
Abstract
In the present two experiments, we explore the possibility of swift attenuation of capture by irrelevant features in the contingent-capture protocol. Some prior research suggests that feature attenuation might be most efficient for fixed, anticipated irrelevant features and that varying irrelevant features from trial to trial can undermine their successful attenuation. Here, we exploited this dependence of attenuation on feature certainty to test if attenuation contributed to contingent-capture effects in a capture-probe version of the contingent-capture protocol. In line with the swift attenuation of irrelevant features, salient but target-dissimilar singleton cues that were consistently coloured diminished recall of probes at their locations. This was in comparison to inconsistently coloured target-dissimilar singleton cues. Nonetheless, probe-recall was still better at target-dissimilar cue locations than at non-singleton locations in the cueing display, indicating attenuation of task-irrelevant features rather than their complete suppression.
Introduction
A long-standing debate in visual attention research concerns the extent to which task-irrelevant features capture attention. Proponents in this debate traditionally defended a bottom-up (e.g., Theeuwes, 2010) or a top-down view (e.g., Folk et al., 1992) of visual attention. According to the bottom-up perspective, salient stimuli capture attention automatically, overriding top-down goals of the observer. In contrast, the top-down perspective suggests that only stimuli matching an observer’s current search goals capture attention, while salient but task-irrelevant stimuli are ignored. Recently, the debate progressed towards more integrative views, detailing the contributions of both factors (e.g., Gaspelin & Luck, 2018b).
Evidence for the bottom-up perspective comes from the additional singleton protocol (Theeuwes, 1992): When instructed to search for a shape singleton (i.e., a unique shape among homogeneous non-singleton/distractor shapes), presenting a salient-but-irrelevant colour singleton (e.g., one green stimulus among red non-singletons) at a non-target location delays observer’s search for the target. These singleton-presence costs seem to support a bottom-up view of visual attention as attention is seemingly captured by the salient colour distractor, overriding participants’ goal to search for shapes. However, Bacon and Egeth (1994) argued that attention capture in the additional singleton protocol is not necessarily bottom-up. Instead, participants might have searched for “uniqueness” (i.e., were in singleton-detection mode), as searching for a feature discontinuity is sufficient to find the shape-singleton target in half of the trials (i.e., in distractor-absent trials). In distractor-present trials, the salient colour distractor would have also matched the top-down set for uniqueness and captured attention in a top-down-driven manner.
Bacon and Egeth (1994) demonstrated that salient colour distractors failed to capture attention when participants were in feature-search mode. Presenting the predefined target shape among differently shaped distractors (e.g., a diamond among circles and various polygons) forced participants to look for the specific target shape. Under these conditions, there were no singleton-presence costs of the salient colour distractors. This could be due to the lack of match between the singleton distractor and search goals of the participants. However, Gaspelin et al. (2015) argue that attenuation, in the form of active suppression, of known irrelevant singleton colours could contribute to the lack of interference or capture by the singleton.
Gaspelin et al. (2015) tested their hypothesis in a capture-probe version (Kim & Cave, 1995) of the additional singleton paradigm: Participants searched for a specific target shape among heterogeneous distractor shapes while trying to ignore an irrelevant but salient colour-singleton distractor presented away from the target in half of the trials. In one-third of all trials, Gaspelin et al. (2015) briefly presented letters at all target and distractor positions and asked their participants to report as many letters as possible. They found the highest recall for letters presented at the target location, while recall for letters at the irrelevant singleton distractor locations was reduced relative to letters at any non-salient (i.e., non-singleton) distractor position. This lower-than-baseline performance suggests that salient-but-irrelevant distractors are not merely ignored but that their processing is even actively attenuated, in fact, that it is suppressed relative to non-singleton distractors (cf. Chelazzi et al., 2019).
Importantly in the context of the present study, Kerzel and Barras (2016) identified a critical prerequisite for more effective attenuation in the form of successful suppression of singleton distractors: their feature consistency. These authors found no attention capture by consistently coloured singleton distractors (e.g., if the distractor was always red) during the search for a shape, replicating Bacon and Egeth (1994). However, when the colour of the salient distractor varied unpredictably, it interfered with target-search performance, implying failed attenuation of inconsistently coloured singleton distractors (see Ma & Abrams, 2023; Won et al., 2019).
Here, we tested the extent to which task-irrelevant but salient features are processed in the contingent-capture protocol (Folk et al., 1992; for a meta-analysis, see Büsel et al., 2020). Folk et al. (1992) used spatial cueing while participants searched for a feature-defined (e.g., green) target. Briefly, before the target-search display, a spatially non-informative cue appeared at one of the potential target locations: either a valid cue at the subsequent target’s position or an invalid cue at a subsequent distractor’s position. Salient singleton cues either carried a searched-for target feature (e.g., green) or not (e.g., red). Folk t al. (1992) found a top-down contingent-capture effect: cue validity effects (i.e., faster search times in valid than in invalid trials) for target-similar cues—that is, cues matching the participants’ top-down attentional set—but not for target-dissimilar cues that did not match participant’s attentional set.
By now, it is certain that attentional priority for top-down relevant features contributes to the contingent-capture effect. According to the contingent-capture hypothesis, only cues that match a search template for the searched-for target feature capture attention, whereas cues not matching the template do not. In contrast, the extent to which non-matching cues lead to attention capture and/or increased feature processing at their location is less clear: Previously, the lack of validity effects by non-matching cues has often been interpreted as an incapability of non-matching cues to attract attention (e.g., Folk et al., 1992). More recent studies have demonstrated, however, that there are boundary conditions in which even non-matching cues elicited validity effects. For example, Folk and Remington (2015) explained that task-irrelevant abrupt onset cues lead to a validity effect during colour search, when they are rare enough to surprise participants. Temporal aspects have also been shown to influence whether or not abrupt onsets capture attention, as Adams et al. (2022) demonstrated validity effects for onset cues at 0 ms stimulus-onset asynchrony (SOA), and Ruthruff et al. (2019) at longer SOAs. Gaspelin et al. (2016) identified search difficulty as an additional factor that conceals attention capture by (non-matching) onset cues: During easy search conditions, in which target and distractor colours are highly distinct, onset cues do not lead to a validity effect, but when target and distractor colours are very similar, the same task-irrelevant onset cues do lead to a validity effect.
Another common finding within the conventional contingent-capture protocol is same-location costs (SLCs; e.g., Carmel & Lamy, 2015). Here, non-matching cues do not merely fail to elicit a validity effect but actually trigger an inverted validity effect (i.e., faster reaction times [RTs] in invalid trials than in valid trials), which some authors attributed to suppression (Belopolsky et al., 2010; Schoeberl et al., 2018; but see Carmel & Lamy, 2014). Further studies on the role of suppression in the contingent-capture protocol were not entirely conclusive. Burnham (2020) combined the capture-probe procedure with the contingent-capture protocol: Participants performed a search task on the majority of trials (the search trials). In the remaining trials (the probe trials), after the cues, letters appeared at all potential target locations, and participants reported as many letters as possible. Results showed that probe-recall was worse for letters at locations of target-dissimilar/non-matching cues than at locations of target-similar/matching cues (Burnham, 2020). Critically, probe-recall at locations of target-dissimilar cues was higher than that at un-cued (non-singleton) locations, thereby missing the hallmark of suppression used by Gaspelin et al. (2015). Note, however, that even unsuccessful suppression allows for degrees of attenuation of the irrelevant features. For example, although participants might not be able to successfully suppress attention capture on each and every trial, it is possible that, on average, active attenuation based on an occasionally successful suppression of the singletons contributed to their weaker capture effects in comparison to the relevant, top-down matching cues.
Similarly, most electrophysiological evidence also suggests that non-matching cues are not suppressed. More specifically, researchers did not find a distractor positivity (PD) in response to non-matching cues (e.g., Eimer & Kiss, 2010; Grubert & Eimer, 2016; Kerzel & Huynh Cong, 2021; Livingstone et al., 2017). However, there are exceptions to this rule. When Goller et al. (2020) used an irrelevant singleton cue of the same colour as was used for a target-display distractor, they observed a PD to the irrelevant cue, despite the fact that this was an abrupt-onset cue (Experiment 1 of Goller et al., 2020). In addition, a recent study by Harris et al. (2022) reported a significant and numerically larger PD for non-matching cues than for matching cues. And again, the question is whether a PD tells the full story about the processing of non-matching cues. Even in the absence of evidence for overall net suppression—that is, a lacking PD—it is possible that a mixture of occasionally successful suppression and capture or degrees of attenuation modulated the processing of and the capture by non-matching cues. To further underline this, Barras and Kerzel (2016) found no PD towards a colour distractor in the feature search mode. Behaviourally, however, Gaspelin et al. (2015) found evidence for direct and active suppression of salient colour distractors under feature search conditions in their capture-probe task.
What is clear so far is that attentional capture (e.g., validity and/or interference effects) and suppression effects (capture-probe performance) can vary, leaving ample space for the possibility of feature attenuation. We, therefore, aimed to address this possibility in the current study. First, to tap into swift initial attenuation, we introduced an SOA manipulation where, in 0-ms cue-probe SOA conditions, probes were presented together with the cues, at cue onset. In 150-ms cue-probe SOA conditions, probes were presented 150 ms after the cue. This allowed us to tap into potential early modulations of processing priorities at salient-but-irrelevant cue locations (Adams et al., 2022; Arita et al., 2012) and to see if active inhibition (or attenuation) builds up over time (cf. Kim & Cave, 1995; Moher & Egeth, 2012). Second, we manipulated the colour consistency of salient target-dissimilar/non-matching cues: While non-matching cues were consistently coloured throughout a whole block (blue), in another block, non-matching cue colour unpredictably varied from trial to trial, under inconsistent conditions (blue, green, and yellow). Both measures (of SOA and of consistency) were taken in a capture-probe variant of the contingent-capture protocol. If attenuation of features of non-matching cues plays a role in the contingent-capture protocol, we expect a diminished probe (here: digit) report from target-dissimilar cue positions consistently compared with inconsistently coloured non-matching cue conditions. As is the case for suppression, an (on average) attenuation (e.g., in the form of occasionally successful suppression) should be easier where the salient feature can be predicted (under consistent conditions) than where it cannot be predicted (under inconsistent conditions). In addition, if attenuation is based on proactive monitoring for the to-be-attenuated features, attenuation effects could theoretically be observed even under 0-ms SOA conditions (cf. Arita et al., 2012). However, if attenuation takes time to build up and participants first have to attend any salient singleton, even if irrelevant, evidence for the effect of attenuation could be restricted to the longer SOA condition (cf. Kim & Cave, 1995).
In analogy to the arguments of Wöstmann et al. (2022) for the choice of the specific evidence criterion for suppression, it is imperative to spell out which criterion we consider to represent (more vs. less) attenuation in the current study. We took a significantly weaker accuracy of probe reports of cued probes under (consistent vs. inconsistent) non-matching conditions relative to matching conditions as a criterion of (more vs. less) attenuation of cue processing. Note that it would usually be difficult to decide if the lower advantage of cued non-matching than matching cues reflects less versus more capture of attention or less versus more attenuation. However, the currently hypothesised difference of a stronger drop in probe-recall performance between cued non-matching relative to matching conditions under consistent versus inconsistent (non-matching cue colour) conditions would be predicted by a modulating role of attenuation only.
Experiment 1
Methods
Participants
Thirty-nine participants were tested in the present experiment. Participants were students at the University of Innsbruck, gave informed consent, and received course credit for their participation. We determined our sample size based on several considerations: Burnham (2020) estimated Gaspelin et al.’s (2015) suppression effect to be dz = .7879. According to this estimate, we would have needed 12 participants to detect attenuation effects of the same size with a power of 80% and an α-level of .05 (one-sided). However, we were interested not only in attenuation effects within one non-matching cue condition (i.e., consistent) but also in differences between subjects in the two non-matching (i.e., consistent vs. inconsistent) cue conditions, as well as their temporal development. Due to these newly introduced factors and the need to compensate for random influences under online or unstandardised experimental conditions (see the following sections), we chose to test a considerably higher number of participants.
Apparatus
The present experiment was conducted online. Stimulus presentation and response collection were managed by OpenSesame (Mathôt et al., 2012). Experiments were hosted on JATOS (Lange et al., 2015).
Stimuli
As the experiment was conducted online, the exact colour values (e.g., in the CIE colour space) and stimulus sizes are unknown. Hence, we report RGB values and pixels. In the fixation display, four grey (RGB: 128, 128, 128) circles (diameter: 64 px; line thickness: 1 px) surrounded a central fixation cross (8 px × 8 px) at the corners of an imaginary square with a side length of 192 px. In the cueing display, the circles thickened (11 px), and random white digits between 0 and 9 were presented within the circles. One circle was a colour singleton, while the remaining three circles were grey non-singletons. In search trials (70% of trials), following another brief fixation display, four coloured Ts (29 px × 36 px) were presented within the circles. Two Ts were rotated 90° clockwise, and two in 90° counter-clockwise. One of the Ts was coloured in the relevant target colour (red; 140, 0, 0), while the remaining three Ts in the target-search display were coloured cyan (0, 65, 65), magenta (90, 50, 90), and grey (see the previous section). Thus, singleton search was prevented, and feature (colour) search was enforced. In the consistent non-matching cue colour blocks (see below), the non-matching cue was always blue (0, 0, 255). In the inconsistent block, across trials, the non-matching cue could randomly be blue, yellow (255, 255, 0), or green (0, 128, 0).
Design and procedure
For the present experiment, we used a 2 × 2 × 2 × 2 factorial design consisting of four blocks. The two factors validity (valid and invalid) and cue-match (matching and non-matching) were varied within each block. The remaining two factors, non-matching cue consistency (consistent and inconsistent) and cue-probe SOA (0 ms and 150 ms), were varied between blocks. Block order was balanced across participants. Within each block, participants completed 70% search and 30% probe trials. In search trials, the fixation display was presented for 1 s. Next, the cueing display appeared for 100 ms. Following an inter-stimulus interval of 50 ms, the target-search display was presented for 50 ms. An exemplary trial can be seen in Figure 1a. Participants had to search for the colour-defined target and to press a key depending on the orientation of the T inside the target. Cues were predictive of neither the upcoming target position nor the target orientation.
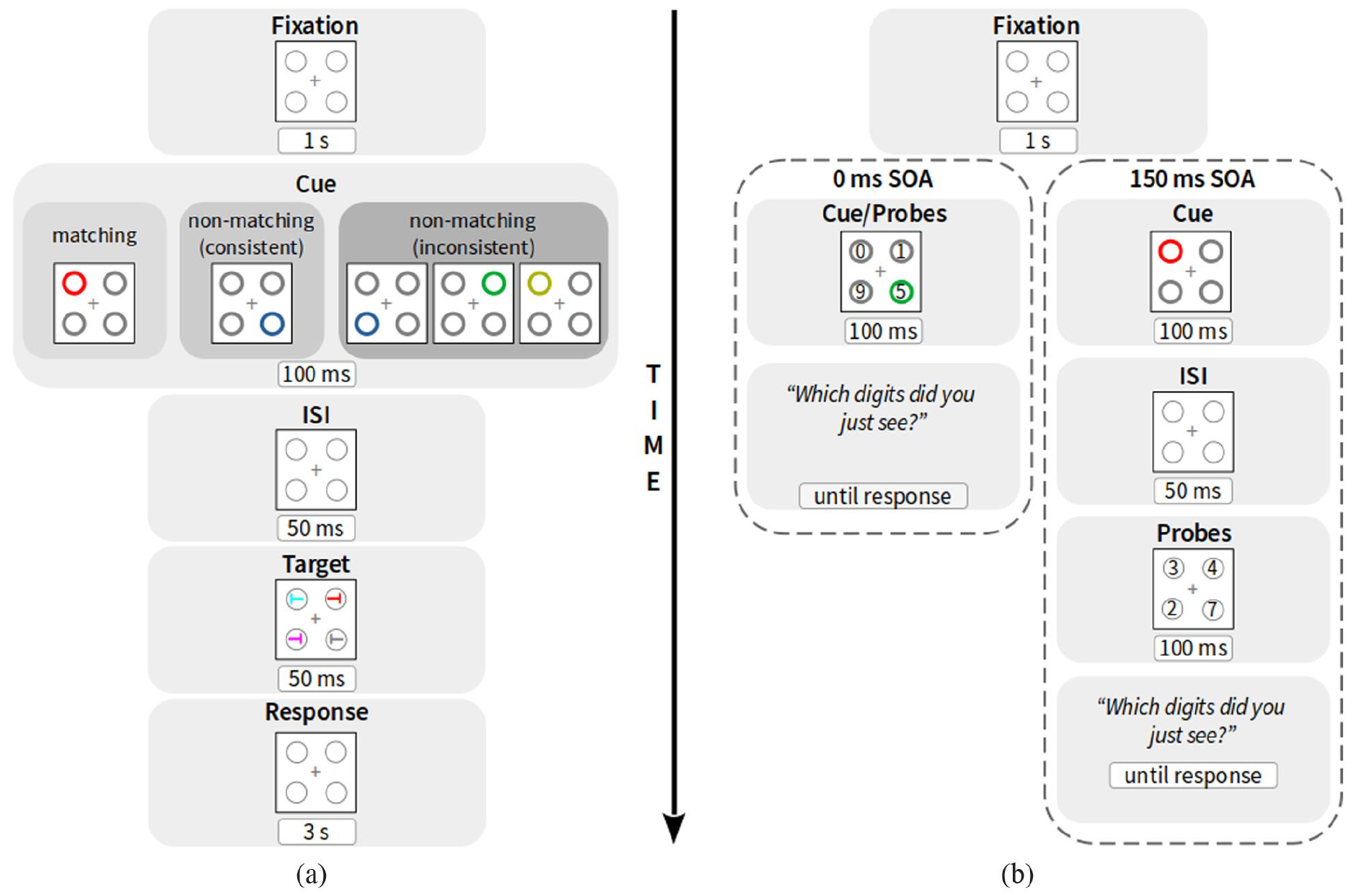
(a) Example of a search trial with matching, consistent non-matching, or inconsistent non-matching cue colours (from left to right). In the inconsistent non-matching condition, only one of the three possible cue displays was shown per trial. Note: In 0-ms cue-probe SOA blocks, digits were also present within the cues on each search trial (not depicted). (b) Examples of probe trials under 0 ms (left) and 150 ms (right) cue-probe SOA conditions. ISI: inter-stimulus interval.
In probe trials of 0-ms cue-probe SOA blocks (Figure 1b), probe digits were presented in all cueing displays, so as not to alert participants of an upcoming alternative probe task and, thereby, inadvertently influencing their strategy in processing the cueing display in search-versus-probe trials. In 150-ms cue-probe SOA blocks, probe digits appeared for 100 ms instead of the target display.
Before each block, participants completed 20 practice trials (not analysed); breaks were allowed every 50 trials.
Results
Search trials
We only analysed RTs from correct trials. Means and SDs of RTs were calculated separately for each subject and all possible factor combinations. Trials with RTs exceeding subjects’ condition means by 2.5 SDs were trimmed (2.6%), and overall, accuracy was 95%. We ran repeated-measurements analyses of variance (ANOVAs), with the factors validity (valid and invalid), cue-match (matching and non-matching), non-matching cue consistency block (consistent and inconsistent), and cue-probe SOA (0 ms and 150 ms), on both RTs and error rates (ERs). Note that the only difference between the search trials in 0-ms and 150-ms cue-probe SOA blocks was the presence of digits in the cueing display in the former and their absence in the latter. Although we did not have any particular predictions about the influence of digits in the cueing display on search performance, we wanted to control for any variance stemming from this manipulation.
Response times
Main effects were found for validity, F(1, 38) = 96.33, p < .001, ηp2 = .72, and SOA, F(1, 38) = 40.78, p < .001, ηp2 = .52. Furthermore, significant two-way interactions were found for validity and cue-match, F(1, 38) = 103.53, p < .001, ηp2 = .72, cue-match and cue-probe SOA, F(1, 38) = 5.56, p = .024, ηp2 = .13, and non-matching cue consistency block and SOA, F(1, 38) = 5.49, p = .024, ηp2 = .13. As all variables entered a four-way interaction, F(1, 38) = 4.33, p = .044, ηp2 = .10, we focus on this interaction between all variables.
First, looking at trials from 150-ms cue-probe SOA blocks, we found a conventional contingent-capture effect in blocks with consistently coloured non-matching cues: Matching cues led to a 46-ms validity effect (i.e., an advantage in valid compared with invalid conditions, calculated as invalid performance minus valid performance), t(38) = 10.78, p < .001, d = 0.62, whereas non-matching cues did not capture attention (p > .05). The same pattern was found for blocks with inconsistently coloured non-matching cues, with significant validity effects for matching cues, 51 ms, t(38) = 7.44, p < .001, d = 0.74, and none for non-matching cues (p > .27).
Next, focusing on cue-probe 0-ms SOA blocks, a similar pattern of results was found for blocks with consistent non-matching cues, with a significant validity effect for matching cues, 46 ms, t(38) = 4.45, p < .001, d = 0.47, and an absence of attention capture by non-matching cues (p > .81). However, in blocks with inconsistent non-matching cues, not only did matching cues capture attention, 44 ms, t(38) = 6.73, p < .001, d = 0.64, but also non-matching cues, 17 ms, t(38) = 2.98, p = .005, d = 0.25 (see Figure 2). A comparison of validity effects by non-matching cues in consistent and inconsistent non-matching cue conditions revealed that the validity effects of inconsistent non-matching cues reliably differ from the validity effects of consistently coloured non-matching cues, t(38) = 2.21, p = .03, d = 0.51.
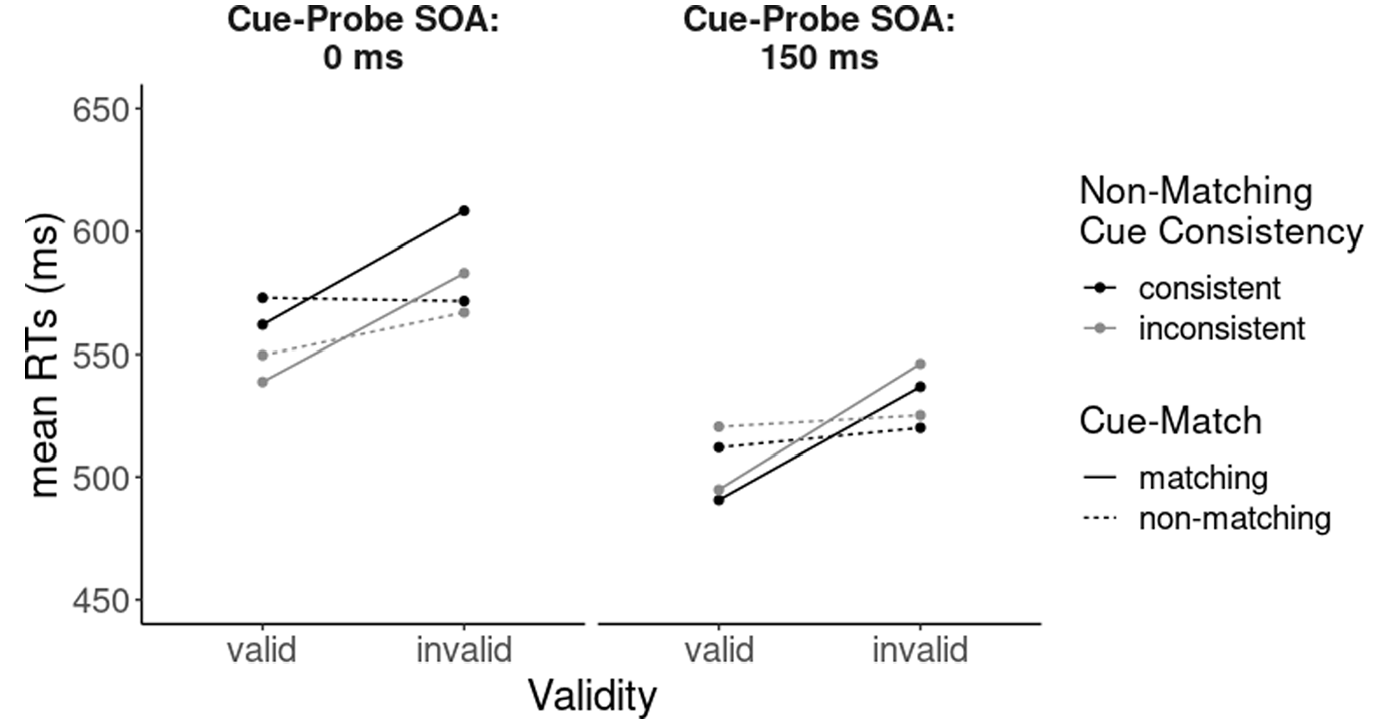
Three-way interaction between cue-match, non-matching cue consistency block, and cue-probe stimulus-onset asynchrony (SOA) in response times (RTs) of search trials.
Error rates
An identical ANOVA with arcsine-transformed ERs revealed main effects for validity, F(1, 38) = 137.28, p < .001, ηp2 = .78, cue-match, F(1, 38) = 7.51, p = .009, ηp2 = .17, and SOA, F(1, 38) = 8.02, p = .007, ηp2 = .17. Again, as validity and cue-match entered a two-way interaction, F(1, 38) = 7.76, p = .008, ηp2 = .17, and cue-match, non-matching cue consistency block, and cue-probe SOA entered a three-way interaction, F(1, 38) = 7.29, p = .01, ηp2 = .16, we focused on the higher-order interactions.
The interaction between cue-match and validity was due to different validity effects elicited by matching and non-matching cues: Matching cues led to a validity effect, 3.5% errors versus 6.6% errors, for valid and invalid cues, respectively, t(38) = 9.32, p < .001, d = 1.42. Non-matching cues led to a validity effect as well, although smaller, 3.7% versus 4.4%, for valid and invalid cues, respectively, t(38) = 4.92, p < .001, d = .78.
Concerning the three-way interaction between cue-match, non-matching cue consistency block, and SOA, we found that matching cues increased ERs compared with non-matching cues but only under 0-ms SOA conditions (matching cues: 5% vs. non-matching cues: 3%), t(38) = 3.48, p = .001, d = 0.62. Elsewise, ERs by matching and non-matching cues did not vary across SOA and non-matching cue consistency conditions (p > .07).
Capture-probe trials
Participants’ probe performance was calculated for digits presented at cued (i.e., singleton cue location) and un-cued locations (i.e., grey non-singleton locations). On each given trial, correctly reported probes were coded as 100% accuracy for singleton locations, whereas each correctly identified probe at non-singleton locations was counted as 33.3% accuracy. Averages were then calculated for each subject, probe location, and experimental block (i.e., consistently or inconsistently coloured non-matching cues).
Only one trial had to be excluded because more than four digits were reported. In the remaining capture-probe trials, participants reported 1.8 digits on average. The results of the probe-recall performance are shown in Figure 3.
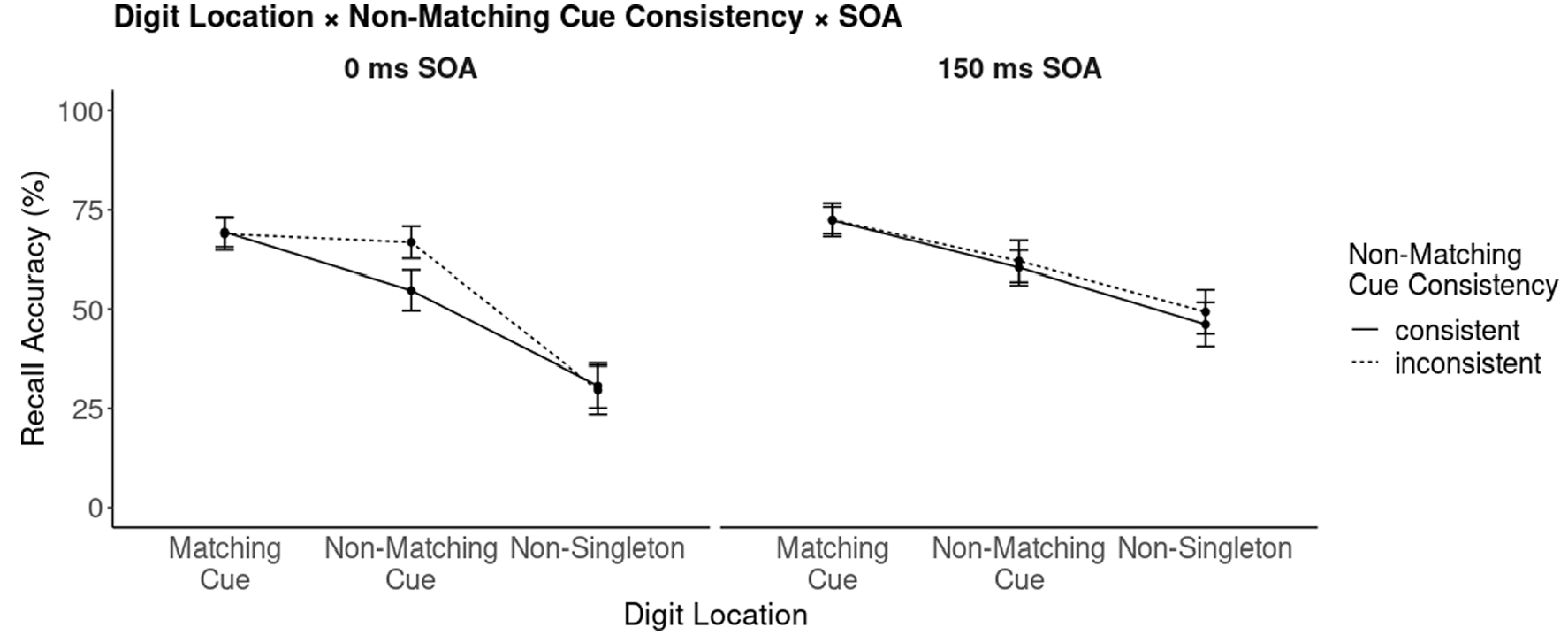
Three-way interaction between cue-probe stimulus-onset asynchrony (SOA), non-matching cue consistency blocks, and digit location in probe-recall. Error bars represent the 95% CI.
Participants’ recall accuracies were fed into a repeated-measurements ANOVA, with the factors digit location (matching singleton cue, non-matching singleton cue, non-singleton [in the cueing display]), non-matching cue consistency block (consistent, inconsistent), and cue-probe SOA (0 ms, 150 ms). Significant main-effects were found for all variables: digit location, F(2, 76) = 155.33, p < .001, ηp2 = .80, non-matching cue consistency block, F(1, 38) = 10.91, p = .002, ηp2 = .22, and cue-probe SOA, F(1, 38) = 26.12, p < .001, ηp2 = .41. In addition, the 2 two-way interactions between digit location and non-matching cue consistency block and between digit location and cue-probe SOA were significant, with F(2, 76) = 15.09, p < .001, ηp2 = .28, and F(2, 76) = 33.90, p < .001, ηp2 = .47, respectively. However, as all three variables entered a three-way interaction, with F(2, 76) = 11.58, p < .001, ηp2 = .23 (Figure 3), we examine this latter interaction more closely.
First, looking at recall accuracies under 0-ms cue-probe SOA conditions, we found a significant reduction of recall accuracy for digits presented at non-matching cue locations (55%) as compared with matching cue locations (69%) in consistently coloured non-matching cue blocks, t(38) = 8, p < .001, d = 1.04. In contrast, no significant reduction was found in blocks with inconsistently coloured non-matching cues, matching: 69% versus non-matching: 67% (p = .1). Digits at non-matching cue locations were recalled better than digits at non-singleton locations, both in consistent non-matching cue blocks (55% vs. 31%), t(38) = 7.66, p < .001, d = 1.4, and inconsistent non-matching cue blocks (67% vs. 30%), t(38) = 11.76, p < .001, d = 2.29.
One striking difference between the two cue-probe SOA conditions was the accuracy of probe-recall at non-singleton cue-display locations, where digits were recalled significantly better in 150-ms than in 0-ms cue-probe SOA conditions, both in consistent and inconsistent non-matching cue blocks. In blocks with consistent non-matching cues, recall accuracy for probes at non-singleton cue-display locations was 31% in the 0-ms and 46% in the 150-ms cue-probe SOA condition, t(38) = 7.90, p < .001, d = 0.86. The same, even larger, difference was found in blocks with inconsistent non-matching cues, with recall accuracies of 30% in 0 ms and 49% in 150 ms cue-probe SOA blocks, t(38) = 9.94, p < .001, d = 1.06. Similarly, SOA increased participants’ probe performance at locations of non-matching cues, but only in the consistent non-matching cue blocks, with 55% in the 0-ms and 60% in the 150-ms cue-probe SOA condition, t(38) = 2.29, p = .028, d = 0.38.
To explore the possibility that subjects learned to also attenuate inconsistently coloured non-matching singleton cues over the course of a block, we repeated the analyses with the added variable of block half (first half vs. second half), which indicated whether participants were in the first or second half of a given block. The reasoning behind this approach was that if participants learned to attenuate inconsistent non-matching cue features, performance for probes at inconsistent non-matching cue positions should approach performance for probes at consistent non-matching cues, at least in the second half of a block. However, block half entered an interaction neither with digit location and non-matching cue consistency block nor with digit location, non-matching cue consistency block, and cue-probe SOA (both p > .11), indicating that there was no meaningful learning effect regarding inconsistent non-matching singleton cue features across the course of a block, independent of SOA.
Discussion of Experiment 1
Experiment 1 investigated the temporal development of information accumulation and processing in the capture-probe protocol by varying the time between cue-display onset and the appearance of probe digits to be reported by the participants. We demonstrated that the time given to process irrelevant singleton-cue and non-singleton (colour) information in the cueing display selectively affected the accuracy of probe-recall, especially of probes at non-singleton cue-display locations and, most relevant for the present research, at the locations of consistently coloured non-matching cues relative to matching cues. For the latter, Experiment 1 demonstrates that, at early stages of information accumulation, information at locations carrying salient but task-irrelevant information (i.e., non-matching cues) can be selectively down-weighted when the irrelevant salient feature is consistent across trials. If the salient and task-irrelevant feature varies unpredictably from trial to trial, information at the cue’s location is processed to the same extent as information at locations carrying salient task-relevant singleton features. This finding is consistent with the findings of Gaspelin and Luck (2018a), that only specific features about which participants have first-order knowledge can be down-weighted (or their processing be attenuated, as we put it).
We further demonstrated that feature attenuation is a dynamic process which unfolds over time, as suggested by two findings. First, the aforementioned recall performance difference between probes presented at the position of consistently coloured salient non-matching cues (relative to probes at the position of a matching cue) in 0-ms cue-probe SOA conditions was missing under 150-ms cue-probe SOA conditions. Second, when given time (in our case, 150 ms), information to be accumulated at the non-singleton locations caught up with information accumulation from non-matching singleton-cue locations. The latter effect might be due to attenuation, too, as we will explain in the following section.
The observed drop in recall performance for probes at the position of a consistently coloured but not of an inconsistently coloured non-matching cue in 0-ms cue-probe SOA trials is similar to the finding of Kerzel and Barras (2016) that only inconsistently coloured non-matching cues prioritised processing of information at their location similar to matching cues. However, the fact that inconsistency did not significantly alter the attention capture influence of the irrelevant/non-matching cues under 150-ms cue-probe SOA conditions is, therefore, seemingly in contrast to that of Kerzel and Barras (2016).
One glaring difference between the experimental setup of Kerzel and Barras (2016) and ours is that our targets appeared well after the cues (i.e., a cue-probe SOA of 150 ms), while the task-irrelevant colour-singletons appeared simultaneously with the targets in the experiments of Kerzel and Barras. Information accumulation (e.g., of location and/or identity) may be more susceptible to irrelevant feature inconsistency at early processing stages than at later stages. In addition, the consistency of irrelevant colour singletons is also known to be less decisive for the degree of capture during colour search (e.g., Weichselbaum & Ansorge, 2018). This might be due to the fact that it is generally more difficult to suppress irrelevant but salient colours if targets are defined in the same feature dimension of colour (cf. Liesefeld et al., 2019; Liesefeld & Müller, 2019). To note, participants of Kerzel and Barras searched for targets defined in a different feature dimension compared to the singleton distractors, namely, shape targets, when their susceptibility to capture by irrelevant colour singletons was found to be stronger under inconsistent compared with consistent conditions.
Finally, we did not find a pattern of probe-recall accuracy that qualifies as suppression as defined by Gaspelin et al. (2015): worse probe-recall at suppressed irrelevant singleton locations than at non-singleton locations. Hence, it is more correct to refer to our result as “attenuation of information” rather than as “suppression.” However, research has shown that feature repetition enhances the suppression of salient-but-irrelevant stimuli (Experiment 1 of Pomper & Ansorge, 2021; Stilwell & Vecera, 2020). As the grey non-singleton colour was repeated most often, it is possible that participants learned to suppress grey non-singletons as well and, possibly, to a larger degree than the colour of the irrelevant singleton cues. In addition, rejection in groups was possible with non-singleton distractors but not with singleton cues (cf. Duncan & Humphreys, 1989). Hence, if the more-efficient suppression of the non-singleton colour than of the singleton colours caused this apparent absence of suppression, varying non-singleton colours randomly from trial to trial should make the suppression of salient-but-irrelevant singletons apparent in probe-recall performance. We tested this possibility in Experiment 2.
However, the differences between consistently and inconsistently coloured non-matching singletons on probe-recall were not accompanied by corresponding validity effect differences in search trials. It is true that, in Experiment 1, we also found a difference in the validity effects of consistently coloured versus inconsistently coloured non-matching cues under 0-ms conditions. This result is partly in line with performance in probe trials, where it also appeared as if inconsistently coloured non-matching cues captured attention more. However, this finding was not replicated in the second experiment. In addition, even in the inconsistent non-matching 0-ms SOA condition, validity effects in search trials were not as large as validity effects under matching conditions although performance in the probe trials would have suggested similar capture by both of these cues. This possibly points to a role of post-attentional influences of the inconsistently coloured non-matching cues on probe performance, for instance, a bias to start refreshing working memory representations of just seen and to-be-reported numbers at cued locations (cf. Kerzel & Renaud, 2023).
Experiment 2
In Experiment 1, we observed a significant variation of probe-recall accuracy at singleton cue locations between consistent non-matching and matching cue conditions (but not between inconsistent non-matching and matching cue conditions), at least at early stages of cue processing (i.e., with the 0-ms cue-probe SOA). However, while participants seemed to be able to attenuate processing of consistently coloured non-matching cues more effectively than inconsistently coloured non-matching cues, the criterion of suppression, according to Gaspelin et al. (2015)—that is, lower recall accuracies at suppressed (singleton) locations than at non-suppressed non-singleton locations—was not met.
Experiment 2 set out to investigate whether the feature consistency of non-singletons may have led participants to attenuate or suppress non-singleton locations as well or even more. Note that in the study by Gaspelin et al. (2015), non-singletons were consistently coloured as well. However, in their feature search, non-singletons still varied in the task-relevant shape dimension, which was not the case with our cue-display non-singletons.
Methods
Participants
Fifty-seven participants were tested in this experiment. Participants were, again, students at the University of Innsbruck, gave informed consent, and either received course credit for their participation or completed the experiment as partial course fulfilment.
Apparatus
As in the previous experiment, Experiment 2 was conducted online using the same software used in Experiment 1.
Stimuli
Experiment 2 was similar to the 0-ms cue-probe SOA blocks of Experiment 1. However, new non-singleton colours were introduced: yellowish (105, 105, 0), brown (74, 58, 22), and light blue (114, 159, 207). Colours were chosen to appear approximately equiluminant and sufficiently different from the target and singleton cue colours.
Design and procedure
Experiment 2 utilised a 2 × 2 × 2 × 2 factorial design consisting of four separate blocks. Besides validity (valid and invalid), cue (match and non-match), and non-matching cue consistency block (consistent and inconsistent), we introduced the factor cue displays’ non-singleton consistency (consistent and inconsistent). Although validity and cue-match varied within blocks, non-matching cue consistency and cue displays’ non-singleton consistency varied between blocks. Block order was balanced across participants. As in Experiment 1, per block, participants completed 70% of search trials and 30% of probe trials, randomly interspersed with search trials.
Results
As in Experiment 1, RTs deviating more than 2.5 SDs from each participant’s respective condition mean were removed (2.7%). Only correct trials were used for RT analyses (93.7%).
Search trials
Response times
A repeated-measurements ANOVA, with the factors validity (valid and invalid), cue-match (matching and non-matching), non-matching cue consistency block (consistent and inconsistent), and cue displays’ non-singleton consistency (consistent and inconsistent), was performed. Main effects were found for validity, F(1, 56) = 91.95, p < .001, ηp2 = .62, and non-matching cue consistency block, F(1, 56) = 4.32, p = .042, ηp2 = .07. Overall, we found a validity effect of 22 ms, and RTs in blocks with inconsistently coloured non-matching cues were 11 ms slower than those in blocks with consistently coloured non-matching cues.
The only significant interaction was found between validity and cue-match, F(1, 56) = 103.68, p < .001, ηp2 = .62, reflecting contingent capture: Matching cues produced a validity effect of 44 ms, t(56) = 12.78, p < .001, d = .59, whereas non-matching cues did not (p = .7).
Error rates
The identical analysis with arc-sine-transformed ERs as the dependent variable resulted in a significant main effect of validity, F(1, 56) = 79.19, p < .001, ηp2 = .59, and a significant interaction between validity and cue-match, F(1, 56) = 17.27, p < .001, ηp2 = .24. However, all four variables significantly interacted with each other, F(1, 56) = 5.25, p = .026, ηp2 = .09. Therefore, we focused on the higher-order interaction in our post hoc analyses.
First, we looked at blocks with consistently coloured non-singletons in the cue displays. When non-matching cues were consistently coloured, significant validity effects were found for matching cues (valid: 5% vs. invalid: 7.5%), t(56) = 5.17, p < .001, d = .68, but not for non-matching cues (p > .05). The same was found for blocks with inconsistently coloured non-matching cues, where only matching cues elicited a validity effect (valid: 4.8% vs. invalid: 7.1%), t(56) = 5.02, p < .001, d = .75, but not non-matching cues (p > .25). However, when looking at blocks with inconsistently coloured non-singletons in cue displays, this pattern was slightly more intricate. Again, starting with blocks with consistently coloured non-matching cues, significant validity effects were found for matching cues (valid: 4.9% vs. invalid: 8.5%), t(56) = 6.25, p < .001, d = 0.65, but also for non-matching cues, albeit being far less pronounced (valid: 7% vs. invalid: 7.1%), t(56) = 2.13, p = .037, d = 0.19. When non-matching cues were inconsistently coloured, validity effects were, again, only observed for matching cues (valid: 3.6% vs. invalid: 5.9%), t(56) = 5.39, p < .001, d = 0.79, but not for non-matching cues (p > .25).
Capture-probe trials
As in Experiment 1, participants’ probe performance was calculated for digits presented at matching and non-matching cue locations and at locations of non-singletons in the cue displays. As in Experiment 1, participants reported 1.8 digits on average. Participants’ mean probe-recall accuracies were fed into a three-factorial repeated-measurements ANOVA, with the factors digit location (matching cue location, non-matching cue location, non-singleton location in the cue display), non-matching cue consistency block (consistent, inconsistent), and cue displays’ non-singleton consistency (consistent, inconsistent).
We found a significant main effect for digit location, with F(2, 112) = 195.20, p < .001, ηp2 = .78. Recall accuracies, on average, were 66%, 55%, and 29% for digits at matching cue locations, non-matching cue locations, and non-singleton positions in the cue display, respectively (all differences p < .001). Digit location entered significant two-way interactions, one with non-matching cue consistency block, F(2, 112) = 4.21, p = .017, ηp2 = .07, and one with cue displays’ non-singleton consistency, F(2, 112) = 12.97, p < .001, ηp2 = .19.
First, examining the interaction between digit location and non-matching cue consistency block (Figure 4a), we found a similar pattern as in Experiment 1. In consistent non-matching cue blocks, the difference between recall accuracies at positions of matching (66%) and non-matching (54%) cues was significant, t(56) = 9.79, p < .001, d = 0.98. The same, yet less pronounced, was found in blocks with inconsistently coloured non-matching cues, where recall accuracy was better at positions of matching (65%) than at positions of non-matching (56%) cues, t(56) = 7.79, p < .001, d = 0.69. To check whether this difference was significant, we calculated participants’ individual differences in recall accuracy between matching and non-matching cue locations, separately for consistent and inconsistent non-matching cue blocks. A paired-sample t-test revealed a significant difference, t(56) = 2.76, p = .008, d = 0.41. In contrast, recall accuracies for digits presented at cue displays’ non-singleton locations did not differ between consistent and inconsistent non-matching cue blocks (p = .3).
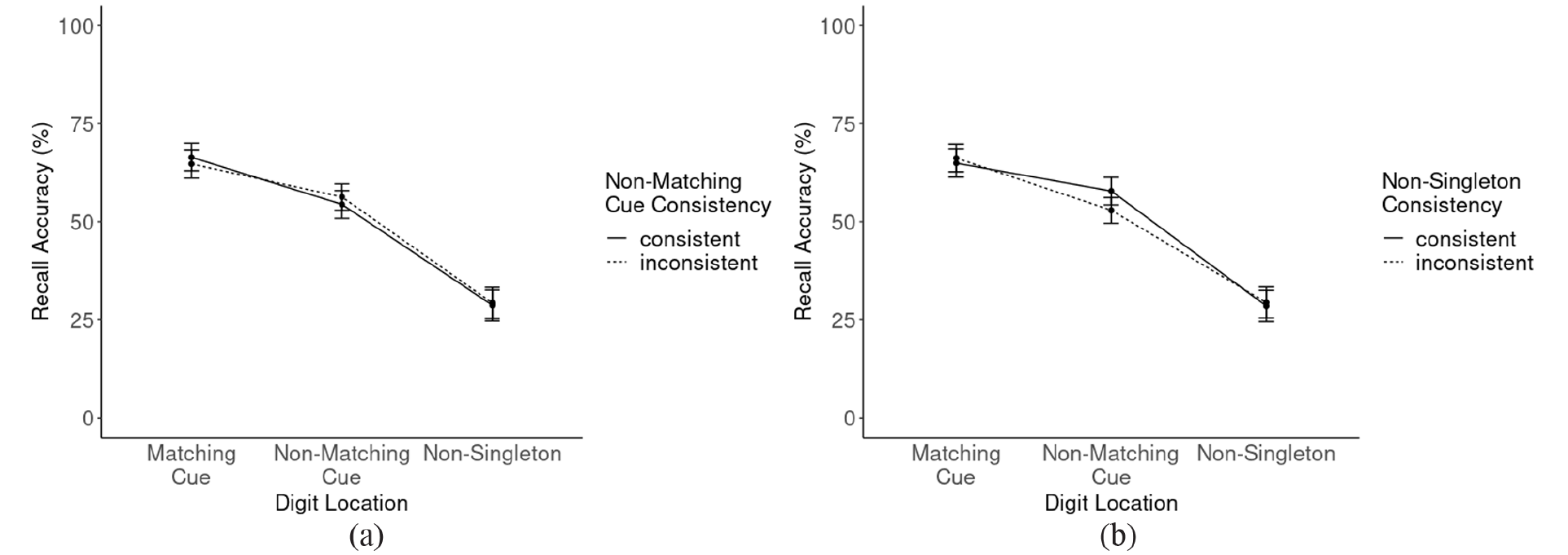
Probe-recall accuracies as a function of (a) digit location and non-matching cue consistency block and (b) digit location and cue displays’ non-singleton consistency. Error bars represent the 95% CI.
Next, turning to the interaction between digit location and cue displays’ non-singleton consistency (Figure 4b), we found no difference in recall accuracies at positions of matching cues between consistent and inconsistent cue displays’ non-singleton conditions (65% vs. 66%, respectively; p = .29) or at positions of cue displays non-singletons (29% vs. 29%, respectively; p = .28). However, at locations of non-matching cues, digits were recalled more precisely, when non-singleton cues were consistently coloured (58%) than when their colours varied unpredictably (53%), t(56) = 4.60, p < .001, d = 0.41. This finding may explain the difference between Experiment 1 and the present Experiment 2 concerning the recall accuracies at locations of matching and non-matching cues, as the introduced non-singleton colour variation may have made non-matching cues less salient. However, this is only speculative, as the three-way interaction between all variables was not significant (p = .42).
Again, no influence of learned inconsistent feature attenuation was found in a separate analysis with block half as an additional factor.
Discussion of Experiment 2
Experiment 2 examined whether cue displays’ non-singleton consistency in Experiment 1 may have reinforced participants’ suppression (or attenuation) of non-singleton features more effectively than that of even consistently coloured non-matching singleton cues. Here, we did not find evidence for this. However, it seems that cue displays’ non-singleton inconsistency did decrease the processing of non-matching singleton cue features. This might reflect some sort of dilution of attention across the non-singletons under inconsistent non-singleton conditions, such that fewer resources were available for the singleton cues—a factor that might have had a higher impact in case of the less-relevant non-matching cues than in case of the relevant matching cues.
Regarding the influence of non-matching singleton cue (in)consistency, we found, similar to Experiment 1, worse recall accuracies at non-matching singleton cue locations relative to matching singleton cue locations, more so under consistent than under inconsistent non-matching singleton cue conditions. However, in comparison to Experiment 1’s 0-ms SOA condition, this difference was weak.
Pooled analysis of Experiments 1 and 2
Similar to Experiment 1, results from Experiment 2 indicated an influence of non-matching singleton cue (in)consistency. However, while in Experiment 1, recall accuracy for inconsistently coloured non-matching cues was higher than that for consistently coloured non-matching cues and did not differ significantly from recall accuracy for probes at matching cue locations, this pattern was less pronounced in Experiment 2. Therefore, we ran a pooled analysis across Experiments 1 and 2. From Experiment 1, only 0-ms cue-probe SOA conditions were used. From Experiment 2, we only used data from consistent non-singleton cue blocks. Overall, data from 96 participants were included in this analysis. We ran a repeated-measurements ANOVA on recall accuracies with the factors digit location (matching cue location, non-matching cue location, and non-singleton location in the cue display) and non-matching cue consistency block (consistent and inconsistent).
We found main-effects for both digit location, F(2, 190) = 313.25, p < .001, ηp2 = .77, and non-matching cue consistency block, F(2, 190) = 7.56, p < .01, ηp2 = .07. However, as both factors entered a significant interaction, F(2, 190) = 16.60, p < .001, ηp2 = .1, we focused on the interaction. Recall accuracies did not differ between consistent and inconsistent non-matching cue blocks at matching cue locations (p = .19) and at non-singleton locations (p = .8). However, at non-matching cue locations, subjects recalled digits better in inconsistent blocks (62%) than in consistent blocks (56%), t(95) = 5.48, p < .001, d = 0.46. When comparing recall accuracies between matching and non-matching cue locations, there was a large difference in consistent blocks, matching: 67% versus non-matching: 56%, t(95) = 10.14, p < .001, d = 0.85. Although there was still a significant difference in inconsistent blocks, it was considerably less pronounced, matching: 66% versus non-matching: 62%, t(95) = 3.79, p < .001, d = 0.26.
Our pooled analysis shows that, even with a large sample, the consistency effect on probe-recall from non-matching cue locations from Experiment 1 is overall stable.
General discussion
The present experiments set out to investigate the attentional attenuation of irrelevant cues in the contingent-capture protocol in early and later stages of cue processing by way of a capture-probe protocol. Across experiments, we made use of feature consistency (vs. inconsistency) as a necessary or at least facilitating precondition of more (vs. less or no) attenuation: In one type of block, we used consistently coloured non-matching cues, and in another type of block, inconsistently coloured non-matching singleton cues. Furthermore, to tap into swift initial suppression and study its development across time, we used a cue-probe interval of 0 ms (in Experiments 1 and 2) and varied cue-probe SOA (in Experiment 1).
Search performance
We found clear-cut contingent-capture effects, with strong and significant validity effects under matching conditions and weaker to non-significant validity effects under non-matching conditions. This is in line with much prior research (e.g., Folk et al., 1992; for a review and meta-analysis, see Büsel et al., 2020). In comparison, manipulations of the cue-probe intervals or of the consistency of the non-matching cue colour on validity effects were very weak to non-existent. In our RTs, we did not find any influence of non-matching cue consistency on the contingent-capture effect using a conventional 150-ms cue-to-target SOA. In addition, a difference between validity effects in the 0-ms cue-probe SOA conditions between consistently versus inconsistently coloured non-matching cues were only found in Experiment 1 and could not be replicated in Experiment 2.
Probe-task performance
Our results regarding probe-task performance revealed more evidence of active attenuation of the processing of the consistently coloured non-matching cues compared with the matching cues than of the inconsistently coloured non-matching cues compared with the matching cues: When non-matching cues were consistently coloured, participants’ memory of probes (here: digits) at cued locations was significantly reduced compared with matching cues. With inconsistently coloured non-matching cues, the same performance difference was either not significant (Experiment 1) or significantly weaker (Experiment 2).
As with our RT findings in search-task trials, this is in line with a number of prior studies showing that repetition of irrelevant salient distractor colours supports their attenuation, if not even their suppression (Geyer et al., 2008; Kerzel & Barras, 2016; Pomper & Ansorge, 2021; Stilwell & Vecera, 2020; for suppression of frequently repeated irrelevant positions, see Heuer & Schubö, 2019; Wang & Theeuwes, 2018). Previous research has shown that experience with a specific distractor feature improves subjects’ attenuation of the processing of this feature (Vatterott & Vecera, 2012; but see Won et al., 2019). Hence, the participants’ higher expectancy of the non-matching singleton cue feature due to the increased presentation probability may have aided participants to successfully attenuate capture by the non-matching singleton cue feature in the consistent condition but less so in the inconsistent condition.
Note that other studies have demonstrated that also second-order suppression (i.e., the suppression of a feature dimension rather than a single known feature) is possible and is not necessarily less effective than first-order suppression (e.g., Ma & Abrams, 2023; Won et al., 2019). Importantly, however, these studies used shape targets and colour singletons as salient distractors. It is possible that the dimensional overlap in our study (i.e., colour targets and colour distractors/cues) made it difficult for participants to apply second-order suppression to a dimension that was also relevant to the task (e.g., Pomper et al., 2023; Weichselbaum & Ansorge, 2018; but see Kerzel & Huynh Cong, 2020). Maintaining search templates for features from different dimensions is known to weaken top-down guidance (Biderman et al., 2017). Hence, it might be possible that in other forms of top-down control (e.g., “negative templates” for suppression), the dimensional relationship between relevant and irrelevant stimuli does play an important role (Liesefeld et al., 2019; Liesefeld & Müller, 2019). In addition, even if second-order features were used for suppression in the current study, these features were shared across consistent and inconsistent feature conditions. Therefore, second-order feature suppression was not responsible for the differences between consistent and inconsistent conditions.
In the present experiments, even consistently coloured non-matching cues were probably not suppressed in all trials or even not in a majority of the trials. This was reflected by a better recall from locations of non-matching singleton cues than from non-singleton positions. Hence, significant reductions in recall accuracy at inconsistently coloured compared with consistently coloured non-matching cue locations indicate “more-or-less” rather than an “all-or-nothing” form of suppression or attenuation. In fact, such consistency-dependent attenuation might have played a role in the performance-dependent effects of colour consistency in the non-singleton conditions of Experiment 2, too. An attenuating influence of higher relative feature probabilities—here, of the non-matching cue colours relative to the non-singleton colour(s) (in the cueing display)—might have also been responsible for the unexpected observation of lower probe-recall from locations of non-matching cues where the cue displays’ non-singletons were inconsistently coloured (Experiment 2).
According to this argument, attenuation would have been more diluted across all four positions in cue displays (cf. Schoeberl et al., 2019) with consistently coloured non-singletons, as the latter would have been successfully attenuated to some extent, too, thereby weakening the potential performance difference to the non-matching cues. Still, we note that probe-recall from the locations of the non-singletons or from the location of the matching cues was not modulated by the consistency of non-singleton colours, indicating that this potential influence was too weak to overcome the singleton-specific effects and that it was “washed out” by the presence of three non-singletons and, thus, a levelled chance of the probe to be recalled from any of these three non-singletons.
Search and probe performance compared
Concerning the relation between validity effects in search-task trials and performance in the probe-task trials, we observed potentially interesting differences. First, cue validity effects of non-matching cues were the same at a more conventional 150-ms cue-target SOA, regardless of consistency. Second, however, performance in the probe-task trials was influenced by cue consistency. In fact, cue consistency influenced recall performance based on digit processing starting together with the cues. Assuming that proactive cue-elicited attenuation decreases across time since cue onset, digit processing at cue onset might have simply been more sensitive to consistency-dependent attenuation than the cue validity effect that is measured via responses to the trailing targets. Note that this is not the first experiment demonstrating that behavioural validity effects are not always sensitive enough to measure (differences in) attention capture. For example, Grubert and Eimer (2016) found no validity effects for (consistent) non-matching cues in RTs but did find a delayed, yet significant, N2pc towards non-matching cues.
This pattern of an early attenuation effect decreasing across time would be better in line with the idea of proactive attenuation (as, e.g., successful inhibition in at least a fraction of the trials) on the basis of templates for to-be-ignored features (cf. Arita et al., 2012; Forstinger et al., 2022; Moher et al., 2014; Woodman & Luck, 2007) than with the search-and-destroy hypothesis of Moher and Egeth (2012). According to the latter theory, irrelevant distractors would first capture attention and only then be actively attenuated or inhibited. Given that Moher and Egeth reported evidence for capture followed by active suppression of to-be-ignored distractors with distractor-probe intervals of 117 ms versus 167 ms, on the face of it, probe durations (of 100 ms) in the current study were too short to allow for initial capture before attenuation during probe-recall. In addition, as there was no evidence in the current study for more active attenuation of consistent than inconsistent non-matching cues in the cue validity effect when the cue-target SOA was 150 ms, our results would be more consistent with proactive attenuation and its subsequent decay across time.
Overall, our results are similar to those of Burnham (2020): Probes from top-down matching cue locations were recalled better than probes from locations of non-matching cues. However, the difference in recall accuracies between consistent and inconsistent non-matching cue conditions indicates a down-weighting, or attenuation of consistently coloured non-matching cues does indeed play a role in the contingent-capture protocol. Nonetheless, the present experiment does not call into question the role of top-down attentional guidance towards relevant features. We found contingent-capture effects both in search and capture-probe trials of the contingent-capture protocol: Top-down matching cues captured attention while top-down non-matching cues failed to do so. In addition, participants reported information from locations with matching cues with a higher precision than information from any other location in the cueing display in conventional 150-ms cue-probe SOA conditions. In essence, attenuation seems to play a role in the contingent-capture protocol but is usually not observed in RTs under the typically used search conditions, with a positive SOA between cues and targets (Burnham, 2020; see cueing effects in search trials of the present Experiment 1). Rather, the simultaneous presentation of task-irrelevant cues/distractors and targets or a continuous measure of attention versus suppression in the cue-target interval allows for suppression effects or attention capture to be observed (Arita et al., 2012; Gaspelin et al., 2015; Kerzel & Barras, 2016).
However, the selective modulation of probe-recall through consistency (vs. inconsistency) of non-matching cue colour(s) in the absence of a like difference in the validity effect might also be due to the fact that the two effects occur at different processing stages. For example, Kerzel and Renaud (2023) have argued for a post-attentional effect of cueing on probe-recall. Analogously, the present pattern of results might also reflect that cueing or at least cue consistency affected probe-recall via post-attentional processing stages, such as the order in which information in working memory is refreshed, giving digits at cued locations a higher chance to be successfully reported than digits at non-singleton locations, more so where attenuation of non-matching cue features is not at work (under inconsistent conditions).
Limitations
In our Experiment 1, the data suggest that presenting probes simultaneously with the cues might influence cue processing. When cues and probes were presented simultaneously, inconsistently coloured non-matching cues produced a validity effect. Although this validity effect was smaller than that of matching cues (17 ms vs. 44 ms, respectively), it was reliable, suggesting that increased perceptual load, increased cognitive load, or a mixture of both influenced singleton cue processing (Lavie et al., 2004). Nevertheless, the basic pattern of contingent capture (i.e., search goals modulating the extent to which distractors/cues capture attention) was observed. Generally, capture by irrelevant distractor features from the same feature dimension as is used to define the target—here, colour—is more persistent than capture by irrelevant features from a different feature dimension, meaning that distractor suppression and target search based on the same feature dimension could be particularly difficult (Liesefeld et al., 2019; Liesefeld & Müller, 2019; Pomper et al., 2023; Weichselbaum & Ansorge, 2018). In addition, as top-down control is generally subject to limited capacity or resources (cf. Huynh Cong & Kerzel, 2022), successful suppression of a salient colour distractor during the colour search could be particularly difficult if additional perceptual demands are imposed by the probe digits in the cueing display (cf. Lavie, 2005). However, Experiment 2 utilised a 0-ms cue-probe SOA as well and did not find any evidence for attention capture by non-matching cues by way of validity effects (1 ms; p > .7). Therefore, this influence of perceptual load on cue processing does not seem to be particularly reliable.
Furthermore, while conducting online experiments is beneficial for the acquisition of data from larger samples, this comes at the cost of lesser control. Although we instructed participants to maintain fixation at the central fixation point, we were not able to check if participants followed this instruction, and we cannot reconstruct the exact stimulus properties (e.g., size and luminance) seen by our participants. However, we were able to replicate the conventional contingent-capture effect, leading us to believe that our inclusion criteria for the data analyses were stringent enough to eliminate data from participants that were unable to perceive the stimuli properly. Furthermore, several studies have demonstrated that it is possible and feasible to measure visual attention effects outside of laboratory conditions (e.g., Experiment 2 in Büsel et al., 2021; Frătescu et al., 2019).
Besides online testing, one may be concerned about the typicality of our study design or, more specifically, the durations of the probe-/cue-displays and the overall cue-target SOA. Regarding the latter, looking at the 64 experiments included in the meta-analysis of Büsel et al. (2020), 33 experiments used a 150-ms SOA, following the original experimental setup of Folk et al. (1992). More critically, however, we presented our cues for 100 ms instead of the more conventional 50 ms. Specifically, 40 of 64 studies included in the meta-analysis by Büsel et al. (2020) presented the cues for 50 ms, while only eight studies presented cues for 100 ms. However, a meta-regression showed that the size of contingent-capture effects does not vary significantly with cue presentation times (but see Carmel & Lamy, 2015, for the influence of cue display durations on SLC).
Conclusion
The research presented in this article demonstrates several novel aspects concerning cue processing in the contingent-capture protocol. We demonstrate that at the early stages of cue processing, participants are capable of attenuating capture by consistently coloured non-matching cues more effectively than by inconsistently coloured non-matching cues, suggesting that templates used for distractor rejection might be limited to one feature at a time. Although our study was able to shed light on the early stages of cue processing using behavioural measures, future research should utilise electrophysiological methods to further our understanding of top-down guided search as well as suppression.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The present research was supported by the Tiroler Wissenschaftsförderung (TWF; 325957).