
Abstract
Visual crowding is the impairment in recognising an object in the presence of nearby clutter. Two prominent characteristics of crowding are that flankers similar to the target cause more crowding than dissimilar ones, and radially located flankers cause more crowding than tangential ones. These observations are based on studies where a target is surrounded by a small number of flankers. Recent evidence, however, suggests that crowding behaves differently in the presence of a dense set of flankers. Here, we examined the effect of target-flanker similarity and radial–tangential anisotropy in dense crowding in order to probe the mechanisms underlying crowding. Experiment 1 found that the presence of dissimilar flankers reduced crowding in sparse settings, particularly in radial locations, as expected, but paradoxically increased crowding in dense settings. This effect was not attributable to perceived grouping. Experiment 2A manipulated the number of similar flankers adjacent to the target embedded in a dense set of dissimilar flankers and found that just one pair of nearby similar flankers drastically impaired target identification. Experiment 2B manipulated the number of dissimilar flankers adjacent to a target embedded within similar flankers, which revealed a modest improvement in performance with increasing number of dissimilar flankers. There was no evidence of radial–tangential anisotropy in Experiment 2A or 2B. These findings are discussed in relation to prevalent theories of crowding. The results, particularly that of Experiment 2A, lend support to the nearest neighbour hypothesis, which explains crowding as a failure of target segmentation from its immediately adjacent neighbours.
Introduction
Visual crowding is the impairment of object recognition in the presence of clutter. Extensive research into this phenomenon has identified a set of characteristics associated with crowding. Of primary interest to this study are the powerful effects of (a) similarity and (b) spatial anisotropy. Specifically, flankers similar to the target cause more crowding than do dissimilar flankers (Chakravarthi & Cavanagh, 2007; Kennedy & Whitaker, 2010; Kooi et al., 1994). Further, flankers located along the line connecting the target to the current fixation (the radial axis) cause more crowding than flankers located along the line perpendicular to the radial axis (the tangential axis), usually termed radial–tangential anisotropy (Greenwood et al., 2017; Kurzawski et al., 2023; Toet & Levi, 1992).
These well-established characteristics are generally observed in typical tests of crowding, where participants are asked to identify a peripheral target surrounded by a small number of flankers. However, the presence of a large number of flankers (so-called ‘dense’ setting), which might be more common in everyday life, seems to lead to outcomes not expected from our observations of crowding in sparser (classic) settings. For example, in an early study, Poder (2006) reported that while a small number (8) of red oriented rectangles surrounding a blue target rectangle impaired the latter’s orientation discrimination, crowding became weaker in the presence of a much larger number (80) of red rectangles, countering the expectation that more flankers around the target should lead to more crowding.
More recently, Van der Burg et al. (2017) deployed a genetic algorithm to determine the configuration of flankers that would lead to the least crowding in a dense display made up of 1 near-vertical target surrounded by 284 closely packed vertical and horizontal flankers. Their results showed that the similarity of only the nearest neighbours mattered: crowding was lowest when the tangentially placed flankers were dissimilar to the target. These results, once again, contradict our expectations that more dissimilar flankers should weaken crowding (Manassi et al., 2012; Rosen & Pelli, 2015). This was reinforced by another study by the same researchers (Van der Burg et al., 2024) where they found that the similarity of the flankers immediately adjacent to the target in a dense display determined target identification performance. Specifically, when all eight flankers adjoining the target were similar or dissimilar to the target, identification performance was the lowest or highest, respectively; the orientation of the remaining 250+ flankers did not alter performance much. Further, if the orientations of the eight immediately adjoining flankers were allowed to randomly vary, target identification varied as a function of the proportion of (dis)similar flankers. Thus, it appears that the similarity of only the nearest flankers matters in dense settings.
One intriguing finding in their studies was that, contrary to expectations of the radial–tangential anisotropy, similarity of the tangential flankers was more influential than that of the radial flankers (Van der Burg et al., 2017). This, once again, demonstrates that outcomes in dense settings are not straightforwardly predictable from those in the well-studied sparse settings. One reason that the tangential flankers were more influential might have been because of target-flanker grouping. Bornet et al. (2021) tested several computational models to examine if they could account for crowding in dense displays. Models that allowed some sort of segmentation and grouping among the objects were more successful at reproducing human behaviour. For example, in one model (LAMINART), the tangential flankers could group with the target if they were similar (i.e., vertically aligned) but not if some of them were dissimilar (e.g., horizontally oriented). Hence, dissimilar tangential flankers led to less crowding by preventing grouping between target and flankers. Such grouping by alignment was not possible with radial flankers. That is, the inverted radial–tangential anisotropy might be due to the usage of vertical targets, which can align only with other vertical lines placed in tangential locations but not in radial locations. A different target (say a horizontal line) might result in the standard radial–tangential anisotropy in such settings.
Studies examining crowding in dense contexts indicate that the traditional explanations of crowding, such as feature pooling (Balas et al., 2009; Freeman et al., 2012; Greenwood et al., 2009; Levi, 2008; Parkes et al., 2001; Pelli et al., 2004; van den Berg et al., 2010), substitution (Ester et al., 2014; Hanus & Vul, 2013; Strasburger & Malania, 2013), or limitations in attentional resolution (Chakravarthi & Cavanagh, 2007; He et al., 1996; Intriligator & Cavanagh, 2001), are either insufficient to explain such results or need to be modified. Other proposals that incorporate either local (Van der Burg et al., 2017, 2024) or global (Herzog et al., 2015; Manassi et al., 2012, 2013) grouping of elements in the display might be more successful at explaining crowding in both dense and sparse settings. Thus, examining whether the typical characteristics observed in sparse settings are also present in dense crowding might shed light on the overall mechanisms underlying this ubiquitous phenomenon.
In this study, we systematically examined the effect of target-flanker similarity on dense crowding, particularly in relation to radial–tangential anisotropy. We tested this question with multiple types of stimuli and organisations of flankers, while also using a different manipulation of similarity (contrast polarity instead of alignment/orientation) compared to recent studies on dense crowding, to assess the generality of the findings in dense crowding. These would also allow us to probe the mechanisms of crowding in both dense and sparse settings.
Experiment 1
In this experiment, we compared the effect of the similarity of flankers nearest to the target across sparse and dense settings. The nearest flankers could be either all similar, or a subset of them could be dissimilar. This allowed us to also manipulate the location of the dissimilar flankers: they could be either along the radial axis or the tangential axis. Findings from sparse settings predict that the presence of dissimilar flankers should lead to less crowding, and this alleviation should be stronger with radial dissimilar flankers than with tangential dissimilar flankers. However, this pattern might not hold in dense settings (Van der Burg et al., 2017). Further, in addition to these manipulations, we also examined the effect of the similarity of distant flankers, which would complement the tests conducted by Van der Burg et al. (2024). If the similarity of only the nearest flankers is relevant, then the similarity of the distant flankers should not affect target identification.
Method
Participants
Twenty participants (13 female, 7 male) aged (mean ± SD) 22.4 ± 4.1 years, were tested. Participants self-reported normal or corrected to normal vision. All participants gave written, informed consent. The study was approved by the Psychology Ethics Committee at the University of Aberdeen.
Materials and Stimuli
The experiment was conducted on a HP EliteDesk 800 G4 computer running Ubuntu 20.04 LTS operating system controlling a 19-inch Sony Trinitron GDM-F520 CRT (Sony Corporation, Tokyo, Japan) display, with a refresh rate of 100 Hz and a resolution of 1,024 × 728 pixels. Viewing distance was set to 57 cm with head position secured by a chinrest. This setup results in ~38 pixels per degree of visual angle. Stimuli were generated and presented using MATLAB with Psychophysics Toolbox extensions (Brainard, 1997; Kleiner et al., 2007; Pelli, 1997).
Stimuli consisted of a target T and ‘squared theta’ flankers, each with a width and height of 0.5°. Previous studies have successfully used such squared theta flankers to crowd orientation discrimination of target T’s (Chakravarthi & Bertamini, 2020; Feng et al., 2007; Tripathy & Cavanagh, 2002; Tripathy et al., 2014). The target was presented at an eccentricity of 6° and could take one of four orientations, whereas the flankers could be in one of two orientations. These characters were either black (luminance = 0.25 cd/m2) or white (91.5 cd/m2) presented on a mid-grey background (45.9 cd/m2). A yellow fixation cross (78.3 cd/m2), with lines of length 0.4°, was presented throughout each block.
This experiment compared performance in dense displays to that in sparse displays. In the sparse conditions (Figure 1, top row), a target was presented along with four flankers – above, below, left, and right of the target, with a centre-to-centre spacing of 1°. In the sparse-rad condition, the radial flankers (left and right) were of the opposite polarity as the target, and the other two flankers (above and below) had the same polarity as the target; in the sparse-tan condition, only the tangential flankers had the opposite polarity; in the sparse-both, all four flankers had the opposite polarity, and in the sparse-none, all flankers had the same polarity. In the dense conditions (Figure 1, middle row), the target was surrounded by 5 rings, consisting of 124 flankers. The rings were spaced 1° apart, with the nearest ring 1° from the target. Adjacent flankers within each ring were 0.75° apart. 1 In the dense-rad condition, only the nearest two radial flankers had the opposite polarity as the target; in the dense-tan, the nearest two tangential flankers had the opposite polarity; in the dense-both, the nearest radial and tangential flankers had the opposite polarity, in the dense-none, all flankers had the same polarity as the target. We also included three conditions (Figure 1, bottom row) with the same local configuration as the dense conditions (excluding the dense-none), but the polarity of each flanker in the outer four rings was chosen randomly to be positive (white) or negative (black). These three mixed-polarity dense conditions test if the similarity of only the immediately adjacent flankers in dense displays determines crowding. Finally, we also included an unflanked condition as a baseline.

Illustration of stimulus configurations. Top row: sparse displays; middle row: dense displays with uniform-polarity outer rings; bottom row: dense displays with mixed-polarity outer rings. Columns depict manipulation of dissimilarity among flankers immediately adjacent to the target: either all flankers had the same polarity as the target (first column) or a pair of tangential (second), radial (third), or both (fourth) flankers had the opposite polarity as the target. The stimuli were presented randomly to the left or right of fixation. The target T was black in one half of the experiment and white in the other half, counterbalanced across participants.
Procedure
The experiment started with a practice block, which acquainted the participants with the task as well as with the various conditions to be tested. Each trial started with a fixation screen for 700 ms during which a yellow fixation cross along with two green circles of diameter 0.1° at the potential target locations (left and right of the target) was presented. The stimulus was then presented for 150 ms on either the right or the left side of fixation. The participants were asked to report the orientation of the target T using the arrow keys. The next trial started immediately after response.
Each of the 12 conditions described above was tested 7 times in a block, and the order of presentation of the conditions was randomised within the block. The experiment included a total of 12 blocks, and hence each condition was tested with 84 trials over the course of the experiment (a total of 1,008 trials in 12 blocks of 84 trials each). Target polarity was kept constant in the first half of the experiment and changed to its opposite in the second half. The order of target polarity was counterbalanced across participants.
Perceived Grouping Ratings
Grouping has been argued to play a significant role in crowding (Herzog et al., 2015; Manassi et al., 2012, 2013) and also forms part of the explanation of crowding in dense settings (Bornet et al., 2021; Van der Burg et al., 2024). Hence, we sought to test if grouping might explain the pattern of results in this experiment. A new set of nine psychophysically experienced observers (seven female, age: 34.3 ± 5 years) were asked to rate the strength of grouping between the target T and the surrounding flankers in each condition. The experiment was conducted online through the Inquisit platform. The stimulus settings were comparable to that in the main experiment: stimulus presentation duration was 150 ms; the presentation order of conditions was randomised; target (and its flankers’) location on each trial was randomly selected between the left and right of a fixation cross; participants were instructed to place the screen at their typical viewing distance (approximately 40 cm) and fixate on the central cross, which stayed on throughout the study. Since viewing distance and monitor properties such as resolution could not be fixed, the exact eccentricity, size, and spacing of stimuli might vary across participants. However, since these factors (eccentricity, spacing and size of objects) scale proportionally with the relevant parameters (distance to screen, screen resolution), the stimuli are comparable, and the findings should be generalisable to the main experiment. Following Manassi et al. (2012), we asked participants to report on a 0 to 10 scale ‘How much does the target T stand out from among the flanker boxes?’, with 0 being ‘Does not stand out’ and 10 being ‘Strongly stands out’. We then subtracted these scores from 10 to obtain perceived grouping scores, where 0 would indicate no grouping and 10 strong grouping between the target and the flankers. We then averaged the ratings across four presentations for each condition.
Results
Data and analysis code for all experiments are available at OSF: https://osf.io/3gpf4/.
Participants’ accuracy in reporting the target orientation in the absence of flankers was high (mean ± SEM = 0.96 ± 0.01). Note that chance performance is 0.25. Further, participants’ average performance across all conditions was relatively high even in the presence of flankers: both in sparse (0.76 ± 0.02) and, surprisingly also, in dense conditions (0.65 ± 0.03). The latter could be because, unlike some previous experiments (Van der Burg et al., 2017, 2024), the target was easy to locate, as it was at the centre of the rings, and positional pre-cues were presented for a considerable duration (700 ms before stimuli were presented).
Omnibus Analysis
We conducted a two-way repeated measures ANOVA on proportion correct in target identification with context (sparse or dense) and dissimilar flanker configuration (none, tangential, radial, both) as factors. As expected, identifying the target was easier in sparse than dense displays (F[1,19] = 44.1, p < .001,
Sparse Displays
Accuracy was affected by the configuration of dissimilar flankers (F[3,57] = 11.71, p < .001,
Dense Displays
Performance in dense-uniform displays paints a different picture. Identification of a target was affected by the configuration of dissimilar flankers (F[3,57] = 6.87, p < .001,
Effect of Similarity of Distant Flankers
We also included conditions where the polarity of each element in the outer four flanker rings was randomly assigned. To assess the role of non-adjacent flankers on crowding, we conducted a two-way repeated measures ANOVA with polarity of the outer four flanker rings (uniform or mixed) and the configuration of adjacent dissimilar flankers (tangential, radial, both) as factors. There was an almost significant effect of outer-flanker polarity (F[1,19] = 4.16, p = .056,
Perceived Grouping
Figure 2B shows that the target is strongly grouped with the flankers in the dense-mixed conditions. Target-flanker grouping is much weaker in the dense-uniform condition and even less so in the sparse conditions. One might expect that stronger grouping leads to worse performance. However, this pattern was not observed: identification performance was comparable to or even better in the dense-mixed conditions than in the dense-uniform conditions. Further, surprisingly, in the dense-uniform and sparse conditions, grouping seems to be stronger when the nearby four flankers were dissimilar, but identification was either better or not worse in this configuration relative to other configurations. Thus, the observed pattern of target identification results cannot be fully or easily attributable to overall perceived grouping of the target with the flankers.
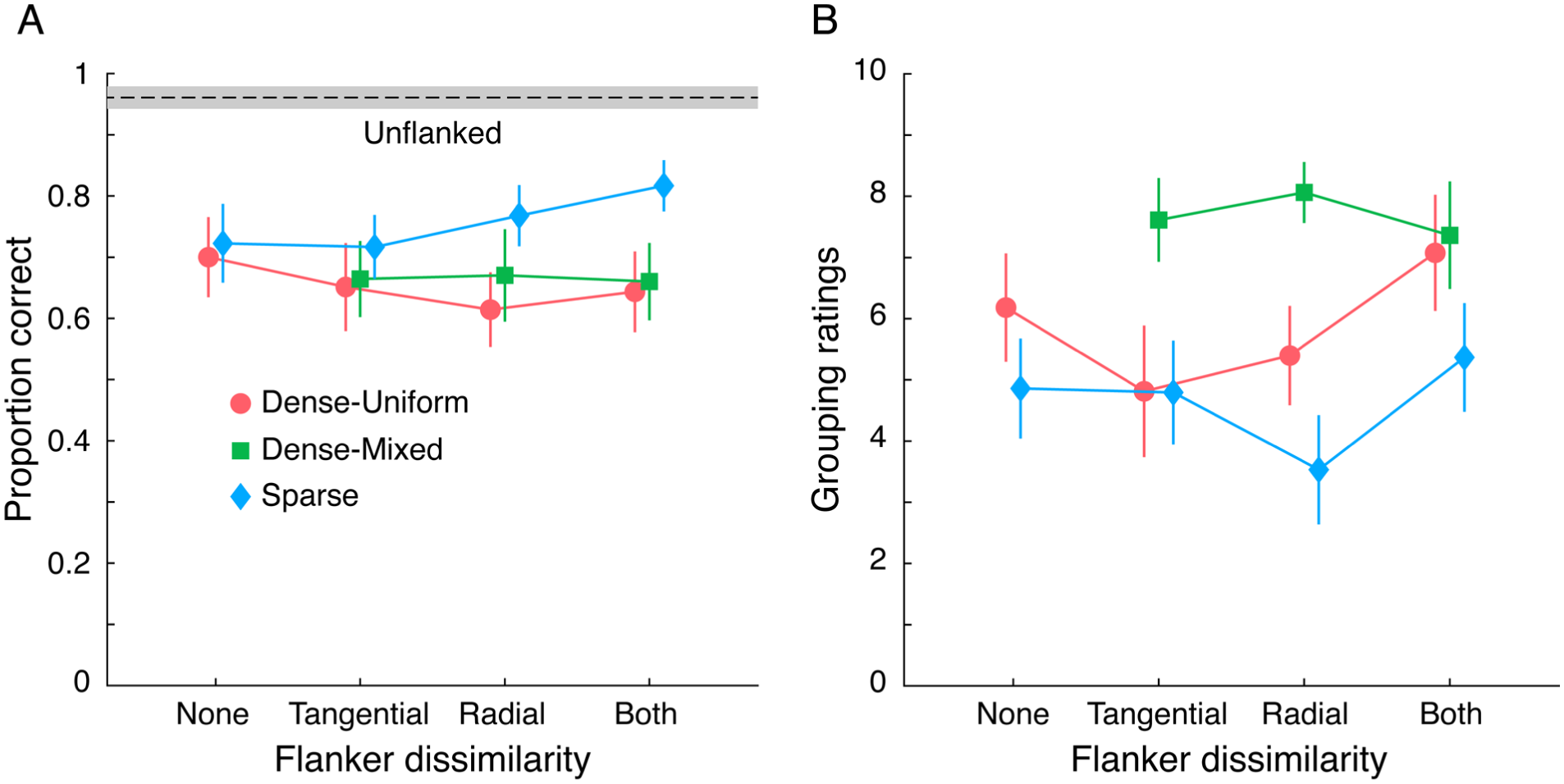
Results of Experiment 1. (A) Crowding: Participants’ accuracy in identifying the orientation of a target T plotted for each of the four dissimilar flanker configurations in each of the three contexts. The dashed line represents accuracy in the absence of flankers. (B) Grouping: Ratings of target grouping with the flankers on a 0 to 10 (0 = not grouped; 10 = strongly grouped) scale in the same conditions as in (A). In both panels, shaded area and error bars represent between-subjects 95% confidence interval.
In summary, the pattern of results, as a function of the similarity of adjacent flankers, was distinct in sparse and dense settings. While the outcomes were in line with expectations in sparse settings, in dense settings we found an unexpected worsening of performance when two flankers adjacent to the target were dissimilar. These results were hard to reconcile with perceived grouping of the target with the flankers.
Experiment 2
Experiment 1 demonstrated that the similarity of nearby flankers affects target identification differently in sparse and dense contexts, and that this cannot be easily attributable to grouping. However, identification performance, while considerably less in the crowded conditions than in the unflanked condition, was not much different between sparse and dense conditions. One would have expected a much larger difference between these conditions, especially since performance was not close to chance performance. In the dense conditions, the flankers were probably bound to each other more strongly than to the target (visible as distinct rings), thus reducing crowding and potential effects of target-flanker dissimilarity. Further, one might worry that the range of performance was too narrow to allow strong inference.
Experiment 2 implemented a design, inspired by that used successfully by Van der Burg et al. (2017, 2024), to induce strong crowding in dense contexts. We also systematically varied the number of similar or dissimilar flankers. Finally, the flanker stimuli were changed to more closely resemble the target and share some of the response characteristics of the target, thus further enhancing crowding (Reuther & Chakravarthi, 2020). These manipulations should together increase the strength of crowding as well as the range of performance across conditions, thus helping shed light on the mechanisms of crowding in dense contexts.
Method
Participants
Experiment 2A and 2B tested 24 participants each (2A: 17 female, 7 male, aged 24.4 ± 6.4 years; 2B: 6 female, 18 male, aged 20.4 ± 1 years). Participants self-reported normal or corrected to normal vision. All participants gave written, informed consent. The experiments were approved by the Psychology Ethics Committee at the University of Aberdeen.
Materials and Stimuli
The materials were the same as in Experiment 1. The target was an oriented T of size 0.6° presented at an eccentricity of 7°. Flankers were letters C, E, F, H, I, and L constructed of vertical and horizontal lines to share features with the target (Figure 3), also of size 0.6°. All objects were placed within a 13 × 13 grid (168 flankers with the target T in the centremost cell), with an inter-cell spacing of 1°. The letter in each cell was selected randomly and could take one of four orientations, thus sharing the same response affordances as the target. Each experiment had three ‘baseline’ comparison conditions: (a) a no flanker unflanked condition, which should indicate the highest performance that can be obtained, (b) all flankers shared the same polarity as the target (‘all-flankers-similar’), which might give the lower bound of performance using this specific stimulus (but see Experiment 1) and (c) all flankers had the opposite polarity as the target (‘all-flankers-dissimilar’), which should give the higher bound performance in dense contexts within this setup. In addition, each experiment manipulated target-flanker similarity in distinct ways.

Illustration of stimuli used in Experiment 2. A target T, in one of four orientations, was presented at the centre of a 13 × 13 grid of rotated letters. The fixation cross is shown at its approximate location. In the actual experiment, the fixation was yellow and presented on the same grey background as the letters. The number of nearby flankers that were similar (top row, Experiment 2A) or dissimilar (bottom row, Experiment 2B) to the target in terms of contrast polarity was manipulated from one to six pairs in either the radial or tangential direction.
Experiment 2A
All flankers were dissimilar to the target, and the number of similar flankers was systematically increased. In the Radial/Tangential 1 pair condition, the immediately adjacent two flankers to the target (one on either side, along the radial or tangential axis, respectively) were similar in polarity to the target; in the Radial/Tangential 2 pairs condition, the immediately adjacent four flankers to the target (two on each side) shared polarity, and so on until the Radial/Tangential 6 pairs conditions (the full row/column).
Experiment 2B
The similarity manipulation was flipped compared to 2A. All flankers were similar in polarity to the target, and the number of flankers dissimilar to the target was increased. The immediately adjacent two flankers to the target (one on either side) were dissimilar in polarity to the target; the immediately adjacent four flankers to the target (two on each side) had the opposite polarity, and so on. As in Experiment 2A, dissimilarity was manipulated either along the radial (horizontal) row or the tangential (vertical) column.
Procedure
The procedure was the same as in Experiment 1, except for the following changes. Experiments 2A and 2B included 15 conditions (no flanker, all-flankers-similar, all-flankers-dissimilar; 1 to 6 pairs of dis(similar) tangential flankers, 1 to 6 pairs of dis(similar) radial flankers). Each condition was tested in 60 trials (total of 900 trials) spread over 12 blocks. As in Experiment 1, target polarity was constant in the first half of the experiment and changed to its opposite in the second half. The order of target polarity was counterbalanced across participants.
Results
Experiment 2A
One participant’s accuracy data were recomputed by flipping their orientation responses by 180°, as they had used the opposite keys for each response. One participant’s data were excluded from analysis as their performance in the unflanked condition was low (0.55). To briefly preview the results (Figure 4), the presence of just one (or two) pair(s) of similar flankers adjacent to the target dramatically reduced performance to as much as when all 168 flankers were similar.
To examine the effect of flanker similarity along the radial and tangential axes in dense settings, we conducted a 2 × 6 repeated measures ANOVA on accuracy data with factors: flanker locations (radial, tangential) and number of similar flanker pairs (one to six pairs). The effect of flanker locations was almost significant (F[1,22] = 3.56, p = .073,
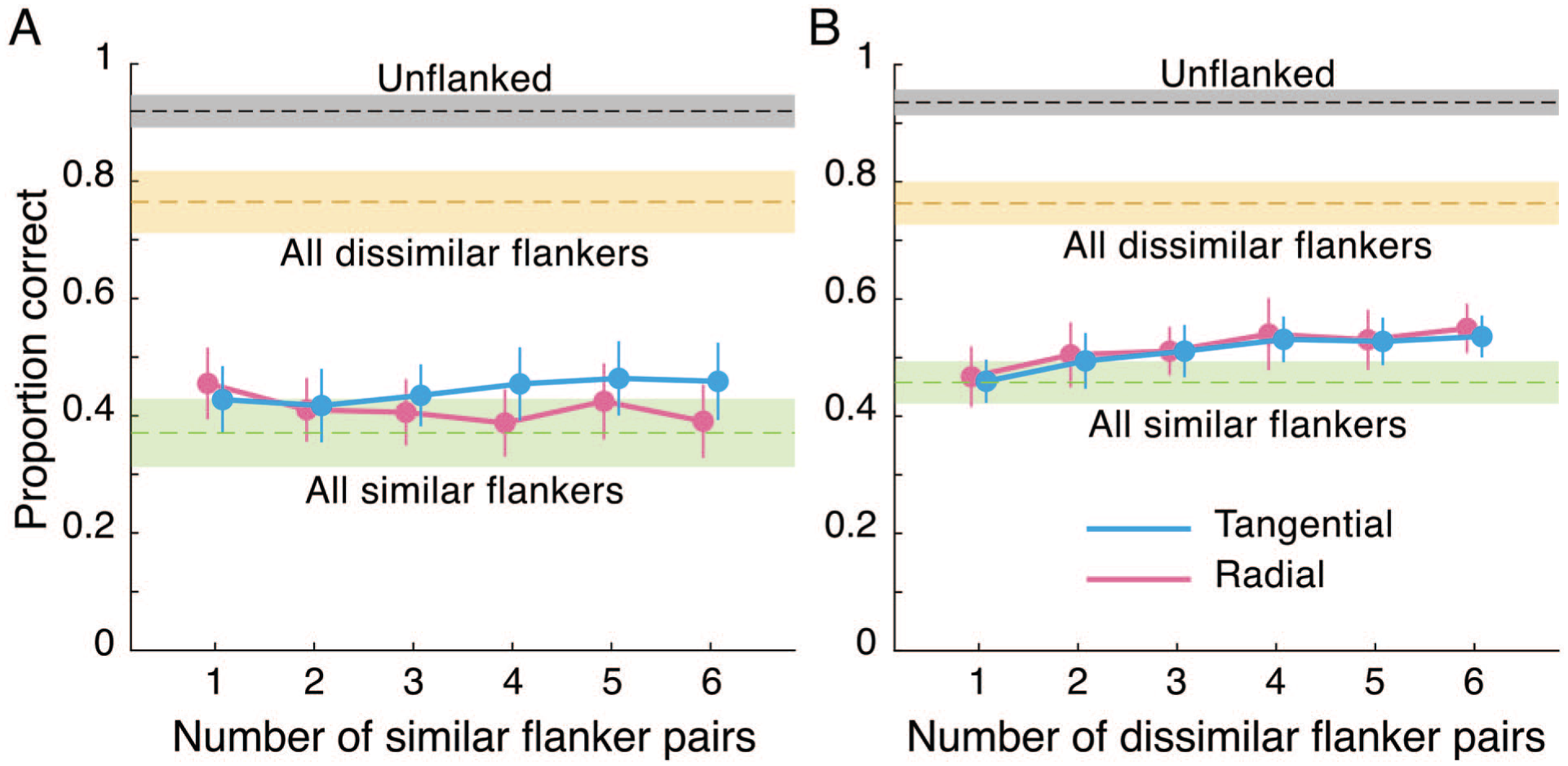
Results from Experiment 2A (left) and 2B (right). Performance is plotted as a function of the number of adjacent flanker pairs that were similar (A) or dissimilar (B) to the target along the radial (pink) or the tangential (blue) axis. Performance when no flankers were presented is indicated by a dashed black line. Performance when all flankers were similar (dashed green line) or were dissimilar (dashed orange line) is also depicted. Error bars and shaded areas represent between-subjects 95% confidence intervals.
We also examined the strength of crowding when only the nearest neighbours were similar to the target. To do so, we compared accuracy (Bonferroni–Holm corrected) in each of the 12 conditions with increasing number of similar flankers to the all-flankers-similar condition (where all flankers were similar to the target). Along the radial direction, performance was better than the all-flankers-similar condition when one (t[22] = 3.8, p = .012) pair of flankers was similar to the target but did not differ with two or more pairs of similar flankers (p’s >.07). Conversely, along the tangential direction, performance did not differ from the all-flankers-similar condition when there were one or two pairs of similar flankers (p’s >.15), but was better when there were three (t[22] = 3.19, p = .032), four (t[22] = 3.48, p = .022), five (t[22] = 3.6, p = .022) or six (t[22] = 3.47, p = .02) pairs of similar flankers.
In summary, the setup used in this experiment led to strong crowding. While there are some differences between the effects of flankers in radial and tangential locations, the main take away is that the presence of just a few similar flankers in the vicinity of the target severely and adversely affects its identification. This effect is so strong that one to two such pairs in tangential locations or three or more pairs in the radial locations are sufficient to reduce performance to that when all flankers are similar to the target.
Experiment 2B
One participant’s accuracy data were recomputed by flipping their orientation responses by 180°, as they had used the opposite keys for each response. Accuracy data were analysed with a similar 2 × 6 repeated measures ANOVA with factors flanker location and number of dissimilar flanker pairs. There was no difference between the effects of radial and tangential dissimilar flankers (F[1,23] = 0.58, p = .454,
We then determined the number of dissimilar flankers needed to overcome the strong crowding induced by the dense set of similar flankers in the grid. Pairwise comparisons (Bonferroni–Holm corrected) indicated that performance in the presence of one or two pairs of adjacent dissimilar flankers did not improve performance relative to when all flankers were similar to the target (all p’s >.12), but further increases in flanker dissimilarity improved performance (radial flankers: three pairs: t[23] = 3.8, p = .008; four pairs: t[23] = 3.65, p = .008; five pairs: t[23] = 3.61, p = .007; six pairs: t[23] = 4.85, p < .001; tangential flankers: three pairs: t[23] = 3.08, p = .025; four pairs: t[23] = 4.3, p = .003; five pairs: t[23] = 3.77, p = .008; six pairs: t[23] = 4.98, p < .001).
Overall, one or two pairs of dissimilar nearest neighbours in a grid of similar flankers does not appear to relieve crowding. This, interestingly, was the case for both radial and tangential dissimilar flankers. With more nearby dissimilar flankers, some relief was observed, although this relief was modest. This could be due to grouping among flankers, often invoked to explain incidences of uncrowding, or could be due to a reduction in the interference from nearby flankers. In either case, the large number of remaining (similar) flankers might still interfere with the target (the results of Experiment 2A indicate that the presence of any nearby similar flankers can lead to strong crowding), and hence crowding is not fully mitigated.
Experiments 2A and 2B were both successful in inducing strong crowding in the presence of dense flankers. However, our manipulations of similarity among nearby flankers did not modulate performance substantially. Only when all flankers were dissimilar was performance high.
Discussion
This study examined the effect of target-flanker similarity on crowding in dense settings, in relation to the radial–tangential anisotropy. Experiment 1 found that radially and tangentially located dissimilar flankers affect target identification differently in sparse and in dense settings. This pattern of results could not be explained by the strength of perceived grouping between the target and its flankers. A primary finding in this study was that the presence of a mere one or two pairs of similar flankers adjacent to the target, when all other flankers were dissimilar (Experiment 2A), was sufficient to cause dramatically strong crowding, to an extent comparable to when all flankers were similar. Conversely, a small number of dissimilar flankers near the target in a sea of similar flankers could not alleviate crowding (Experiment 2B). Further, the location of the (dis)similar flankers (radial or tangential) did not make much of a difference.
There are relatively few studies that manipulated flanker dissimilarity in dense settings. An earlier study in our lab (Chakravarthi & Bertamini, 2020) found no effect of polarity (mixed vs. similar) for crowding in dense displays (Experiment 2A, 2B of the previous study), while observing a clear modulation in performance due to similarity in sparse conditions. The results of Experiments 1 and 2B of the current study are in line with these earlier findings.
Across the current set of experiments, the presence of a dense field of similar flankers led to strong crowding (~30 percentage points, or pp, reduction in accuracy relative to the no flanker condition in Experiment 1 and ~50 pp in Experiment 2). Further, when all flankers were dissimilar to the target, crowding was substantially alleviated, relative to when all flankers were similar (~30–35 pp improvement in Experiments 2A and 2B), although crowding was not eliminated. This is in line with previous results (Manassi et al., 2012; Poder, 2006). However, our manipulations of flanker similarity and their location did not alter performance to a large extent. Specifically, there was little to no effect of similarity of radial versus tangential flankers in dense settings, although their locations did matter, in the expected way, in sparse settings. Further, while there was some improvement in performance due to the increasing number of dissimilar flankers (Experiment 2B), performance remained close to that observed when all flankers were similar to the target.
Interestingly, the presence of a pair (or more) of adjacent opposite polarity flankers did not provide any location cueing advantage for target identification. A portion of crowding has been attributed to an inability to locate the target (e.g., Ester et al., 2014; Huckauf & Heller, 2002; Strasburger & Malania, 2013). The pair of dissimilar flankers should have reduced this location uncertainty and thus improved performance (Keshvari & Rosenholtz, 2016; but see Doerig et al., 2019). The lack of such an effect could be due to the strong crowding from all the nearby similar flankers despite a reduction in location uncertainty. A more likely reason might be because the target locations were already cued on each trial; the possible location of the targets was indicated by green dots lasting for at least 700 ms before stimulus onset. Hence, any advantage due to the reduction of target location uncertainty might already have taken place.
Another noteworthy point is that a subset of conditions in Experiment 2B were conceptual replications of a subset of conditions in Experiment 1. That is, the dense-none, dense-rad, and dense-tan conditions (of the uniform type) in Experiment 1 were comparable to the all-similar, Radial 1 pair, and Tangential 1 pair conditions, respectively, in Experiment 2B. However, the results of Experiment 1 were not replicated, with differences among these conditions being observed in Experiment 1 but not in Experiment 2B. This discrepancy might be attributable to differences in stimulus and organisational differences between the experiments (see below).
Radial–Tangential Anisotropy
The location of (dis)similar flankers (radial vs. tangential) made little to no difference in outcomes in dense settings. There were mild effects in Experiments 1 and 2A, and no differences in Experiment 2B. In Experiment 1, radially placed dissimilar flankers caused slightly more crowding than tangential dissimilar flankers. In Experiment 2A, performance was slightly worse with radial similar flankers than tangential ones. In both cases, the radial–tangential difference, whatever its sign, was weak. This lack of a difference might appear hard to reconcile within theories that posit a fixed oval zone of interference (standard pooling and attentional theories). Specifically, an increasing number of similar flankers should lead to more crowding along the radial direction than the tangential direction. The absence of this effect in Experiment 2A might be because performance with a single pair of similar flankers was already close to that of all-flankers-similar condition. Although it is not the theoretical floor (chance levels), it was perhaps the lowest performance that one might expect in these settings; that is, there might have been an effective range compression. Further, there were hints of a difference between the flanker locations – increasing number of similar flankers did not change performance along the tangential direction, whereas performance decreased (even if within a small range) when such flankers were located along the radial direction (significant linear contrast for the decreasing performance). That is, it could be the lack of range in performance that could not reveal potentially existing differences; the strong effect of dense crowding effectively might have buried any existing anisotropy.
Similarly, in Experiment 2B, one might expect an effect of flanker location, where the effect of increasing number of dissimilar flankers should taper off earlier along the tangential direction than in the radial, if the zone of interference is limited to an oval around the target. But the effect of dissimilarity was weak enough that we might not be able to see such a difference if it did exist. Overall, the weak radial–tangential anisotropy is potentially a larger issue for pooling theories than for grouping and nearest neighbour theories, which do not posit an oval interference zone, but further studies are needed to test this possibility. Even though this finding mildly favours the grouping and the nearest neighbour theories, these latter proposals must nevertheless be able to account for the well-established radial–tangential anisotropy in sparse contexts. Currently, they do not provide explicit mechanisms that incorporate the anisotropy.
Role of Flanker Configuration
Experiments 1 and 2 used different flanker configurations in inducing dense crowding. Did these differences affect outcomes? First, we note that outcomes for the same conditions were comparable across Experiments 2A and 2B (unflanked, all-similar-flankers, and all-dissimilar-flankers). Further, both Experiments 2A and 2B produced stronger overall crowding than Experiment 1 (~50 pp vs. ~30 pp when all flankers were similar to the target). Experiment 1 used flankers arranged in concentric rings which could produce perceptual grouping among flankers to form rings while in Experiment 2 flankers were organised in a grid. Hence, there might have been stronger target-flanker grouping in Experiment 2 than in Experiment 1, leading to stronger crowding in Experiment 2. Part of this stronger crowding might also be attributed to the combined influence of greater eccentricity (7° vs. 6°), more flankers (168 vs. 124), and the use of letter flankers that shared response-relevant features with the target in Experiment 2. Unsurprisingly, such stimulus differences affected the strength of crowding. Nevertheless, the performance range as a result of flanker similarity manipulation was modest in both ring and grid configurations.
Such stimulus and organisational differences might also explain why a small number of dissimilar flankers in an otherwise uniform display modestly but statistically worsened performance, relative to when all flankers were similar in Experiment 1, but not in Experiment 2B. That is, the outcomes were different for a subset of conditions in Experiment 2B that were conceptually equivalent to the dense-uniform conditions in Experiment 1. Specifically, the all-flankers-similar, the Radial 1 pair, and the Tangential 1 pair conditions in Experiment 2B were comparable to the ‘None’, ‘Radial’, and ‘Tangential’ conditions, respectively, within the set of dense-uniform conditions in Experiment 1. In the latter set, performance was higher in the ‘None’ condition than in the other two. This difference was on the order of 10 pp. However, in Experiment 2B, all three conditions were indistinguishable. This discrepancy in outcomes might be due to the organisational differences among flankers. It is plausible that the introduction of dissimilar flankers in Experiment 1 broke the coherence of the closest ring, paradoxically releasing its constituent elements to interfere with the target individually. In Experiment 2, flankers were letters arranged in a rectangular grid, sharing response-relevant features with the target. In this configuration, target-flanker grouping was as strong as between flankers; that is, no comparably strong sub-group like a ring existed to be disrupted. Here, introducing dissimilar flankers might affect both target-flanker and flanker-flanker grouping, thus not altering outcomes.
Can Prevalent Theories of Crowding Explain These Results?
Over the course of half a century, several theories of crowding have been proposed. These have however been designed to explain the characteristics of crowding in sparse settings. Would these theories, and their modern successors, which attempt to explain crowding in more complex settings, explain our results?
A common explanation of crowding is that it occurs due to pooling of a target object’s and its flankers’ features (Freeman et al., 2012; Greenwood et al., 2009; Levi, 2008; Parkes et al., 2001; Pelli et al., 2004), where ‘feature’ is defined broadly. This pooling, which can be simple averaging (Greenwood et al., 2009; Parkes et al., 2001; van den Berg et al., 2010) or complex statistical summarisation (Balas et al., 2009; Keshvari & Rosenholtz, 2016), is argued to occur over a limited region of space around the target, sometimes termed the ‘integration field’. Its size is thought to be governed by the Bouma law (Pelli & Tillman, 2008). This law states that the radii of the oval are proportional to the target’s eccentricity, irrespective of the type of objects as long as the targets and flankers are similar. The proportionality constant is variable across individuals but is, on average, half the target eccentricity for radial flankers and a quarter of target eccentricity for tangential flankers. There are exceptions to this rule, with the proportionality constant varying widely depending on stimulus parameters such as duration, object contrast, flanker preview, among others (Rashal & Yeshurun, 2014; Soo et al., 2018; Tripathy & Cavanagh, 2002; Tripathy et al., 2014). Overall, this pooling theory posits a local, but fairly large, zone of interference.
A similar, yet separate, proposal is that crowding arises from the limited resolution of selective attention (Chakravarthi & Cavanagh, 2007; He et al., 1996; Intriligator & Cavanagh, 2001). According to this attentional theory, foci of selective attention needed to select and process an object have a minimum resolution at each retinal location, much coarser than acuity at that location (Intriligator & Cavanagh, 2001). This minimal size is well characterised by the Bouma law. If more than one object falls within the smallest attentional selection window, they cannot be individuated and processed separately, leading to crowding. In this account, dissimilar flankers are filtered out to the extent possible, leading to less crowding.
Such proposals arguing for a limited zone of interference have been challenged by several intriguing findings. The configuration of flankers around a target strongly modulates crowding, even when local features are kept the same. For example, oriented Gabors aligned in the form of a ring around a target Gabor led to minimal crowding compared to when the same Gabors did not form a ring (Livne & Sagi, 2007). Along the same vein, Saarela et al. (2010) found that regularity of spacing between target and flankers increased crowding, while factors that should modulate crowding according to the Bouma law, such as the number of flankers or the proximity of the nearest flanker, did not. Extending this line of research, more recent studies (Herzog et al., 2015; Manassi et al., 2012, 2013) provide strong evidence for ‘uncrowding’ – grouping among flankers to the exclusion of the target leads to much reduced crowding despite an increasing number of flankers. This set of results argues against pooling within a limited region, since the local information remains the same, yet the amount of crowding differs between conditions. In summary, the grouping theory argues that crowding is stronger when the target groups with the flankers, and weaker when the flankers group among themselves but not with the target. Importantly, there is no fixed local zone of interference. The processing of the entire visual scene contributes to the ability to identify any individual element.
Van der Burg et al. (2017), based on their findings that the similarity (or alignment) of only immediately adjacent tangential flankers modulates target identification in dense settings, proposed the nearest neighbour hypothesis. This proposal argues that the Bouma law breaks down in dense displays and only the ability to segment the target, which is aided by the dissimilarity of the nearest neighbours, accounts for crowding. Van der Burg et al. (2024) extended a model of perceptual grouping to explain the mechanisms of the nearest neighbour hypothesis. Their model allows the (neural) activity elicited by an object to be influenced by the activity due to its nearest neighbours, depending on their proximity and similarity. Close and similar objects increase shared activity, which prevents the segmentation of the target, leading to crowding. This is thought to occur in all settings – sparse or dense displays. One could argue that this amounts to grouping of the objects preventing the individuation of the target, but where this process occurs only over adjacent objects. The model does not explicitly incorporate any radial–tangential anisotropy and hence treats both sets of flankers equally. However, the implementation appears to privilege tangential flankers; this is because these flankers are considered as closest to the target when the two-dimensional display is unwrapped in the model’s one-dimensional implementation. This choice is, of course, arbitrary and can be changed to radial flankers being closest. Thus, as implemented, tangential flankers are more powerful. Nevertheless, a 2D implementation of the model could incorporate the standard radial–tangential anisotropy.
The crowding outcomes in the sparse conditions of Experiment 1 can be accommodated by all of the above theories, using well known principles of the role of similarity and radial–tangential anisotropy. The pattern of results in dense settings, however, is different from that in sparse settings. One point to note up front would be that while there is statistical evidence for differences in performance across the various dense conditions, the differences are modest (~10 pp at most). Thus, for all practical purposes we might argue that they are not that different; in fact, no differences were observed for conceptually similar conditions in Experiment 2B.
The pooling and attentional theories of crowding would argue that, in dense conditions, unlike in sparse settings, there is a large number of similar flankers close to the target that will interfere with the target despite the presence of a small number of nearby dissimilar flankers. The similar flankers cause strong crowding, irrespective of the location of the dissimilar flankers, thus explaining the pattern of observed results. However, since these theories adhere to the Bouma law, they would predict that more dissimilar flankers within the integration field should lead to less crowding. Hence, crowding should be weaker in the dense-mixed conditions compared to the dense-uniform conditions. However, we did not find any difference between the two conditions. Nevertheless, pooling theories that depend on high-dimensional statistical summary mechanisms (Balas et al., 2009; Keshvari & Rosenholtz, 2016) would argue that a few dissimilar flankers would not alter the summarisation process much and hence lead to comparable crowding across these conditions. The argument would be analogous for dense-mixed conditions; in such conditions, the different polarity elements would also be pooled or averaged into a texture, within which the target will be incorporated and hence cannot be identified. That is, while simple averaging proposals might find it hard to explain these results, more complex higher-dimensional averaging processes might be able to account for them.
These results can, however, also be explained by grouping and the nearest neighbour hypotheses – the presence of a large number of nearby similar flankers would lead to target-flanker grouping and thus maintain high levels of crowding. The small number of nearby dissimilar flankers might not disrupt these underlying grouping and mutual interference processes.
The results of Experiment 2B can, for the most part, be explained by these theories, in the same way. Strong crowding is induced by the nearby similar objects, despite the presence of a few dissimilar flankers. The modest improvement in performance with increasing number of dissimilar flankers can also be explained by these theories. More dissimilar flankers will produce less interference (pooling and nearest neighbour theories), be filtered out (attention), and/or lead to increased flanker-flanker grouping and hence uncrowding (grouping and nearest neighbour theories).
Importantly, the results of Experiment 2A favour the nearest neighbour hypothesis. It is remarkable that the presence of just one pair of adjacent similar flankers (Radial 1 pair or Tangential 1 pair conditions) among a dense set of dissimilar flankers dramatically reduces performance. Accuracy falls from that in the all-dissimilar-flankers condition (or something close to it) to that in the all-similar-flankers condition. Pooling, high-dimensional summary statistics, or attentional resolution would not expect such a dramatic reduction in performance. They might predict a small reduction in performance relative to the all-dissimilar-flankers condition, due to the small number of nearby similar flankers pooled with the target. But the few flankers should not interfere with the target as much as a dense set of similar flankers. The nearest neighbour hypothesis can, however, accommodate this finding. Van der Burg et al. (2024) found that when a small number of the closest flankers were similar to target, performance was indistinguishable from when all flankers were similar to the target; in these cases, accuracy was close to chance levels. That is, in such a configuration, the similarity of the other, more distant objects in the display did not affect performance. Their results were well accounted by a model that incorporates local grouping and mutual influence among just the nearest neighbours. We find a similar result, where a small number of nearby similar flankers lead to strong crowding.
Task Relevance of Similarity
As noted above, adjacent dissimilar flankers did not modulate target identification performance in dense settings. One alternative explanation for this set of findings attributes it to the task relevance of the similarity manipulation. In our experiments, similarity was manipulated via contrast polarity. However, polarity is irrelevant to the orientation discrimination task used in the current study. Recent evidence seems to suggest that crowding operates independently within feature domains, especially for features such as orientation and surface properties like colour or polarity (Cass & Van der Burg, 2023; Greenwood & Parsons, 2020; Yashar et al., 2019). That is, target-flanker dissimilarity along a task irrelevant feature dimension does not modulate identification performance. For example, if the task is orientation discrimination, colour similarity (or lack of it) between the target and its flankers would not modulate performance, while orientation similarity would. This might explain the lack of a strong effect of dissimilar flankers in our experiments. It is a plausible explanation for our results, particularly in Experiments 1 and 2B. Further, in Experiment 2A, where the presence of just two adjacent similar flankers dramatically reduces identification accuracy might be argued to be due to the fragility of the effect of similarity in the task irrelevant dimension.
While this explanation appears reasonable, there are a few findings that must be taken into consideration. First, both Experiments 2A and 2B demonstrated a strong effect of similarity on crowding; the difference between conditions where all flankers shared the polarity with the target and where all flankers did not share the polarity was substantial ~30 to 40 percentage points. Similar effects have been found in previous studies with task irrelevant similarity manipulations with a large number of flankers (e.g., Manassi et al., 2012). Second, polarity did modulate identification performance in sparse settings (Experiment 1), even though there were similar flankers in some of these conditions (sparse-rad and sparse-tan conditions). Third, the contrast polarity of the target was fixed for each half of the experiments (the order was counterbalanced across participants). This allows for feature-based attention to play a role by enabling filtering of flankers with the opposite polarity (Maunsell & Treue, 2006). This would strengthen any potential weakness in cross-dimensional similarity effects on crowding. Fourth, the presence of target-adjacent flankers similar in polarity did make a large difference in performance (Experiment 2A, where one pair of similar flankers reduced performance dramatically). If similarity along a task irrelevant dimension was not effective, then we should not expect them to make such a large difference.
Fifth, there is substantial evidence that similarity in dimensions other than the task relevant one also strongly modulates crowding. For example, Kooi et al. (1994) found strong effects of similarity for task irrelevant features (similarity in luminance polarity while the task was target orientation, as in this study). Similar findings have been reported across multiple studies (Chakravarthi & Cavanagh, 2007; Kennedy & Whitaker, 2010; Scolari et al., 2007; Soo et al., 2018), where differences in polarity and colour substantially reduced crowding as measured along a different dimension – often orientation discrimination. Overall, it appears that almost any kind of featural difference that leads to ‘pop-out’, even when task-irrelevant, reduces interference. Hence, the Bouma law proposed by Pelli and Tillman (2008) argues that the law specifically applies to similar objects. Further, differences across targets and flankers that are not task relevant nevertheless modulate performance (e.g., contrast: Rashal & Yeshurun, 2014; Soo et al., 2018; transients: Greenwood et al., 2014). Finally, it is interesting to note that the computational model that implements the nearest neighbour hypothesis (the CODE model: Van der Burg et al., 2024) incorporates similarity between adjacent elements, but does not specify that it has to be along task relevant dimensions. Thus, while task relevance of similarity is a plausible alternative, it is likely not the sole or main determinant of outcomes in dense contexts. The influence of the nearest neighbour is probably driving performance.
Conclusion
We tested the effect of similarity in relation to radial–tangential anisotropy on crowding in dense settings. We found that the patterns of results are different for sparse and dense flankers, which grouping cannot easily explain. Importantly, a small number of similar flankers near a target among a dense set of dissimilar flankers lead to strong crowding, which is best explained by the nearest neighbour hypothesis of crowding.
Footnotes
Acknowledgements
I thank Amarachi Orisakwe, Eric Stenbaum, and Glen McKay for help with data collection, Marlene Poncet for discussions throughout the project and for helpful comments on multiple drafts, Mauro Manassi, Nika Adamian, and Soren Andersen for discussions about the interpretation of the results, and Oliver Hamlet for extensive discussions on Close Proximity Gathering and its connections to visual crowding.
Ethical Considerations
Experiment 1 received ethical approval from the Psychology Ethics Committee at the University of Aberdeen (PEC/5131/2022/10). Experiments 2A and 2B also received ethical approval from the same committee (ID numbers: 1521176 and 2178327, respectively).
Consent to Participate
Participants provided written informed consent.
Funding
The author received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.