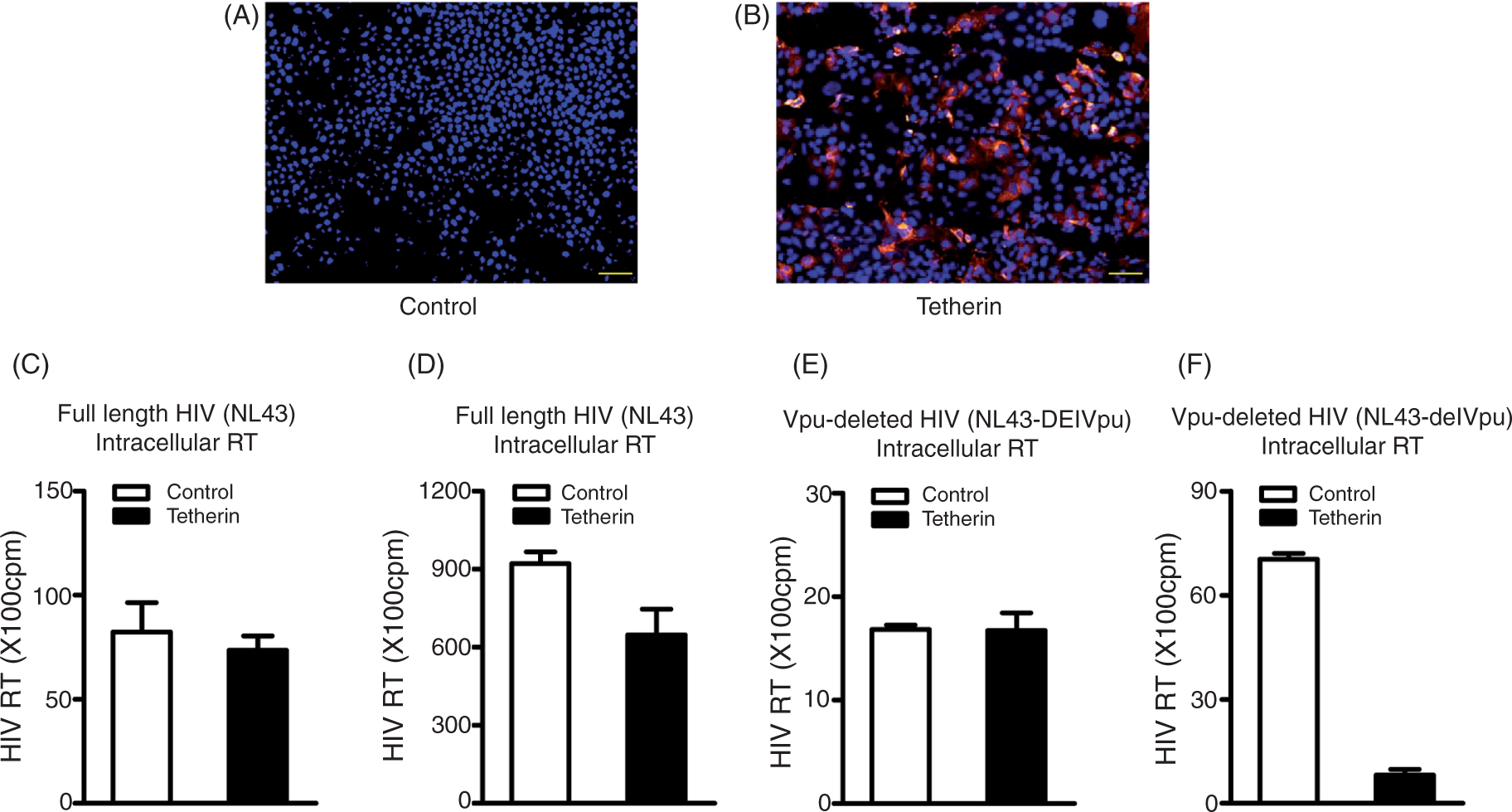
Tetherin has little effect on HCV release from hepatocytes
Having demonstrated the functional expression of exogenous tetherin in hepatocytes, we next examined whether tetherin has the ability to inhibit HCV release from Huh7 cells. The HCV JFH-1-infected Huh7 cells were transfected with pCR-tetherin-HA and then cultured for additional 48 h. Exogenous tetherin expression had little effect on HCV RNA levels, either within cells (Figure 4A) or in the supernatant (Figure 4B). In addition, there was no difference in the production of infectious HCV between the cells transfected with the plasmid expressing tetherin and control cells (Figures 4C, D). The observation that tetherin had little effect on HCV was further confirmed by immunofluorescence assay, showing that the expression of HCV core protein was not affected by tetherin expression in hepatocytes (Figures 5A, B). As tetherin is able to directly retain virions in cells, the colocalization of tetherin and viral proteins is considered as key evidence for the action of tetherin against viruses. Thus, we examined whether there is a colocalization of tetherin and HCV core protein in JFH-1-infected cells. However, the colocalization of tetherin and HCV core protein was not observed in JFH-1-infected Huh7 cells (Figure 5D).
Effect of exogenous tetherin on HCV JFH-1 release. The HCV JFH-1-infected Huh7 cells were transfected with plasmid pCR-tetherin-HA or vector pCR-HA at day 3 post-infection. Cell lysates and culture supernatant were collected at 48 h post-transfection for the HCV RNA analysis and HCV infectivity assay. The levels of intracellular (A) or extracellular (B) HCV RNA were determined by real-time RT-PCR. The levels of intracellular HCV RNA were expressed as relative ratios to GAPDH mRNA. The levels of extracellular HCV RNA were expressed as copies/ml. The titers of infectious virus within cells (C) or in supernatant (D) were determined by serial dilution/immunofluorescence analyses. The levels of intracellular HCV titers, based on their focus forming units/ml (FFU/ml), were expressed as the fold of control (without tetherin transfection, which was defined as 1). The levels of extracellular infectious HCV titers were expressed as FFU/ml. The results shown in A–D are the mean ± SD of triplicate cultures, representative of three experiments. Mutual effect of tetherin and HCV. Uninfected or JFH-1-infected Huh7 cells were transfected with plasmid pCR-tetherin-HA at day 3 post-infection. Tetherin and HCV core protein expression in Huh7 cells was determined at 48 h post-transfection by immunofluoresence staining with antibodies against tetherin or HCV core protein (magnification, A, B, C:100 ×, scale bars: 100 µm, D: 1000 ×, scale bar: 10 µm). The nuclei were stained with Hoechst 33258. (A) HCV core protein expression in the absence of exogenous tetherin; (B) HCV core protein expression in the presence of exogenous tetherin; (C) tetherin expression in the absence of HCV; (D) tetherin expression in the presence of HCV.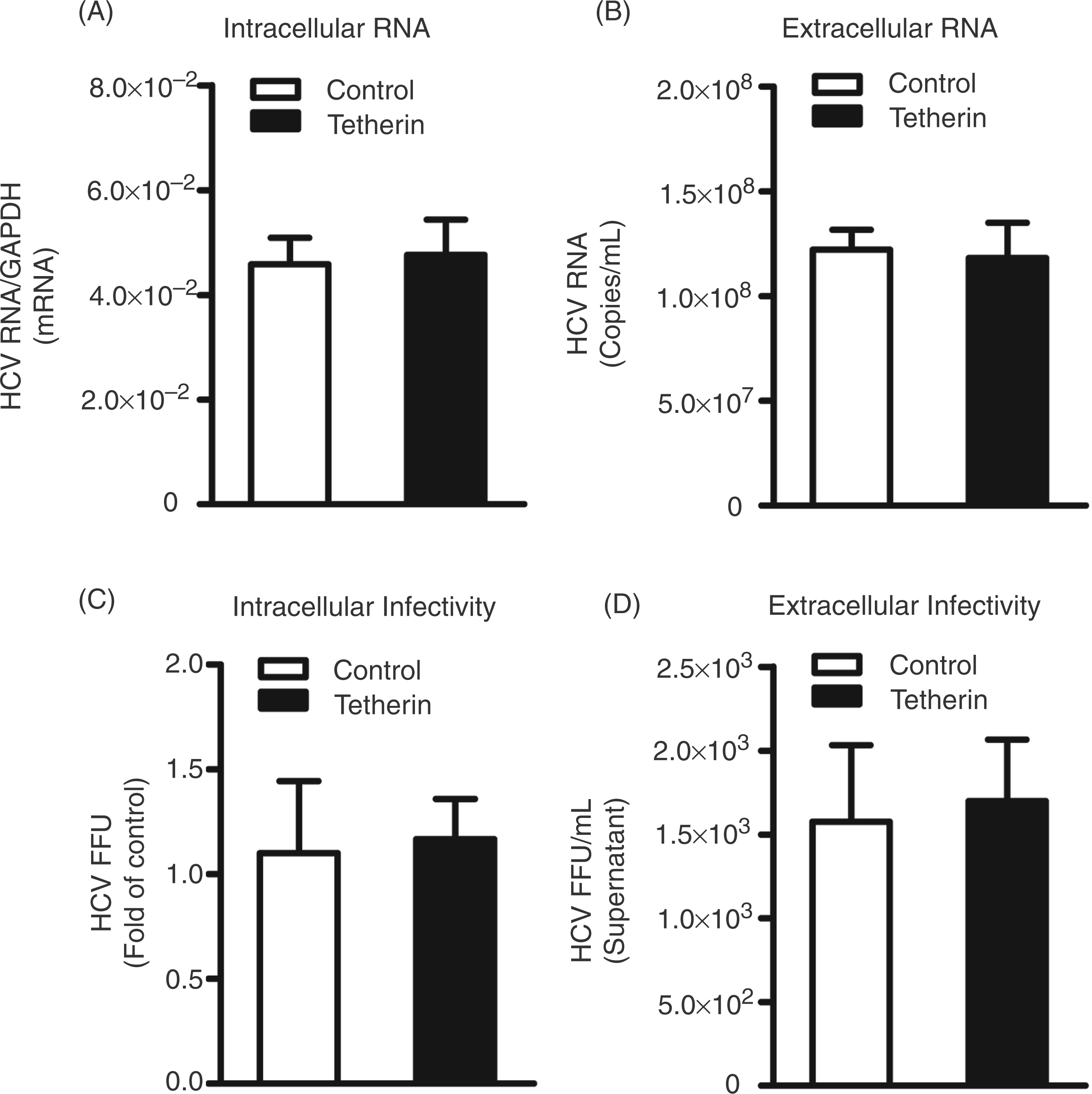
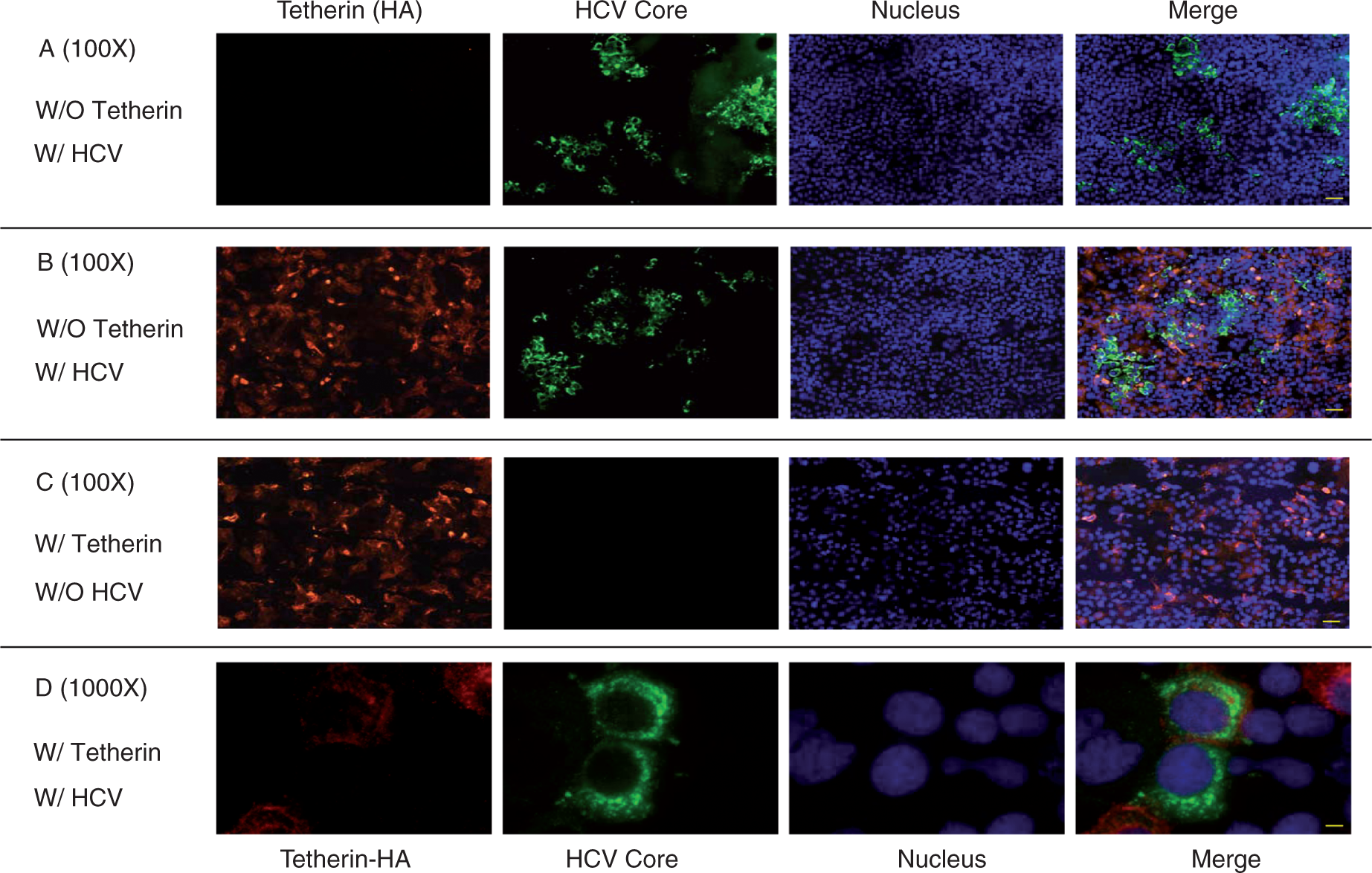
Infection of HCVJFH-1 does not affect the expression and anti-HIV function of tetherin
In addition to the investigation of the impact of tetherin on HCV, we also examined whether HCV infection suppresses the expression and anti-HIV function of tetherin in hepatocytes. As shown in Figure 5, the expression of tetherin in HCV JFH-1-infected cells (Figure 5B) was similar to that in uninfected cells (Figure 5C). Furthermore, there was little difference in HIV RT activity between uninfected and HCV-infected cells co-transfected with tetherin plasmid along with full-length HIV-1 plasmid (Figure 6A) or Vpu-deleted HIV-1 plasmid (Figure 6B).
Effect of HCV infection on tetherin’s action on HIV-1. Uninfected or JFH-1-infected Huh7 cells were co-transfected with plasmids pNL4.3 (Full-length HIV-1) plus tetherin-HA (A) or plasmids pNL4.3-delVpu (Vpu-deleted HIV-1) plus tetherin-HA (B). Co-transfections of proviral plasmid (pNL4.3-delVpu or pNL4.3) with vector pCR-HA served as controls. Supernatant was harvested at 48 h post-transfection and the viral titers were determined by HIV-1 RT assay. The results shown in A and B are the mean ± SD of triplicate cultures, representative of three experiments.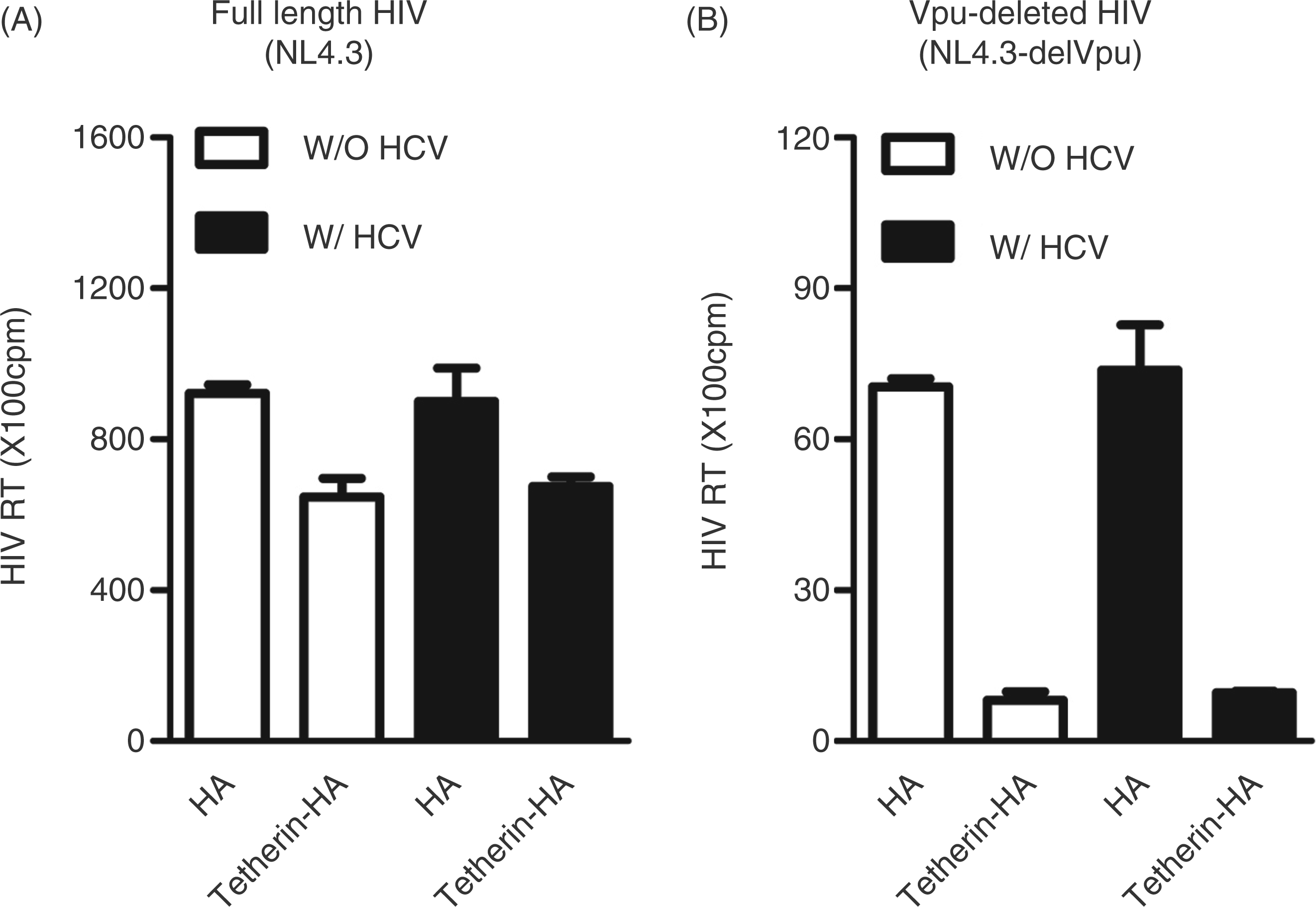
Discussion
Tetherin is an IFN-induced protein that blocks the release of HIV and other enveloped viral particles. As an enveloped RNA virus, HCV possess the structure and biological features that can be targeted by tetherin. Although many human cells do not constitutively express tetherin, its expression can be induced by IFN-α.2,8,11 We showed that human hepatocytes expressed very low levels of endogenous tetherin (Figure 1A), which could be induced by IFN-α treatment (Figure 1B). However, the induction of endogenous tetherin in hepatocytes plays little role in IFN-α-mediated anti-HCV activity (Figure 2). Moreover, the over-expression of exogenous tetherin could not inhibit HCV (Figures 4, 5A, B). In contrast, the expression of tetherin in hepatocytes suppressed Vpu-deleted HIV release (Figure 3F). These data confirmed the reports2,36 that although tetherin had little effect on full-length HIV-1, it potently suppressed Vpu-deleted HIV-1 release (Figure 3F). It is known that HIV-1 normally replicates in T cells or macrophages.37–39 However, a wide variety of many other cell types that are not the target cells of HIV-1, including Hela, HOS and 293T cells, can express endogenous or exogenous tetherins that possess the same virion retention ability.2,40 These findings support our observation that the expression of exogenous tetherin could inhibit Vpu-deleted HIV-1 in human hepatocytes.
The mechanism(s) through which tetherin inhibits the release of viral particles is not clearly understood. As the viruses sensitive to tetherin are all lipid-enveloped, it has been suggested that the lipid envelope may be the vital target of tetherin, which may constitute a simple bridge between host cell and viral lipid bilayers. 40 HCV is a lipid-enveloped virus and replication occurs on a cholesterol-required lipid raft membrane19–21,41 where tetherin is enriched. 14 Our data showed that tetherin had little effect on HCV JFH-1 release, suggesting that the lipid envelope is not absolutely required for the retention action of tetherin. It is known now that the retention activity of tetherin requires colocalization with budding virions.40,42,43 Viruses (such as HIV-1) that can bud from cellular membranes are targeted by tetherin. There is no evidence so far that HCV can bud from cellular membrane, which may explain why tetherin failed to retain HCV in hepatocytes. In addition, this lack of ability to restrict HCV by tetherin could not be related to the counteraction of HCV, as HCV infection of hepatocytes affected neither tetherin expression nor anti-HIV activity of tetherin. These data indicate that the anti-HCV activity of tetherin is negligible in human hepatocytes.
Footnotes
Acknowledgements
We thank Dr Paul D. Bieniasz (The Rockefeller University, New York, NY, USA) for kindly providing plasmids pCR-tetherin-HA, pNL4.3-delVpu and pCR-HA. We also thank Dr Charles Rice (The Rockefeller University, New York, NY, USA) for generously providing Huh7 and Huh7.5.1 cell lines and Dr Takaji Wakita (Tokyo Metropolitan Institute for Neuroscience, Tokyo, Japan) for HCV JFH-1 molecular clone. This work was supported by the National Institutes of Health [grant numbers DA12815, DA22177, and DA27550].
Conflicts of interest
None declared.