
Abstract
Self-prioritization effects found in label–shape matching tasks have been demonstrated extensively. Participants are faster to match label–shape label–shape stimuli against a prelearned mapping, the more self-related the label is. The effect is thought to be driven by special cognitive treatment, saliency, or attentional prioritization of self-referential stimuli. However, the exact underlying mechanism remains elusive. We propose an alternative explanation, namely that self-related labels facilitate matching performance by inducing the so-called self-positivity bias (people tend to represent themselves more positively than they represent others), which is conducive to matching. To test this idea, we skipped the learning of label–shape mappings in our task. Furthermore, we used a shape–shape matching task. Thus, participants did not have to determine whether label–shape combinations match a previously learned mapping, but merely whether two geometrical figures are the same (match) or not (mismatch). Crucially, each trial was preceded by a more or less self-related prime word. This setup enabled us to measure self-prioritization effects in the absence of task directed links between target and self-related stimuli. In Experiment 1, self-related labels (“YOU”, “FRIEND”, “STRANGER”) resulted in faster matching responses. Experiment 2 replicated the effect and ruled out word length as an alternative explanation (prime words: “YOURSELF”, “FRIEND”, and “OTHER”). Our findings suggest that the self-positivity bias at least contributes to self-prioritization effects. Furthermore, our theory explains interaction effects between match versus mismatch trials and self-relatedness of the labels, as found in many experiments.
Introduction
Not all stimuli are equal: some are more interesting to us than others. How interesting a particular stimulus is depends on the circumstances and the current interests of the perceiver, but some stimuli seem to be interesting no matter what. Numerous studies indicate that stimuli with a certain self-reference attract more attention than stimuli that refer to other individuals. These studies typically use label–shape matching tasks, in which participants have to make speeded judgments whether presented label–shape combinations match prelearned label–shape mappings. To exemplify, before starting the matching task, participants learn to map geometric shapes to the words “YOU”, “FRIEND”, and “STRANGER” (for instance “YOU”is mapped to a circle, “FRIEND” is mapped to a triangle, and “STRANGER” is mapped to a square). During the actual matching task, participants have to make speeded judgments on whether a presented label–shape combination complies with the learned mappings (match) or not (mismatch). Participants tend to perform quicker when matching “YOU” with its shape, as compared to the other two less self-referential words.
Such label–shape matching task findings were found to generalize across age groups (Maire et al., 2020; Singh & Karnick, 2022; Sui & Humphreys, 2017), and work with ingroup/outgroup labels (Enock et al., 2018; Moradi et al., 2015), avatars (Mattan et al., 2015; Woźniak & Knoblich, 2019), future- and past-selves versus current self (Golubickis et al., 2017), own versus others’ faces (Woźniak & Hohwy, 2020), and relevant versus irrelevant identity labels (Macrae et al., 2018). Altogether, such studies strongly suggest a processing advantage if, and to the degree that, the targets of cognitive tasks refer to, and presumably activate, some kind of self-related representation (Sui & Humphrey, 2015a).
Previous work has concentrated on the self-referential nature of the target stimuli and attributed the boosted performance for self-referential stimuli to the special cognitive treatment, saliency, and attentional prioritization that these stimuli are supposed to enjoy (Constable et al., 2019; Humphreys & Sui, 2015; Reuther & Chakravarthi, 2017; Stein et al., 2016; Sui & Humphreys, 2015b; Sui et al., 2013). However, accounts of that kind overlook the systematic outcome of many studies on self-reference. If self-related stimuli really induced a heightened degree of saliency or arousal, and if that would produce the effect, one would expect both match trials and mismatch trials to benefit from self-related stimuli. However, the self-related benefit is commonly restricted to the match trials (for an overview, see Woźniak & Knoblich, 2019).
Here, we argue that this observation invites an alternative hypothesis. Note that match versus mismatch decisions differ with respect to their affective quality: match decisions imply a positive judgment, whereas mismatch decisions imply a negative judgment. Interestingly, there is strong evidence that thinking about oneself induces positivity, as evident from positive implicit self-evaluations (Gawronski & Bodenhausen, 2014; Greenwald & Banaji, 1995). Many believe such self-positivity bias phenomena (for a review see: Mezulis et al., 2004) to be caused by the need for a positive self-view (Allport, 1955; Epstein, 1973; Heine et al., 1999; James, 1890). More methodologically directly relevant evidence for a positive connotation of self-labels was found in a word-label matching task in which rewards and positive emotion labels, instead of self-related words, produce a similar result pattern to that of the self-reference effect (Stolte et al., 2017; Sui & Humphreys, 2015a). Accordingly, it is possible that presenting self-related words or symbols simply primes responses with a positive connotation. Hence, it may not have been the case that participants in the Sui and Humphreys studies were faster at matching self-related stimuli because these induced heightened saliency, arousal or special cognitive treatment; rather, it may have been the mere presence of a self-related stimulus that primes positive match decisions. Although both explanations predict faster matching performance for more self-related stimuli, only our alternative explanation predicts no advantage or even a disadvantage for self-related stimuli during mismatch trials (Woźniak & Knoblich, 2019). Another crucial difference between the two explanations is that our explanation does not require the imperative stimulus itself to be task- (self-) relevant, merely presenting a non-task relevant but self-relevant prime before the imperative stimulus should be sufficient to induce the so-called self-prioritization effect.
The present two experiments aimed to test our hypothesis. There is ample evidence that cognitive processes can be triggered by nominally irrelevant stimuli presented just before the imperative stimulus (Fazio et al., 1986). We applied this technique to disentangle the self-related stimulus, which we assume to be responsible for the priming of responses with a positive connotation (match responses in our case), from the actual imperative stimulus that people responded to. Hence, in the current study, the self-related stimulus was a mere prime of no task relevance that preceded the task-related stimuli, which consisted of pairs of shapes that did or did not match. We predicted that self-related stimuli would facilitate “match” responses but not “mismatch” responses. In Experiment 1, we primed with the words “YOU”, “FRIEND”, or “STRANGER”, and in Experiment 2, we controlled for word length effects by priming with “YOURSELF”, “FRIEND”, or “OTHER”. Lastly, to test whether the two experiments produced statistically equivalent patterns, we analyzed both experiments equating “YOU” with “YOURSELF” and “STRANGER” with “OTHER”.
Experiment 1
In each go-trial of Experiment 1, we primed participants with the words “YOU”, “FRIEND”, or “STRANGER” before presenting them with an unrelated shape-shape matching task wherein they had to judge whether two shapes matched or mismatched. Given that the prime words were entirely task-irrelevant and thus at riskof being ignored, we added no-go trials to ensure some depth of processing of the prime words. Specifically, we presented one of three animal words (“YAK”, “FERRET”, “SKUNK”) in a few trials, in which case participants were to refrain from responding, thus making processing of the prime words task relevant for successful no-go trial performance.
Method
Participants
Taking into account previous studies (Sui et al., 2012) and expecting more noise and performance-related exclusions due to online data collection (for another online self-reference study, see Bentley et al., 2017), we aimed to collect data from about 60 participants (more than three times as many as Sui et al., 2012). Fifty-nine English-speaking adults (38 female, 20 male, 1 unknown) between 18 to 35 years old (Mage = 23.14, SD = 5.29) participated for $1.50 or a course credit (25 from MTurk, 34 from SONA). A further 12 MTurk participants were rejected because they either did not belong to the required age range or they were left-handed. Twelve further participants (11 from MTurk, 2 from SONA) were excluded due to poor performance (less than 65% correct on match-, mismatch- and no-go trials). Participants reported normal motor control of their arm, normal or corrected-to-normal vision, good command of English, and no current or previous psychiatric or neurological diagnosis. Finally, all participants were naïve to the purposes of the experiment. For MTurk, we required a minimum of 50 previous submissions and a 95% approval rate. The experiment took around 15 min to complete. All studies reported here were approved by the psychology ethics committee of Leiden University and conformed to the Declaration of Helsinki. Informed consent and demographics were collected via Qualtrics (2005) (Qualtrics, Provo, UT, USA).
Procedure
The experiment was programmed in Open Sesame (OSWeb 1.3.8, Mathôt et al., 2012). Participants first read the information letter and provided written consent, and then completed a demographic questionnaire, providing their age, gender, handedness and MTurk ID. They then proceeded to the experiment. After the instruction, the session started with 36 practice trials. Participants could commence to the experimental trials if their overall accuracy was above 70% but otherwise had to undergo another practice block. During the practice trials, feedback (“correct”, “incorrect” or “too slow”) was presented for 500 ms at the end of each trial. The experimental phase consisted of 3 blocks of 144 trials each (126 go-trials and 18 no-go trials). Participants were informed of their overall accuracy at the end of each block. Responses to no-go trials were recorded, and participants who falsely responded to more than 35% of no-go-, match- and mismatch- trials were excluded from the final analysis to ensure data quality. Match and mismatch trials were presented equally often in a random order. Prime words (“YOU”, “FRIEND”, and “STRANGER”) and different shape combinations were counterbalanced and presented in random order. The animal words “YAK”, “FERRET”, “SKUNK” served as no-go stimuli. Participants thus were instructed not to press any key after an animal word and wait for the next word; yet, when they were presented with a prime word, they had to indicate whether the given shapes were matched or not. During instruction, the animal words were introduced with the photos of the animals to eliminate any lack of knowledge.
All stimuli were designed to copy those described by Sui et al. (2012) in terms of size, color, and location. Stimuli were presented with 1,280-by-800-pixel resolution and on a gray background (RGB[168, 168, 168]). Go-trials started with the presentation of one of the three prime words (“YOU”, “FRIEND”, and “STRANGER”) at the center of the screen (mono font in 25 pixels) for 500 ms. Subsequently, the word remained on the screen, and two geometric shapes were presented above and below the identity labels (scale 0.5, 32 by 32 pixels) for 200 ms. The shapes for the matching task consisted of white outlines. The upper shape was located on x = 0, y = −192, while the lower shape was located on x = 0, y = 192. These stimuli were followed by a blank screen, which lasted until response or 1,000 ms maximum. Participants were asked to indicate whether the shape pairings were matched (identical shapes) or mismatched (different shapes) as quickly and accurately as possible by using one of the two keys (v or m) with the two index fingers. The assignment of the keys (match or mismatch) was counterbalanced between participants.
Results and Discussion
After excluding trials with reaction times (RTs) below 150 ms (5% of all trials), we calculated the mean accuracy of participants on go-trials (match and mismatch trials), which was 84.6% (SD = 7.55); and on no-go trials, it was 84.75% (SD = 7.61). We then analyzed RTs and error rates using a repeated-measures ANOVA, as a function of trial type (match, mismatch; within-subjects factor) and prime words (“YOU”, “FRIEND”, and “STRANGER”; within-subjects factor). Mauchly’s test indicated that the assumption of sphericity had been violated for the effect of word; therefore, we implemented Greenhouse–Geisser corrections. The RT analysis yielded a significant main effect of trial type, F(1, 58) = 144.61, p < .001,
Separate ANOVAs for the two trial types showed that the effect of prime word was significant for match trials, F(2,116) = 4.45, p = .021,
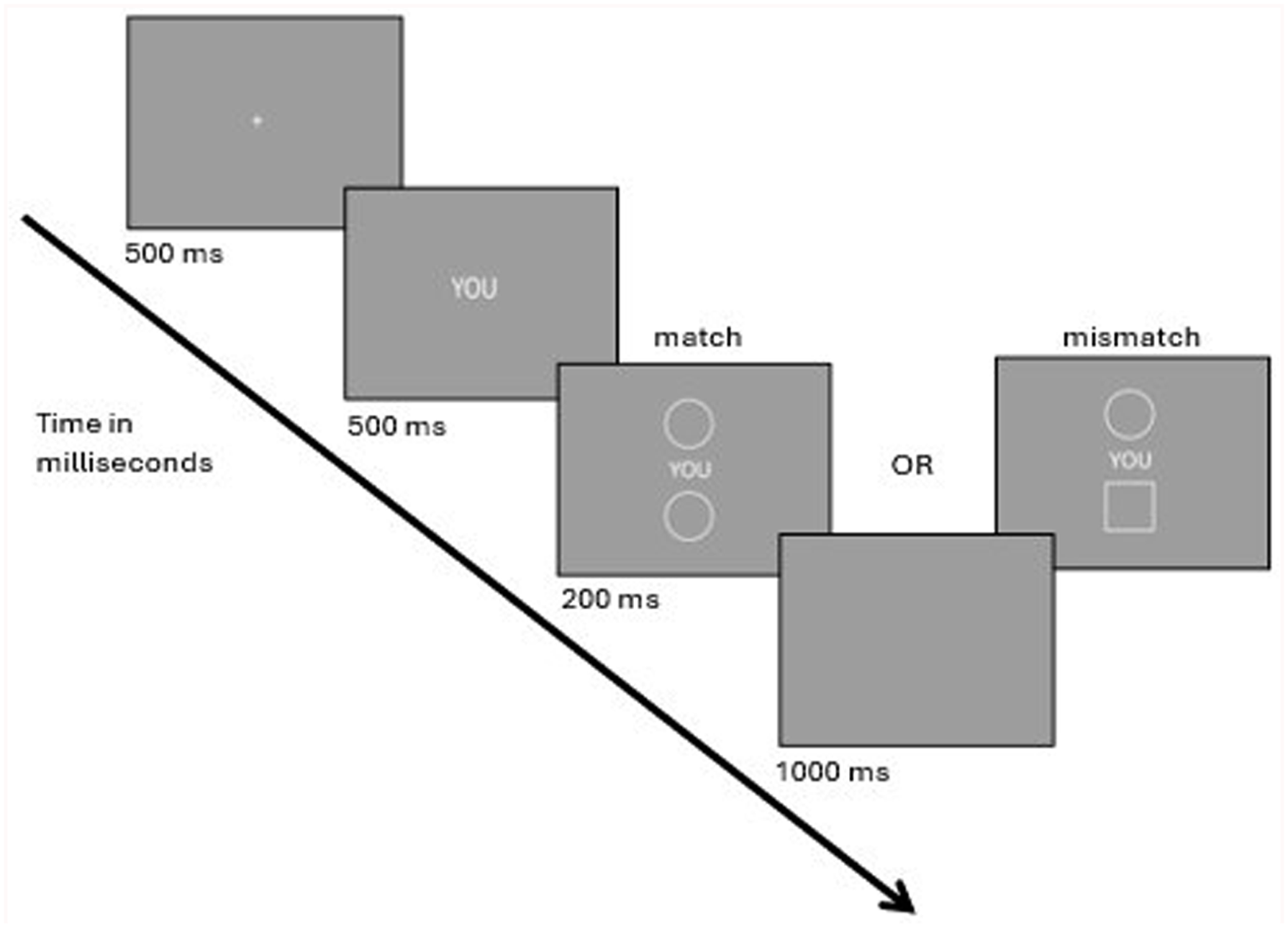
Overview of experimental setup.
Taken altogether, we produced what convincingly looks like a self-prioritization effect. Yet, our paradigm should not produce such an effect according to conventional self-prioritization theorizing. Just like in earlier studies, we found performance to be best in conditions where self-related stimuli were presented. However, this time, these stimuli did not require any response and were not selectively related to one or some of the stimuli to be matched in the task. Furthermore, we replicated previous studies that found the self-prioritization effect only on matching trials.
Experiment 2
Even though the findings of Experiment 1 were as expected, we wondered whether physical features of the priming words might have played a role in generating the effect. Obviously, the priming words were of different lengths, which may have led to different degrees to which they were attracting or distracting attention, which, in turn might have affected our findings. We note that the face validity of this possibility is low, because Sui et al. (2012) have already provided evidence that word length is unlikely to account for the original self-reference effect (Experiment 3C). However, since our task differs from the original design, especially because of the absence of any previous learning of mappings, we conducted Experiment 2 to ascertain that word length is not a factor. The experiment was almost identical to Experiment 1, except that the short word “YOU” was replaced by the word “YOURSELF”, which occupied more physical space and took longer to be read than any of the five ‘OTHER’ words, and the relatively long word “STRANGER” was replaced by the shorter word “OTHER”. If word length were to account for our findings in Experiment 1, performance should be particularly poor for the “YOURSELF” prime, especially in matching trials.
Method
Participants
We tested and analyzed data from 59 adults (39 female, 20 male) between 18 to 29 years old (Mage = 20.58, SD = 2.60), who participated for $1.50 or course credits (53 from SONA, 6 from Prolific). One additional SONA participant was excluded due to left-handedness, and three Prolific participants for poor performance (less than 65% correct on match, mismatch, and no-go trials). The procedure was exactly as in Experiment 1, except that the prime word “YOU” was replaced by “YOURSELF” and the word “STRANGER” was replaced by “OTHER”.
Results
After excluding trials with RTs below 150 ms (8.6% of all trials), we calculated the mean accuracy of participants on go-trials (match and mismatch trials), which was 83.5% (SD = 8.23); and on no-go trials, it was 83.50% (SD = 8.11). In the RT ANOVA, the main effect of trial type was significant, F(1, 58) = 80.12, p < .001,
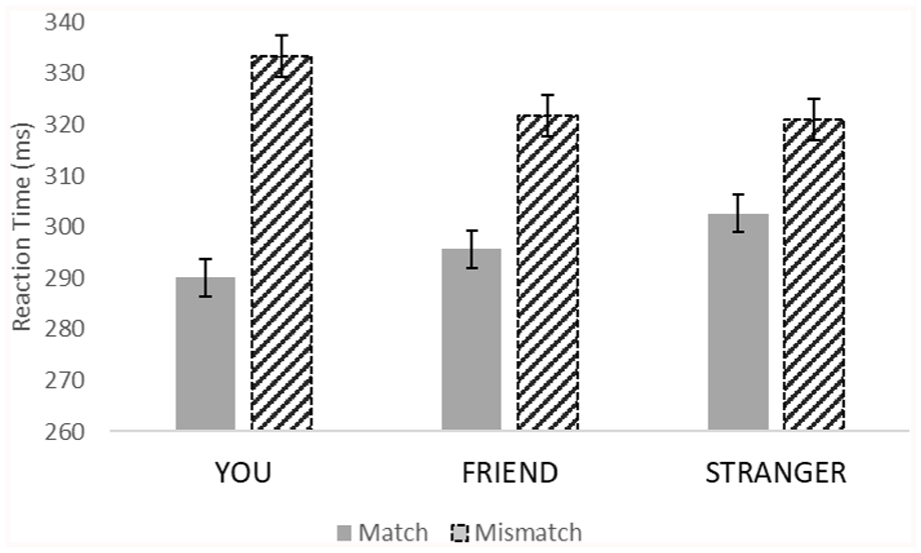
The interaction between trial type and self-reference on reaction time with standard errors in Experiment 1.
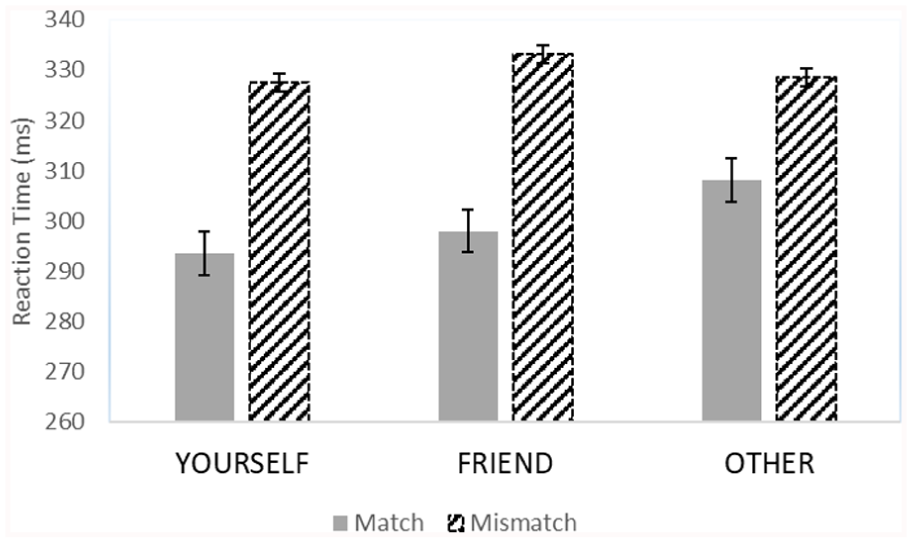
The interaction between trial type and self-reference on reaction time with standard errors in Experiment 2.
To test whether the outcome patterns were statistically equivalent for the relevant matching trials across Experiments 1 and 2, we ran an ANOVA across the data from both experiments (equating “YOU” with “YOURSELF” and “STRANGER” with “OTHER”), with experiment as an additional between-subjects factor. The experiment factor was only involved in the significant three-way interaction, F(2, 230) = 3.87, p = .022,
General Discussion
Our findings demonstrate that self-prioritization effects along the lines of Sui et al. (2012), can be found even if participants do not respond to self-related stimuli. The mere exposure to such stimuli is sufficient to speed up match responses, but not mismatch responses. We take this pattern to imply that special theorizing is not needed for explaining self-prioritization effects. It seems sufficient to assume that self-related stimuli have a more positive connotation than self-unrelated stimuli. If we assume that people represent themselves in terms of representational networks of feature codes that are describing them (Me-Files, in the terminology of Hommel, 2018), this suggests that self-representations include positive codes (or more positive than negative codes). Accordingly, if a stimulus is triggering the self-representation, other events that also include positive feature codes are primed (Eder et al., 2012). This includes match responses but not mismatch responses, which accounts for the observation that self-related stimuli speed up the former but not the latter.
Despite minor numerical differences between our two experiments, the overall ANOVA suggests that the outcome patterns were statistically equivalent for the relevant matching trials. It is true that Sui et al. (2012) reported larger effects (up to 175 ms,
Although we do not rule out additional mechanisms causing self-reference effects in the label–shape tasks, we consider our priming account as more consistent with Occam’s razor (parsimony) than other more elaborate explanations of the self-prioritization effect, as it views this effect as just another example of a larger set of observations that all point to the apparent fact that people tend to represent themselves as more positive than they represent others. This parsimonious account thus challenges accounts that claim separable, independent processes underlying self- and positive emotion/reward biases (Stolte et al., 2017; Sui & Humphreys, 2015a; Sui et al., 2013; Yankouskaya et al., 2023). Furthermore, our account is at odds with the Self-Attention Network framework (Humphreys & Sui, 2015). This account claims that self-relevant stimuli are powerful cues for attention and can modulate performance in a relatively automatic manner. However, such automatic attention manipulation would not predict the often-found interaction between match-mismatch behavior and self-relevance (none of the aforementioned theories do), whereas our account does.
We acknowledge that our method differs substantially from the typical label–shape matching tasks producing self-prioritization effects (Sui et al., 2012). These differences may indeed contribute to the larger effect sizes in the Sui et al. (2012) study. However, they may also trigger different underlying mechanisms engaged during the tasks. For instance, the typical label–shape matching tasks rely on memory retrieval of the prelearned mappings. However, logically, the traditional label–shape matching tasks temporally allow for the effect as demonstrated here to unfold during the presentation of the label–shape pair. Thus, conservatively, our study at least makes a strong case for the self-prioritization effect being partly caused by positive response tendencies caused by self-relatedness of the stimulus, with more self-relatedness of the stimulus inducing quicker matching, but not mismatching. Moreover, this case becomes stronger due to the fact that the predicted and found interaction between self-relatedness and matching versus mismatching is explained by our priming approach, but not by any of the traditional explanations.
Supplemental Material
sj-sav-1-qjp-10.1177_17470218251389726 – Supplemental material for Response Priming Accounts for Self-Prioritization Effects
Supplemental material, sj-sav-1-qjp-10.1177_17470218251389726 for Response Priming Accounts for Self-Prioritization Effects by Stephan Verschoor, Asmin Yurtsever, Tanja Schultz and Bernhard Hommel in Quarterly Journal of Experimental Psychology
Supplemental Material
sj-sav-2-qjp-10.1177_17470218251389726 – Supplemental material for Response Priming Accounts for Self-Prioritization Effects
Supplemental material, sj-sav-2-qjp-10.1177_17470218251389726 for Response Priming Accounts for Self-Prioritization Effects by Stephan Verschoor, Asmin Yurtsever, Tanja Schultz and Bernhard Hommel in Quarterly Journal of Experimental Psychology
Supplemental Material
sj-sav-3-qjp-10.1177_17470218251389726 – Supplemental material for Response Priming Accounts for Self-Prioritization Effects
Supplemental material, sj-sav-3-qjp-10.1177_17470218251389726 for Response Priming Accounts for Self-Prioritization Effects by Stephan Verschoor, Asmin Yurtsever, Tanja Schultz and Bernhard Hommel in Quarterly Journal of Experimental Psychology
Footnotes
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) – SPP 2134, 402790609 and 467058220.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
Supplemental Material
The Supplemental Material is available at: qjep.sagepub.com, ![]() .
.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.